
Abstract
Dax1 (Nr0b1;
Introduction
Dax1
Recently, Dax1 has been identified as an important component of the pluripotency gene regulatory network in mouse embryonic stem cells (ESCs). The expression of Dax1 is affected by different stages of pluripotency [5,6]. Dax1 in conventional ESCs [cultured in leukemia inhibitory factor (LIF)/serum] was higher compared with ground-state ESCs (cultured in 2i/LIF), and it was almost undetectable in epiblast stem cells cultured in medium containing Activin-A and basic fibroblast growth factor (bFGF). Nanog expression is heterogeneous in conventional ESCs and reflects two different and interchangeable cell states, poised to self-renew (N+/Nanog-high subpopulation) or to differentiate (N–/Nanog-low subpopulation) [7,8]. Dax1 and Nanog act in parallel to stabilize mouse ESC pluripotency by directly inhibiting Gata6 transcription, and Dax1 is indispensable for self-renewal of Nanog-low ESCs [9].
The mouse Dax1 gene consists of two exons, that encode amino acids (aa) 1–391 and 392–472, respectively. Structurally, Dax1 contains a ligand-binding domain (aa 205–472) homologous to other nuclear receptors, including the PCFXXLP motif (aa 275–281) and the activation function (AF)-2 domain (aa 463–468). Its N terminus lacks the canonical zinc finger or any other known DNA binding motif, but contains an atypical DNA-binding domain with three repeats of LXXLL motifs instead. In mouse ESCs, Dax1 binds to LRH1 and Pou5f1 and inhibits their transcriptional activity [10,11], through the PCFXXLP motif and the aa 308–380 region, respectively. LXXLL motifs participate in protein–protein and protein–DNA/RNA interactions associated with different aspects of transcriptional regulation [12], but the role of this unique N-terminal extension of Dax1 in the maintenance of pluripotency of ESCs is unclear.
Alternative splicing is a ubiquitous regulatory mechanism of gene expression that allows generation of more than one unique mRNA species from the same precursor mRNA. These differences in coding sequence might affect their stability, localization, translation, or even function. Alternative splicing events can be categorized into at least six major types, including cassette exon skipping, mutually exclusive exons, alternative 5′ splice site selection, alternative 3′ splice site selection, alternative retained intron, and tandem cassette. There are more complex patterns, such as intraexonic alternative splicing [13,14], but far fewer than these main types have been discovered so far. Recently, extensive alternative splicing patterns were observed in ESCs and reinforced the connection between splicing and pluripotency maintenance/cell-fate commitment [15,16]. These findings demonstrate the key role of RNA splicing in regulating the pluripotency phenotype of ESCs. To the best of our knowledge, splice variants of Dax1 have not been identified and characterized in mouse ESCs.
In the present study, 18 novel alternative splice variants of Dax1 were identified in mouse ESCs. The putative proteins of all these variants lacked one to all three LXXLL motifs, and some are accompanied by the loss of the PCFXXLP motif. We characterized the expression pattern, subcellular distribution, and ability to maintain pluripotency of these Dax1 isoforms and highlighted the link between maintaining the pluripotency of Dax1 and its unique N-terminal structure.
Materials and Methods
Cell culture
Mouse ESC lines, including R1 (American Type Culture Collection), CCE (Stemcell Technologies), E14T (provided by Ian Chambers), Nanog:green fluorescent protein (GFP) (a gift from Shaorong Gao), and their derivatives, were cultured in LIF/serum and 2i/LIF as previously described [5]. P19, mouse embryonic fibroblasts (MEF), NIH3T3, and Bend3 cells were maintained in Dulbecco's minimum essential medium supplemented with 10% fetal bovine serum, 2 mM GlutaMAX, and 1% non-essential aa.
In vitro differentiation of ESCs
Embryoid body (EB) formation was performed in petri dishes with serum medium without LIF according to a previously described protocol [9]. Epiblast stem cells were derived from ESCs as reported [17] in N2B27-based medium containing 1% KSR, 10 ng/mL bFGF, and 20 ng/mL Activin A.
Flow cytometry
High and low GFP expressing Nanog:GFP cells (N+/N–) were subjected to sterile sorting (>99.5% purity, 5 × 105 cells) by flow cytometry [18] with four Laser BD FACSAria II Cell Sorter.
RNA isolation, reverse transcription, and polymerase chain reaction analysis
Total RNA was isolated with TRIzol (Invitrogen). cDNA synthesis was performed using the Transcriptor High Fidelity cDNA Synthesis Kit (Roche). 5′ and 3′- rapid amplification of cDNA ends (RACE) was performed with mixed RNA using a SMARTer RACE 5′/3′ Kit (Promega) according to the manufacturer's instructions. RACE polymerase chain reaction (PCR) products were electrophoresed on a 2% agarose gel, and different PCR products were gel-purified and cloned into the pGEM-T Easy vector (Promega) for sequence analysis. First round of nested PCR was performed using KOD FX (TOYOBO) according to the manufacturer's instructions. Second round of nested PCR and semiquantitative PCR were performed using GoTaq Hot Start Master Mixes (Promega). Single-cell PCR was performed according to previous reports [19]. Quantitative PCR (qPCR) was carried out using ExTaq SYBR Green Supermix (Bio-Rad). Primers used in this study are listed in Supplementary Tables S2 and S3.
Plasmid construction
The sequenced correct full-length open reading frame of Dax1-472/404/278/225/38 was subcloned from pGEM-T Easy into pPyCAG and pPyCAG-GFP. Dax1 promoter fragment (positions −500 to +23) was amplified by PCR from mouse genomic DNA and inserted into pGL3-basic vector (Promega). 3Flag-Dax1-472 expression vector and the reporter vector containing Gata6 promoter fragment (positions −1007 to +13) was previously constructed [9].
Plasmid transfection
E14T and NIH3T3 cells were transfected with plasmid DNA using Lipofectamine2000 (Invitrogen) according to the manufacturer's instructions. To obtain a cell line stably expressing the indicated protein, cells were subjected to puromycin (Invitrogen) selection (1.5 μg/mL) after transfection.
Protein extraction, Co-immunoprecipitation, and western blotting
Protein extraction, Co-immunoprecipitation (Co-IP), and western blotting were performed with anti-Dax1 (Ab85671; Abcam), GFP (AG279; Beyotime), FLAG (AF519; Beyotime), and β-Actin (A1978; Sigma), according to a previously described protocol [20].
Chromatin immunoprecipitation and luciferase assay
Chromatin immunoprecipitation (ChIP) and luciferase assay were performed using the ChIP Assay Kit (Upstate Biotechnology) and Dual-luciferase Reporter Assay System (Promega) according to the manufacturer's instructions, as previously described [9]. The primers for ChIP-qPCR were as follows: pGata6-promoter: Forward 5′-GTTACAGCGCTGGATGATTATG-3′, Reverse 5′-AGCTGCTAGCCCTGTCCTCAGA-3′. And pDax1-promoter: Forward 5′-AGAGGTCAGAGTCTAAGTTAATG-3′, Reverse 5′-TGTGTGGCCATGACCTCGAAAG-3′.
Immunofluorescence and images capture
Cells were fixed in 4% paraformaldehyde for 20 min at 4°C and permeabilized with 0.1% Triton X-100 for 15 min, followed by blocking with 1% bovine serum albumin/phosphate-buffered saline for 30 min. Cells were stained overnight with Gata6 antibody (AF1700; RD system, USA) at 4°C, followed by incubation with secondary antibodies for 1 h at room temperature. Cell nuclei were visualized by staining with 4′, 6-diamidino-2-phenylindole dihydrochloride. Images were captured with a confocal microscope (SP1, Leica Microsystems) or inverted biological microscope (Olympus, IX71).
Cell proliferation and colony formation assay
Cell proliferation and colony formation were assessed using a previously described protocol [9]. For proliferation assay, 1.5 × 104 cells were plated in gelatin-coated 12-well plates. After 5 days, viable cells were determined. For colony formation assay, cells were plated in gelatin-coated 24-well plates. After 6 days, alkaline phosphatase (AP) positive colonies were measured. Colonies were scored in three categories: undifferentiated, mixed (partially differentiated), and differentiated.
Apoptotic analysis
Cells were stained with Annexin V-APC (BD Biosciences) and propidium iodide for 10 min at 4°C and were analyzed with Novocyte (ACEA).
RNA-seq data analysis
Published transcriptome data were downloaded from the GEO DataSets database (
Statistical analysis
Data were presented as mean ± standard deviation of at least three independent experiments. The statistical significance of differences was estimated by one-way analysis of variance with appropriate post hoc tests. P < 0.05 was considered statistically significant.
Results
Identification of Dax1 splice variants by RACE
To obtain the full-length sequence of all possible transcripts of Dax1 in mouse ESCs, we performed 5′ and 3′ RACE PCR in a mixed sample of undifferentiated ESCs and ESCs during differentiation (Fig. 1A). Agarose gel electrophoresis showed that multiple bands were present in both RACE amplification products (Fig. 1B), indicating potential splicing events. The resulting RACE PCR products (100–2,000 bp) were subsequently cloned into vector and sequenced. In addition to the known transcripts Dax1-472, 18 novel transcripts of Dax1 were identified and named according to the number of aa in the putative proteins (Fig. 1C).
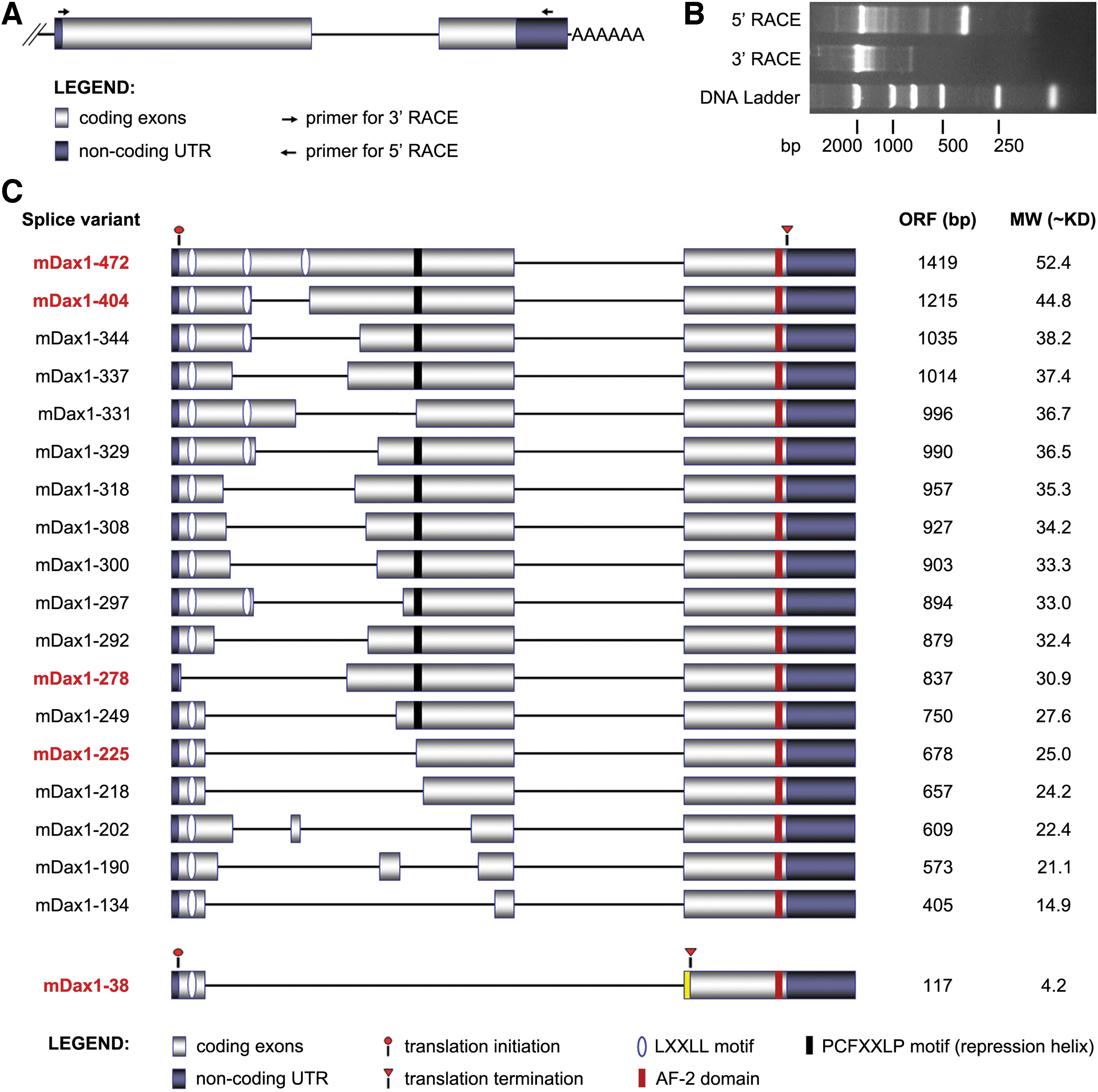
Identification of Dax1 splice isoforms by RACE-PCR.
All the novel transcripts contained full-length exon2 and partially deleted exon1. The putative proteins Dax1-404/344/331/329/297 lacked one LXXLL motif, Dax1-278 lacked all the LXXLL motifs, and other splice isoforms lacked two LXXLL motifs. Dax1-331/225/218/202/190/134 also lacked the PCFXXLP motif. The shortest isoform Dax1-38 not only lacked two LXXLL and PCFXXLP motifs but also had a novel truncated C terminus (TCLACSA) resulting from frame-shift mutations (Fig. 1C, Supplementary Table S1). No novel transcriptional start site (TSS) was found in our experimental system, and all the splice variants were presumed to start with the previously annotated TSS.
Transcription pattern of Dax1 splice variants
The tissue-specific transcription of Dax1 splice variants was detected by semiquantitative reverse transcription-polymerase chain reaction (RT-PCR) using primers within the conserved region. Consistent with previous reports, Dax1-472 was highly expressed in gonads, adrenal glands, and hypothalamus, and many other splice variants such as Dax1-278/225/38 were also found in these specimens. Only Dax1-472 was detected in the kidneys and spinal cord, and expression of Dax1-225 was detected mainly in the thymus (Fig. 2A). Dax1-472, Dax1-249, Dax1-225, and Dax1-38 transcripts were highly expressed in ESC lines such as R1, CCE, E14T, and teratocarcinoma cell line P19. None of the Dax1 transcripts was detected in MEF and somatic cell lines NIH3T3 and Bend3 (Fig. 2B).
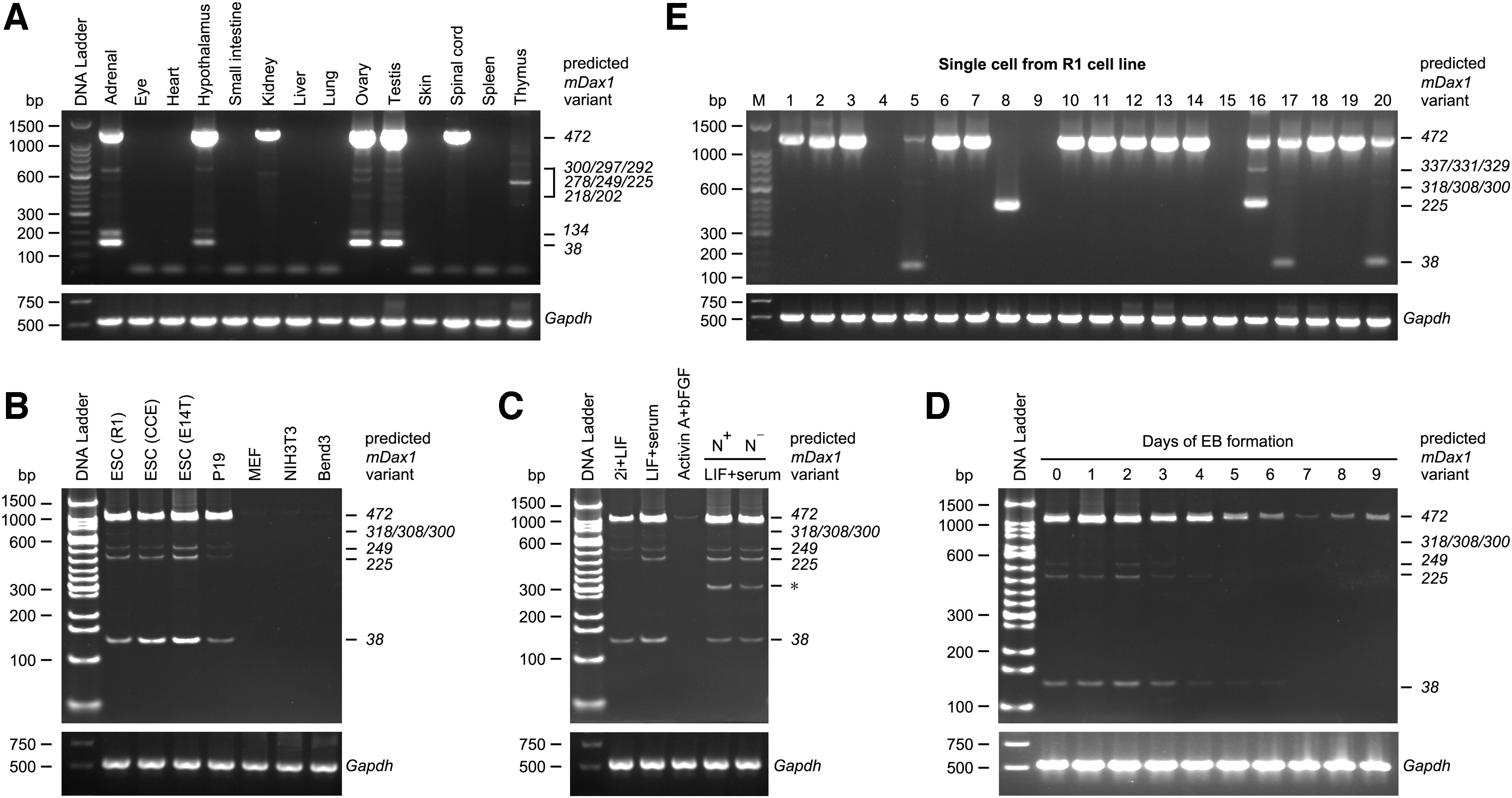
Transcription Pattern of Dax1 splice variants.
Then we detected the transcription of Dax1 splice variants in different pluripotency states. As expected, the transcription level of Dax1-472 in conventional ESCs was higher compared with ground-state ESCs, and it was almost undetectable in epiblast stem cells cultured in medium containing Activin-A and bFGF. Multiple splice variants such as Dax1-225 and Dax1-38 were also detected in ESCs. Nanog expression is heterogeneous in conventional ESCs, while the types of Dax1 splice variants and expression of individual splice variants were not significantly different between Nanog-high and Nanog-low cells (Fig. 2C). During EB differentiation, the transcription level of all Dax1 splice variants showed a gradual decrease (Fig. 2D).
Single-cell RT-PCR was used to detect whether the Dax1 splice variants were exclusive in single cells. Dax1 mRNA could be detected in the majority of cells (17/20). In some cells, only Dax1-472 transcript was detected (12/20), while in others, multiple of Dax1 splice variants coexisted with Dax1-472 (4/20). We also detected a cell with only Dax1-225 transcript (1/20) (Fig. 2E).
Recombinant expression of Dax1 splice variants
To obtain the functional information of Dax1 splice variants, five splice variants with higher expression levels and significant structural differences were selected for recombinant expression, including Dax1-472, Dax1-404 (lacking 1 LXXLL motif), Dax1-278 (lacking 3 LXXLL motifs), Dax1-225 (lacking 2 LXXLL motifs and PCFXXLP motif), and the shortest isoform Dax1-38. Stable Dax1 splice variants -GFP fusion protein overexpression (OE) cell lines were established individually in ESCs (E14T), Dax1 knockout (KO)-ESCs, and NIH3T3 cells. Semiquantitative RT-PCR was used to evaluate the transcription of Dax1 in E14T OE cell lines (Fig. 3A). The expression levels and molecular weight of the Dax1 splice variants were further verified by western blotting in NIH3T3 series (Fig. 3B) and in Dax1KO-ESC series (Supplementary Fig. S1A). Stable Dax1 splice variants OE (without fused GFP) cell lines were also established individually in E14T and Dax1KO-ESCs. This indicated that all these transcripts could be translated into proteins, which can be specifically recognized by Dax1 antibodies.
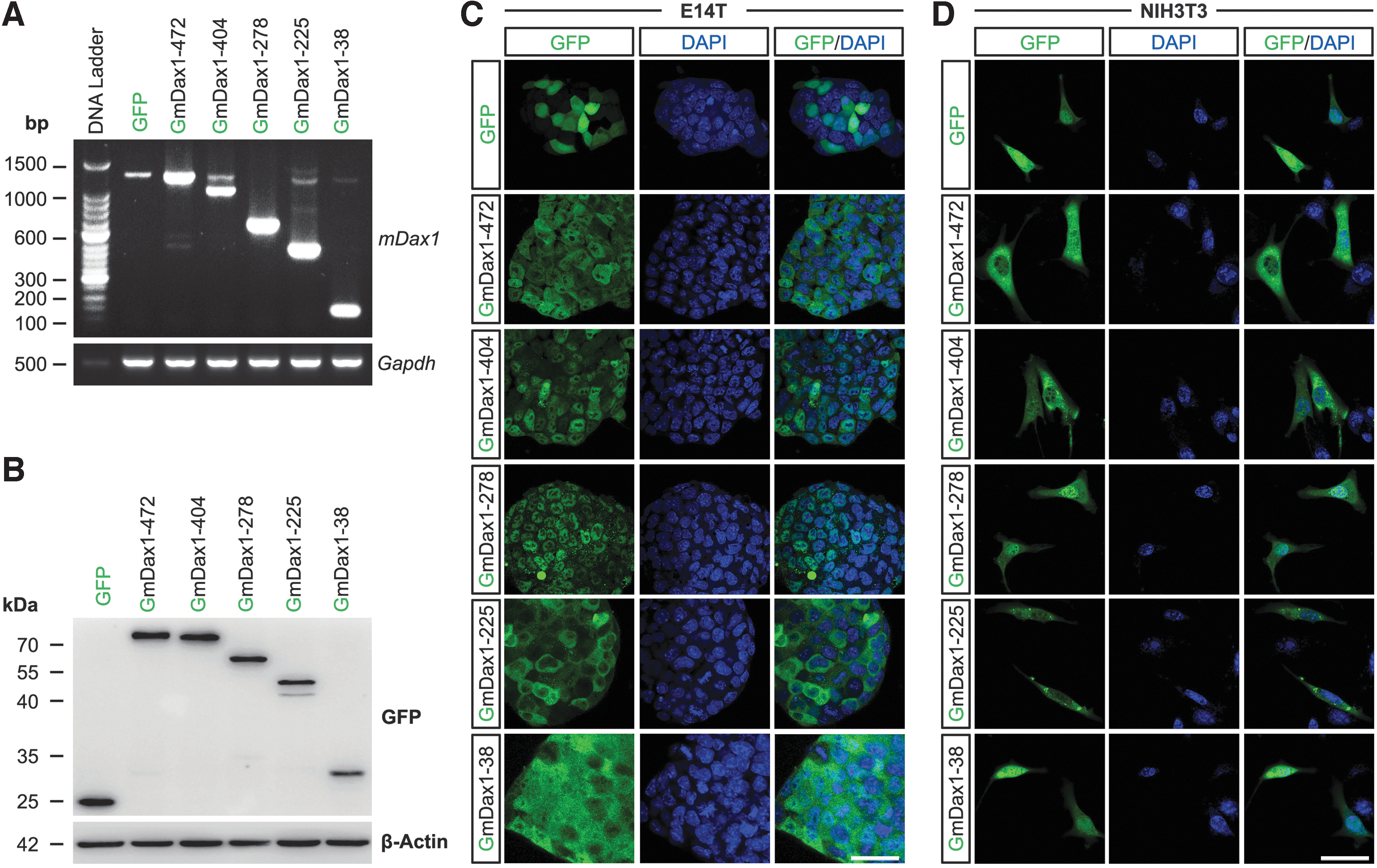
Expression and subcellular localization of Dax1 splice variants.
The endogenous expression of Dax1 splice variants was also detected by western blotting. We observed a ladder like distribution of proteins in ESCs but not in MEF and NIH3T3 (Supplementary Fig. S1B). This reflects the existence of Dax1 splice variants at the protein level.
Subcellular localization of Dax1 splice variants
Previous studies using truncated or mutated forms of Dax1 suggested that the LXXLL motif plays a role in nuclear localization of Dax1 [21,22]. It was interesting to observe the subcellular distribution of Dax1 splice variants, as they all lacked the LXXLL motifs (to varying degrees).
In Dax1 variants-GFP OE E14T cell lines, Dax1-472 was distributed in both the cytoplasm and nucleus, which was consistent with the previous reports. The distribution of Dax1-404 was the same as for Dax1-472. Dax1-225 was localized predominantly in the cytoplasm, and Dax1-38 was evenly distributed throughout the cell. Surprisingly, Dax1-278 was distributed only in the nucleus, although it lacked all the LXXLL motifs (Fig. 3C). The subcellular distribution of the five splice variants in NIH3T3 cells was similar to that in ESCs except for Dax1-278, which was distributed mainly in the nucleus, but could also be detected in the cytosol (Fig. 3D). This observation suggested that the LXXLL motif was not essential for the nuclear localization of Dax1.
The ability of Dax1 splice variants to maintain pluripotency of conventional ESCs
Consistent with previous reports [9], Dax1-472 OE cells exhibited normal ESC morphology with a slight reduction in the AP positive colony-forming and proliferation ability (Supplementary Fig. S2A–D). qPCR analysis showed that expression of ExEn lineage marker Gata6 was downregulated in Dax1-472 OE cells. The cellular and molecular properties of Dax1-404 OE cells were similar to Dax1-472 OE cells. Expression of Gata6 was downregulated in these two cell lines (Supplementary Fig. S2E). There was no significant difference between Dax1-278, Dax1-225, and Dax1-38 OE cells and control cells. Similar results were achieved in Dax1 variants-GFP OE E14T cell lines (data not shown).
To exclude the potential impact of endogenous Dax1, we evaluated the role of Dax1 splice variants in maintaining pluripotency in Dax1 KO ESCs [5]. When transferred from 2i/LIF to LIF/serum culture, Dax1KO ESCs showed a differentiated phenotype (Fig. 4A), The AP-positive colony size, number of colonies, and total cell number were significantly reduced (Fig. 4B–D), and the percentage of apoptotic cells was increased (Fig. 4E). Expression of differentiation markers was increased significantly in Dax1KO ESCs (Fig. 4F). As expected, Dax1-472 completely reverted the transcriptional and phenotypic effects of Dax1 KO. Dax1-404 was also able to rescue these phenotypes, while splice variants Dax1-278, Dax1-225, and Dax1-38 could not. These results suggested that Dax1-404 and Dax1-472 had redundant functions, and the third LXXLL motif of Dax1 was dispensable for its pluripotency-maintenance function.
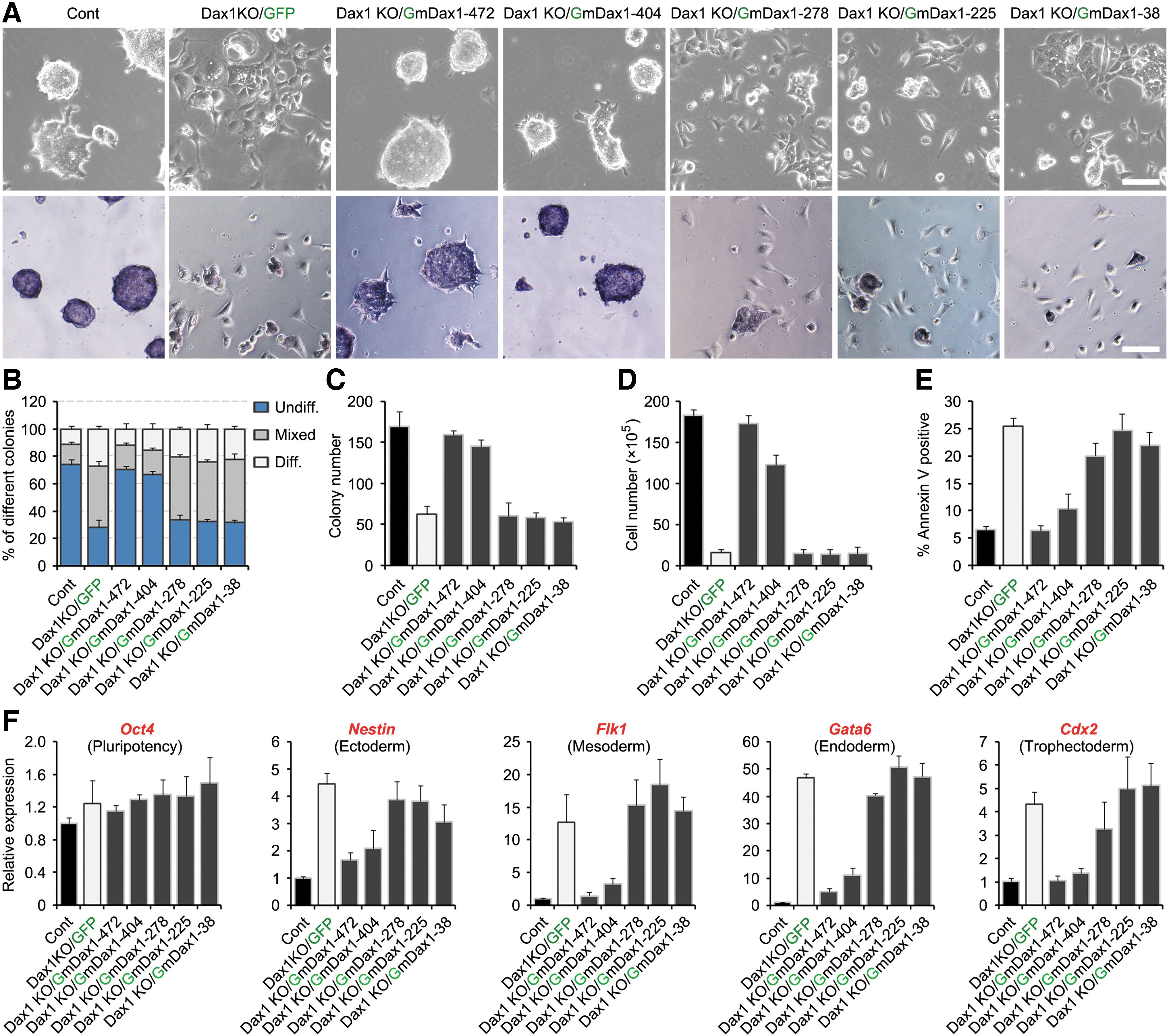
Phenotypic properties of the Dax1KO cell lines overexpressing the indicated Dax1 splice variant. Five splice variants fused with GFP were stable exogenous expression in the Dax1KO cell lines.
Effects of Dax1 splice variants on transcriptional regulation of Gata6
Dax1 is an important pluripotency transcription repressor, which prevents the commitment of ExEn by directly inhibiting Gata6 transcription [9]. It is not known whether these novel splice variants retain the biological function of Dax1. As expected, restoring expression of Dax1-472 completely reverted the excessive expression of Gata6 in Dax1KO ESCs (Figs. 4F and 5A). ChIP confirmed that Dax1-472 bound to the Gata6 promoter (Fig. 5B). Luciferase assay showed that Gata6 promoter activity was repressed by Dax1-472 (Fig. 5C). Dax1-404 had a similar transcriptional inhibitory effect as Dax1-472 on Gata6, while Dax1-278 and Dax1-225 OE had no effect. Dax1-38 bound to Gata6 promoter but failed to repress the transcriptional activity of Gata6 promoter (Fig. 5B, C). The above results indicated that the N-terminus of Dax1 was responsible for binding to the promoter of Gata6 and repressed its transcription, but the third LXXLL motif in Dax1 was dispensable for the function of Gata6 transcriptional repressor.
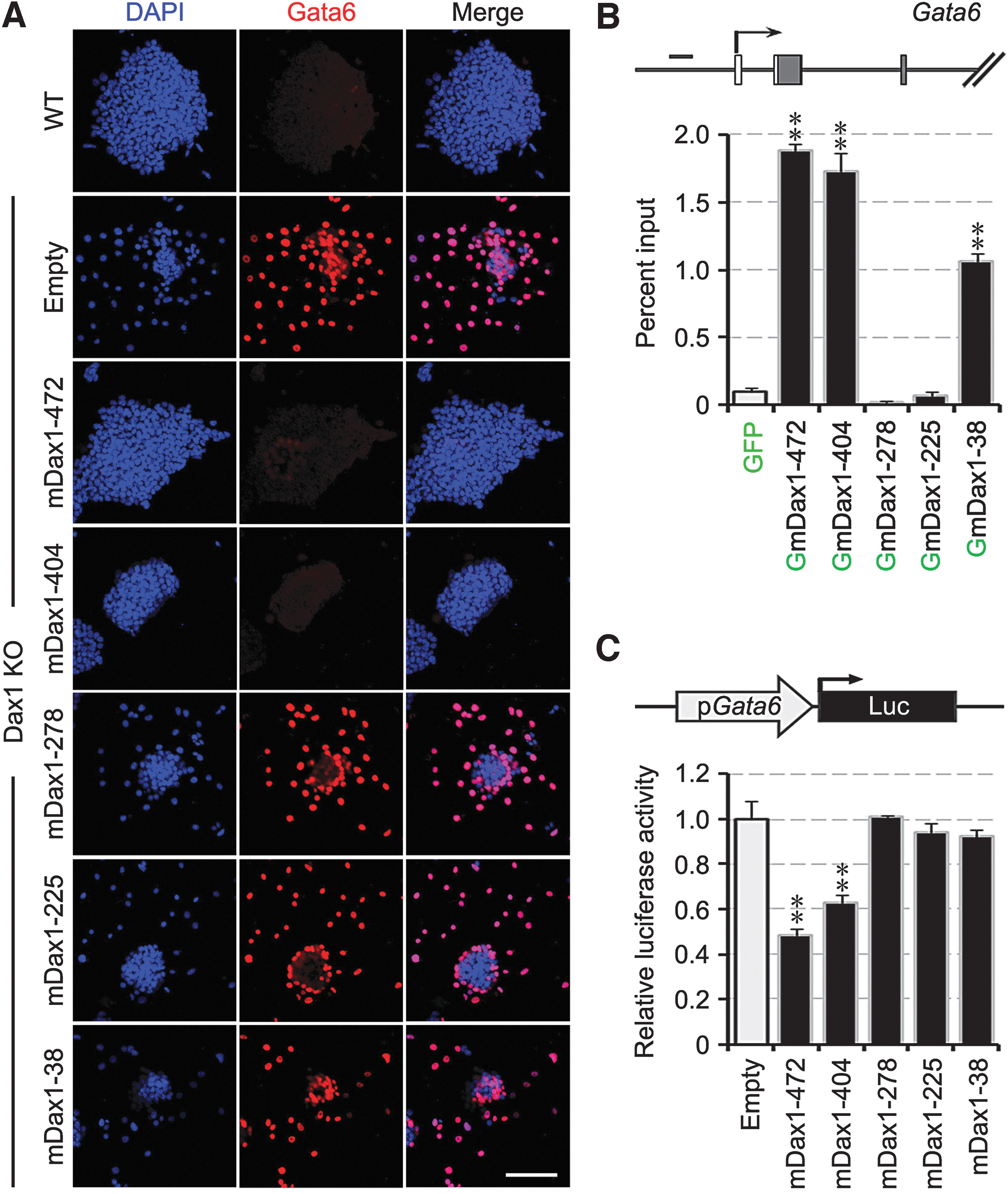
Effects of Dax1 splice variants on transcriptional regulation of Gata6.
The ability of Dax1 splice variants to repress Dax1 transcription and to form dimers with Dax1-472
Previous studies showed that Dax1 can repress the activity of its own promoter [23]. ChIP and luciferase assay were performed to detect the binding and inhibitory effects of these novel splice variants on Dax1 promoter. To exclude the potential interference of endogenous Dax1, these experiments were conducted in Dax1KO ESCs. As expected, Dax1-472 bound to and repressed the activity of Dax1 promoter. Dax1-404 had a similar transcriptional inhibitory effect as Dax1-472 on Dax1. The remaining three splice variants alone had no inhibitory effect on Dax1 transcription, although Dax1-38 bound to the Dax1 promoter (Supplementary Fig. S3).
It is reported that Dax1 forms antiparallel homodimers through the LXXLL motifs and AF-2 domain [24]. Considering that these novel splice variants coexpressed with Dax1-472, the interaction between these splice variants and Dax1-472 was analyzed. Co-IP demonstrated that all the five isoforms bound to Dax1-472, but the binding affinity between Dax1-38 and Dax1-472 was significantly weaker (Fig. 6A).
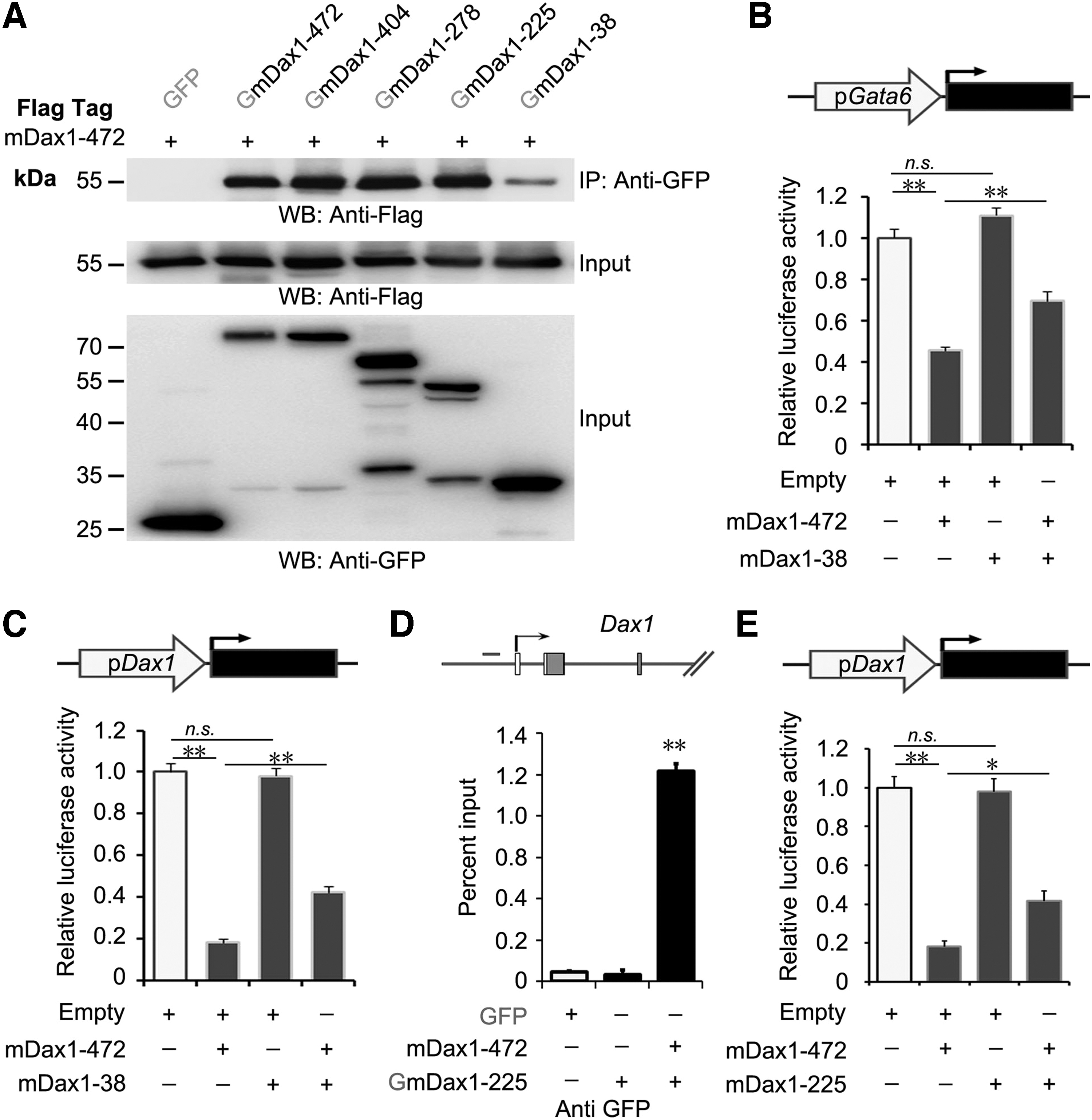
Interaction between Dax1 novel splice variants and Dax1-472.
The antagonistic effect of Dax1 splice variants on Dax1-472-mediated transcriptional repression
Dax1-38 bound to the promoter of Gata6 and upregulated its expression in the presence of Dax1 (mainly Dax1-472) by OE (Supplementary Fig. S2E). We hypothesize that Dax1-38 could compete with Dax1-472 on the binding site of the target promoter. As expected, the Gata6 and Dax1 promoter activity detected in the cells coexpressing Dax1-472 and Dax1-38 was significantly higher than in the cells expressing Dax1-472 alone (Fig. 6B, C).
Since these splice variants formed dimers with Dax1-472 (Fig. 6A), whether this protein–protein interactions affect the transcriptional repressive activity of Dax1-472 was detected. ChIP confirmed that Dax1-225 bound to the Dax1 promoter when coexpressed with Dax1-472 (Fig. 6D). The Dax1 promoter activity detected in the cells coexpressing Dax1-472 and Dax1-225 was significantly higher than in the cells expressing Dax1-472 alone (Fig. 6E). The above results suggested that the splice variants of Dax1 antagonized the function of Dax1-472 in different ways.
Discussion
Alternative splicing is prevalent in metazoan genomes, and the splicing pattern is dynamically regulated in different tissues and cell types, including ESCs. How much of this alternative splicing is functional at the protein level is an open question [25]. The alternative splice variants of some well-known pluripotent transcription factors (such as Oct4, Sox2, and Nanog) in pluripotent and nonpluripotent cells have been discovered and their functions evaluated [26]. An ESC-specific isoform of the FOXP1 gene can stimulate the expression of pluripotency genes and repress lineage-specific genes [27]. To our knowledge, this is the first report on the discovery and functional characterization of alternative splice variants of Dax1, which is an important guardian of pluripotency and self-renewal in mouse ESCs. The functional identification of these new splice variants provides more clues to elucidate the mechanism of pluripotency.
RACE-PCR, a traditional method for discovering splice variants [28,29], was used to identify the splice variants of Dax1 in this study. We also performed the expression pattern analysis of Dax1 variants using the existing RNA-seq data. From the analysis, we identified mDax1-249 and mDax1-225. The results showed that mouse Dax1-472 accounts for the majority, and Dax1-249 and Dax1-225 occupy 0.86% and 4% of the proportion in the NPCs and EB differentiation systems, respectively (Supplementary Fig. S4). Other variants of Dax1 were not detected in the RNA-seq data of these samples, possibly due to their low transcription level. It is suggested that RACE-PCR may be more sensitive to detecting splice variants with lower transcription levels.
Several studies, including ours, have shown that the intracellular localization of Dax1 protein is both nuclear and cytoplasmic in mouse ESCs [9,10,30] as in many other cells. In this study, Dax1-404, like Dax1-472, is a nucleo-cytoplasmic shuttling protein which is mainly in cytoplasm of NIH3T3 cells, and the nuclear localization was enhanced in ESCs. Interestingly, Dax1-278 was found only in the nucleus, and Dax1-225 was localized predominantly in cytoplasm in both cell lines. The molecular mechanism of the different subcellular positions of the two splice variants needs further study.
The transcriptional inhibition of Gata6 by Dax1 plays a key role in maintaining self-renewal [5,9]. In this study, Dax1-404, like Dax1-472, bound to the Gata6 promoter and inhibited its activity. Although studies have shown that Dax1 can bind to the hairpin loop structure in its own and SRA promoters [23], there is little evidence that Dax1 binds directly to DNA. We have not proven that Dax1 can directly bind to the promoter of Gata6 and Dax1. The promoter activity of Gata6 and Dax1 was inhibited by some splice variants of Dax1 (Dax1-472 and Dax1-404), which was only observed in ESC and not in NIH3T3 or HEK293 cells (data not shown). This indicates that unknown factors (protein and RNA) mediate the transcriptional inhibition of Dax1 on Gata6 in ESCs, and the LXXLL motifs and PCFXXLP motif might be involved in the binding of Dax1 to these unknown factors.
The exon part of the mouse Dax1 gene has 75% overall nucleotide sequence homology with its human homolog [1], but the introns are significantly different from each other. Previously, two alternatively spliced transcripts of human Dax1, Dax1A and Dax1α, have been identified in a variety of human tissues [31,32]. Dax1A and Dax1α have two mutually exclusive exon2 that are both located within the intronic region of the known transcript of Dax1. However, this type of isoforms is not found in mice. We recently discovered a similar type of splice variants (intraexonic deletion of exon1) of human Dax1 (unpublished data). This indicates that the alternative splicing pattern of Dax1 found in this study is evolutionarily conservative.
In summary, 18 new transcripts of Dax1 have been identified in mouse ESCs, and their encoded proteins are different from Dax1-472 in subcellular location and function.
Footnotes
Acknowledgments
The authors thank Dr. Ya-zhou Wu for his guidance and suggestions in statistical analysis.
Data Availability
The GenBank Accession No. for 18 novel alternative splice variants of Dax1 cDNA sequence is SUB13069566.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the National Natural Science Foundation of China (Grant No. 82273118) and Natural Science Foundation of Chongqing (Grant No. 20QNPY016).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3