
Abstract
The health of hair is directly related to people's health and appearance. Hair has key physiological functions, including skin protection and temperature regulation. Hair follicle (HF) is a vital mini-organ that directly impacts hair growth. Besides, various signaling pathways and molecules regulate the growth cycle transition of HFs. Hair and its regeneration studies have attracted much interest in recent years with the increasing rate of alopecia. Mesenchymal stem cells (MSCs), as pluripotent stem cells, can differentiate into fat, bone, and cartilage and stimulate regeneration and immunological regulation. MSCs have been widely employed to treat various clinical diseases, such as bone and cartilage injury, nerve injury, and lung injury. Besides, MSCs can be used for treatment of hair diseases due to their regenerative and immunomodulatory abilities. This review aimed to assess MSCs' treatment for alopecia, pertinent signaling pathways, and new material for hair regeneration in the last 5 years.
Introduction
The hair structure is made up of the hair trunk and root, and the hair root is surrounded by hair follicle (HF) structure, which is composed of epithelium and connective tissue. HF is one of the skin's appendages that develops from the ectoderm, and is involved in many functions, such as organism protection, thermal insulation, sensory perception, and social interaction. Mesenchymal and epithelial cells cooperate in HF to regulate hair growth and development [1]. The interaction of signaling pathways, such as Wingless and integrated (Wnt)/β-catenin signaling pathway, transforming growth factor-β/bone morphogenetic protein (TGF-β/BMP) signaling pathway, Notch signaling pathway, and Hedgehog signaling pathway establishes complex regulatory networks during the growth cycle of HF. Furthermore, the vascular and neural networks in HF are related to the skin microenvironment. Hair-related disorders may appear and evolve due to network dysfunction or aberrant signal activation, which may significantly harm human psychological health and social functions [2].
Although alopecia is common in modern people, there are limited effective treatments for these disorders now and cannot satisfy the needs and expectations of patients. The research progress of regenerative medicine has been increasing in recent years, where dermatologists are attempting to treat alopecia using regenerative medicine. Cell-based therapy is crucial for regenerative medicine, which includes mesenchymal stem cells (MSCs), tissue-induced pluripotent stem cells, fibroblasts, and other derivatives, such as platelet-rich plasma, and exosomes (Exos) [3]. These cells and blood products have gradually become the mainstays of regenerative medicine because of their potent capacity for tissue repair and regeneration.
MSCs, as pluripotent stem cells, are found in various mesenchymal tissues, including bone marrow, fat, umbilical cord, skin, tooth root, and others [4]. Therefore, MSCs are crucial for regenerative medicine since they have multiple sources. According to certain research findings, MSCs promote cell proliferation, block cell apoptosis, lower pain, increase blood supply to hasten wound healing, and also restore skin suppleness and barrier integrity [5,6]. Furthermore, MSCs can eliminate tissue inflammation and secrete cytokines and growth factors by inducing immune cells, thus promoting tissue repair [7]. Therefore, MSCs have excellent regenerative and outstanding performance in paracrine and immune control. However, the application of MSCs for hair regeneration is still in its cradle stages and warrants more research.
Morphogenesis of HF and Hair Cycle Transition
HF morphogenesis is crucial for studying hair diseases. First, HF epithelial and dermal cells aggregate to form the HF basal plate, generating new HF. The HF forms hair buds when it enters the dermis. Hair buds gradually thicken and become a columnar structure. Meanwhile, dermal papilla (DP) is formed beneath the basal plate. The structures, such as hair stem and inner root sheath, gradually appear after the formation of hair bulges, forming a complete HF structure [8]. The interaction between epithelium and mesenchyma (epithelial–mesenchymal interaction, EMI) around hair is directly related to HF morphogenesis. HF growth can be promoted by improving EMI through multiple pathways [9].
The hair cycle is a sign of HF activity, consisting of three phases: anagen, catagen, and telogen (Fig. 1). HF significantly grows during the anagen period and enters the subcutaneous tissue, while DP proliferates and hair stem and inner root sheath appear. During the catagen period, HF growth stops, cell apoptosis occurs, and DP significantly decreases. Hair falls off during the telogen period, but some of the regulatory factors begin to prepare for the next anagen period [10]. The variables, such as species, environment, age, and hormones, can affect the maintenance time and characteristics at each period [8,11]. Hair follicle stem cells (HF-SCs), located in the hair bulge of the outer root sheath (ORS), are crucial in the hair cycle [12]. HF-SCs briefly begin to proliferate and migrate downward to the bottom of HF at the early period of anagen [13].
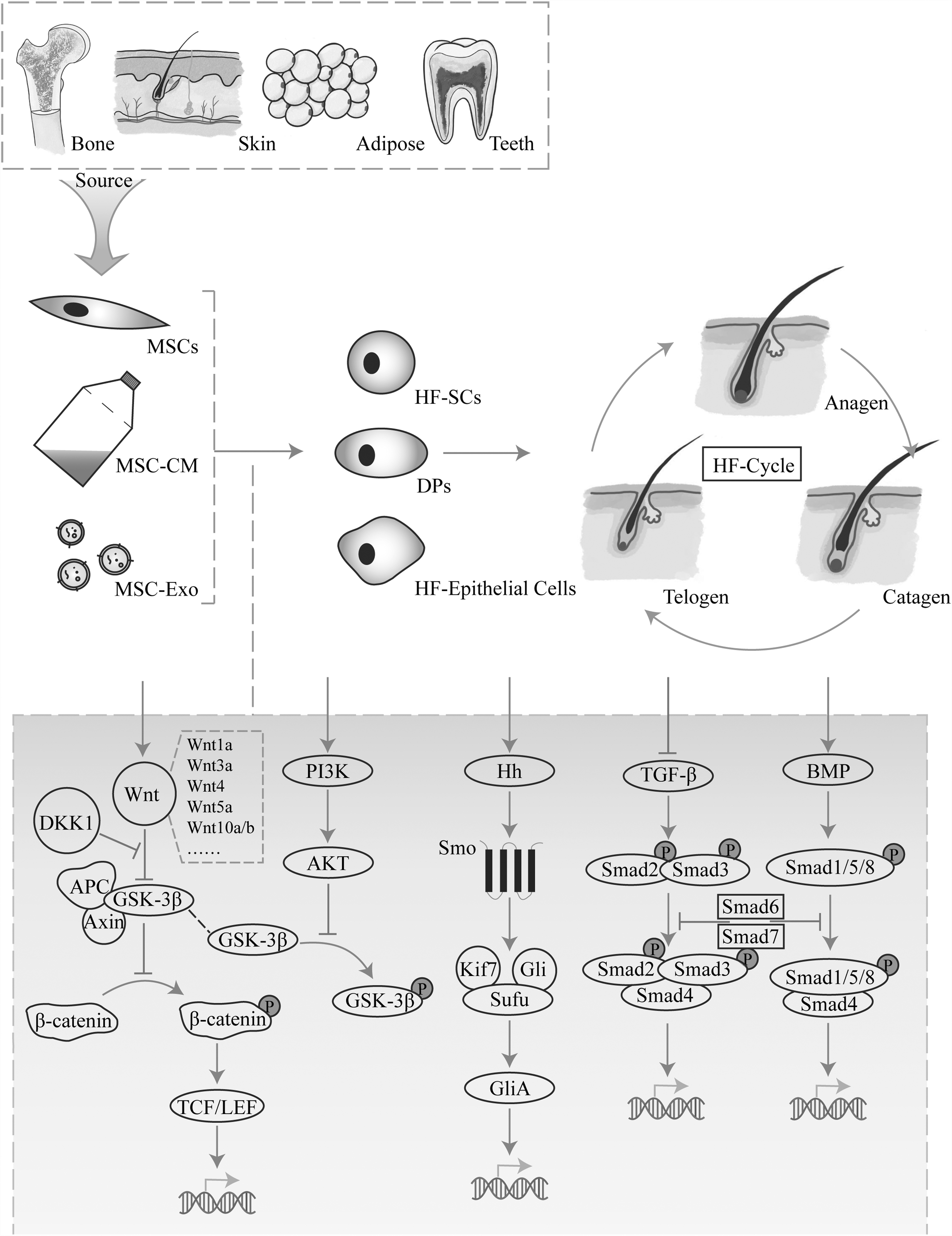
The role and signaling pathways affected by MSCs in HF. MSCs act on HF stem cells, hair papilla cells, and HF epithelial cells through cells, conditioned medium, and substances such as exosomes. MSCs activate Wnt/β-Catenin, AKT/GSK-3β, Hedgehog, BMP signaling pathway, and inhibiting TGF-β signaling pathway to achieve the effect of regulating hair cycle. BMP, bone morphogenetic protein; HF, hair follicle; MSC, mesenchymal stem cell.
Katou-Ichikawa et al. [14] demonstrated that monoclonal antibody A3, an antigen produced by rat malignant fibrous histiocytoma cells, can recognize N-glycan and peptide. Monoclonal antibody A3 can be used to identify stem cells with differentiation potential in hair cycle. The A3 antigen marks both epithelial and mesenchymal cells during the initial stages of anagen, and the signal intensity increases as HF grows. In contrast, only a small number of cells in the permanent area, lower isthmus, and infundibulum are labeled A3 during telogen. In addition, Sox10 can participate in early HF development and modifications to hair cycle [15]. Sox10 can be detected in various regions at different periods. For example, Sox10 appears in the ORS, regressed epithelial strand, and hair bulge/germ cell cap during anagen, catagen, and telogen, respectively.
The discovery of these marker antibodies describes the role of HF-SCs and other components in HF, and can also help in precise monitoring of the alterations and migration of hair cycle. It is advantageous that HF-SCs can be accurately labeled at various phases for studying and examining hair cycle. However, no complete stem cell biomarkers have been identified, making it very difficult to accurately locate HF-SCs in human skin.
The Effect of Various MSCs on Hair Regeneration
The MSCs that influence hair regeneration are divided into two categories: HF mesenchymal stem cells (HF-MSCs) and epithelial stem cells, and MSCs from other sources.
HF-MSCs and epithelial stem cells
HF-MSCs include dermal papilla cells (DPCs) and dermal sheath cells (DSCs), with DPCs located at the hair bulbs and DSCs in connective tissue sheath. Two types of cells connect at the bottom of HF, and these cells continuously renew themselves, differentiate, control hair development, and maintain skin's homeostasis, which can also directly stimulate HF regeneration and hair growth [16].
HF-MSCs can be used to treat alopecia areata (AA)-like in vitro models that are induced by interferon-γ (IFN-γ). Notably, this treatment can decrease the inflammation inside or around the HFs, CD8+T cells, and the expression of genes related to cell apoptosis and immune privilege damage, including caspase1, major histocompatibility complex class I (MHC I), tumor necrosis factor α (TNF-α), and Interleukin (IL)-6. Therefore, the alopecia in AA mice is also alleviated [17]. A study showed that intracutaneous injection of HF-MSCs is effective in androgenetic alopecia (AGA) patients' scalps. The average density of hair was significantly higher in the area where HF-MSCs were injected (23.3 roots/cm2), compared with the control area (0.7 roots/cm2), with no significant side effects [18]. However, HF-MSCs exhibit progressive dysfunction and become shrunken with age, losing their ability to maintain a normal hair growth and cycle, thus resulting in hair loss [19].
DPCs, as key targets in HF and multiple paracrine signal centers, are often used as indicators for observing and evaluating hair regeneration. Avigad Laron et al. [20] demonstrated that DPCs can limit the expression of Sonic hedgehog (Shh) signals to only act on cells near the DPC site. DPCs cause transient activation of resting progenitor cells by regulating Shh signals, thereby improving the function of hair regeneration. Moreover, a single Hoxc gene can awaken dormant DPC niches and promote local HF regeneration through canonical Wnt signaling. Yu et al., [21] showed that this activation does not apply to Hoxc gene activation in all cells, which is a particular way that Hoxc is expressed in DPCs. Histone deacetylase (HDAC) 1 and HDAC2 activation in DPCs can inhibit the expression of cell cycle genes and keep DP quiescent. HDAC1 and HDAC2 affect the expression of Wnt signaling pathways around DPCs during the anagen phase and participate in the hair cycle [22].
C-X-C motif chemokine receptor 4 (CXCR4), as a receptor for C-X-C motif chemokine ligand 12 (CXCL12), is expressed in DPCs and ORS. CXCR4 is activated when CXCL12 is highly expressed in dermal fibroblasts, thus increasing phosphorylation of signal transducers and activators of transcription (STAT) 3 and STAT5 in DPCs and ORS, delaying the transition from resting to growing phase and shortening hair length [23].
However, it is difficult to culture human-derived DPCs now. Therefore, ORS may become an alternative. The loss of immune privilege may lead to DP being attacked by macrophages, leading to the loss of key HF components. ORS cells can downregulate the expression of MHC I in DPCs, restore the immune privilege of HF, and promote EMI [24]. Li et al. [25] successfully extracted and amplified MSCs from ORS through noninvasive hair removal methods and gas–liquid culture, and showed that these cells can express MSC marker antibodies (CD44, CD73, CD90, and CD105).
Moreover, epithelial stem cells may be another alternative to HF-MSCs according to the principle of EMI. Interfollicular epithelial stem-like cells exhibit phenotypic characteristics similar to follicular keratinocytes. Furthermore, these cells can increase the proliferation of DPCs and alkaline phosphatase active cells, affecting the release of EMI mediators, such as platelet-derived growth factor-A, vascular endothelial growth factor, and BMP 2, resulting in HF and sebaceous glandular-like structures [26]. The CD49fhigh epithelial stem cells can induce cocultured HF epithelial cells to form HF-like buds' structures and control the expression of important genes [27]. Gentile P [28] grafted human intra- and extra-dermal adipose tissue-derived HF stem cells into the scalps of AGA patients, and showed that these stem cells, which contain HF-MSCs and epithelial stem cells, can increase hair density.
A recent study showed that epithelial stem cells from various tissues (except for skin), such as the cornea, esophagus, vagina, bladder, and prostate, can respond to skin morphogenetic signals and promote the growth of new HFs, sebaceous glands, and epidermis [29]. These cells also express the transcription factor Tp63, a master gene for the HF lineage. However, the lineage constraint is not unchangeable, it may manifest in different ways depending on where it is present.
The mixture of scalp-derived dermal progenitor cells and foreskin-derived epithelial stem cells forms pear-shaped structures (type I aggregates) [30]. This structure displays early HF markers, and initiates connections between two progenitor cells leading to high expression of the Wnt signaling pathway.
Although there are many recent publications on proliferation, aging, and activity of HF-MSCs, only a few studies or clinical trials have used HF-MSCs to treat alopecia. Therefore, further study is needed to understand the mechanism and relevant pathway regulation because there are few sources of HF-MSCs, which involves complex acquisition methods.
Other sources of MSCs
Other sources of MSCs include adipose MSCs (AD-MSCs), umbilical cord MSCs (UC-MSCs), bone marrow MSCs (BMSCs), hematopoietic MSCs (HMSCs), and dental root MSCs (DT-MSCs) [31]. AD-MSCs, primarily produced from fat tissues that have been surgically removed or separated after liposuction are the most common and easily attainable source of MSCs. UC-MSCs, closest to human embryonic stem cells, have the most regeneration potential. BMSCs, isolated from the bone marrow cavity, have low immunogenicity, their acquisition process is unstable and difficult. These MSCs were initially employed to treat injuries and fractures [32], autoimmune disorders [33], and malignant tumors [34]. Besides, several types of MSCs have been recently used in hair experimentation. MSCs can secrete various growth factors, chemokines, and cytokines, thus improving healing and hair growth. Notably, MSCs from different sources have different surface antigens [35].
Zhang et al. injected BMSCs and conditioned medium (CM) into the back skin of a mouse model and found that Krt25, Cpm, Stmn1, and Mb may be crucial in the BMSC-induced changes of the HF through quantitative proteome analysis employing tandem mass tags [36]. BMSCs can also induce immune tolerance and anti-inflammatory effects in severe AA patients, which differentiate CD3+Foxp1+ regulatory T cells in peripheral blood mononuclear cells and inhibit mRNA levels of proinflammatory cytokines [37]. HMSCs can also improve DPC's viability and activate several molecules in the Wnt pathway (phosphorylated GSK-3β and β-catenin), decrease the levels of Wnt pathway inhibitors (Dickkopf-1 (DKK-1), caspase-1, caspase-3, and IFN-γ receptor) [38]. In addition, HMSCs can reduce inflammatory alterations and growth inhibition of IFN-induced ORS by suppressing the Janus tyrosine kinase (JAK)/STAT pathway [39].
Activation of hypoxia-inducible factors 1α (HIF-1α) and extracellular regulated protein kinases1/2 signaling pathways can regulate paracrine levels in AD-MSCs under hypoxic microenvironment, thus enhancing DPC's proliferation and inducing follicle formation through CM [40]. Furthermore, adipose-derived stromal vascular cells (ADSVCs) are pluripotent stem cells found in adipose tissue. Anderi et al. administered ADSVCs to the scalps of 20 AA patients and showed that ADSVCs can also increase hair density, hair diameter, and resistance to tensile testing [41]. Stromal vascular fraction (SVF) separated from fat includes stem cells, blood vessels, and immune cells. Studies have shown that autotransplantation of SVF can significantly increases hair density and improve scalp keratin score in AGA patients [42].
Also, UC-MSCs can inhibit DKK-1 production, induce the secretion of vascular endothelial growth factor, and protect keratinocytes and DPCs in HFs [43]. Wharton's Jelly MSCs (WJ-MSCs) are found in the umbilical cord. Czarnecka et al. treated the alopecia site of AA patients through a single injection of WJ-MSCs and found an average of 52.2% hair regeneration after 3 months [44]. However, only a few reports have assessed the effects of embryonic MSCs (EMSCs) on hair. Jeon SH showed that EMSCs can accelerate the transition from the telogen to the anagen in C3H mice, and are more effective than AD-MSCs. EMSCs can overexpress NADPH oxidase 4, produce reactive oxygen species, and upregulate fibroblast growth factor 7, interleukin-12B, and teratocarcinoma-derived growth factor 1 under hypoxia (2% O2) environments, thereby inducing the transition of hair cycle from the telogen to the anagen phase [45].
Stem cells from human exfoliated deciduous teeth (SHED) have attracted much interest due to easy accessibility, low immunogenicity, and lack of ethical constraints. SHED-MSCs can upregulate Shh and glioma-associated oncogene 1 (Gli1) in mouse skin, thus promoting hair growth by enhancing the Shh/Gli1 pathway [46]. In one study, SHED-MSCs-CM were injected in 33 male subjects with AGA (six times every month), and showed effectiveness in 75% of the subjects [47]. The effectiveness of SHED-MSCs-CM was not significantly related to the severity of the condition, age, and the use of dihydrotestosterone synthesis inhibitors. However, the effectiveness was related to the Hamilton Norwood Classification level, and maximum hair diameter, villus rate, and multiple HF unit rate are quantitative hair microscopy evaluation indexes.
However, the best MSCs for the treatment of alopecia are unknown. Nonetheless, the processes for harvesting, culturing, and differentiating MSCs should be improved to enhance the use of MSCs for the treatment of various diseases and regenerating tissue.
Exosomes
Exos are lipid bilayer membrane vesicles that are generated by cells. Exos contain lipids, proteins, mRNAs, microRNAs (miRNAs), and other signaling components and have a key role in the paracrine secretion of MSCs [48]. Exos enter target cells through endocytosis or direct fusion to regulate the immune system, enhance cell migration and function, regenerate damaged and apoptotic cells, grow new blood vessels and neurons, and reduce inflammation by inducing a complex chain of biochemical reactions [4]. Exos, microvesicles or microparticles, and apoptotic bodies are considered as external vesicles (EVs). However, most studies have focused on Exos since it is more closely associated with skin and hair regeneration.
MSCs-EVs can directly produce active Wnt factors, thus activating the Wnt pathway and regenerating skin and HF. As a result, MSCs-EVs participate in immune regulation of immune diseases through the secretome, such as AA [49]. MSCs-EVs can also rescue H2O2-induced DPCs' apoptosis, promote their proliferation and migration, and significantly reduce proinflammatory TNF-α signaling pathway [50]. Furthermore, MSCs-EVs can upregulate Axin2, EP2, and LEF1 (the target transcription factors of β-catenin in Wnt pathway), translocate β-catenin into nuclei of DPCs (to promote migration of ORS cells), and enhance the expression of keratin (K) differentiation markers (K6, K16, K17, and K75) in ORS cells [51].
Nonetheless, the apoptotic cells after extensive apoptosis during MSC transplantation are not without benefits. These cells secrete EVs also containing miRNAs, proteomics, and functional factors with regenerative ability [52]. The Wnt/β-catenin signaling pathway is activated during this process. Notably, the mechanical forces of MSC–EV migration from the dermis to epidermis are closely related to the level of the endogenous inhibitor of Wnt signaling (DKK-1). Treadmill exercise can decrease DKK-1 expression, and the number of MSCs–EVs migrating to the stratum corneum increases [53].
MiRNAs, small noncoding RNA molecules, are related to Exo function. Target RNA is linked to miRNAs, which cause its degradation and further prevent protein translation. Besides, miRNAs can orchestrate hair regeneration. Studies have shown that multiple miRNAs, such as miR-122-5p [54], miR-181a-5p [55], miR-200 [56], and miR-218-5p [57] can promote hair regeneration by separately targeting the Wnt/β-catenin signaling pathway or the TGF-β/Smad signaling pathway. Besides, miRNAs can promote the differentiation and movement of hair germ cells by acting on the interfollicular epidermis.
Compared with MSCs, Exos have a small diameter, strong penetration, and a reduced chance of causing immune rejection, and thus are easily accepted by medical ethics. Therefore, Exos have greater development and value in participating in research on controlling hair cycle and growth. Although numerous studies have shown that Exos regulate hair cycle and regeneration, there is no systematic consensus.
Conditioned medium
MSCs-CM contains growth factors and secretions made by MSCs. Some experts suggest that MSCs-CM can be a cell therapy because it works through paracrine secretion theory. For example, human amniotic fluid MSCs (AF-MSCs) CM can stimulate hair regrowth. AF-MSCs with highly expressed Nanog genes can secrete several secretory molecules, such as insulin-like growth factor (IGF), platelet-derived growth factor (PDGF), basic fibroblast growth factor (bFGF), and Wnt7a related to hair regeneration and growth into MSCs-CM, thus increasing DPCs' activity and HF regeneration [58]. Oh et al. [59] showed that MSCs-CM can be optimized by adding TGF-β1 and an agonist of canonical Wnt signaling lithium chloride to normal UC-MSCs, which is called P-CM. The macrophage migration inhibitory factor in P-CM can promote hair growth through β-catenin and p-GSK-3β signaling pathway. MSCs-CM can also promote the development of a 3D gas–liquid culture system that simulates HFs. MSCs-CM with highly expressed Wnt can maintain hair induction ability of DPCs in expanded culture.
The treated DPCs and epidermal cells can then be inoculated into collagen chitosan (CS) scaffold cells, forming dermal aggregates and proliferative cell layers, thus inducing hair regeneration in nude mice [60].
Although MSCs-CM can effectively prevent safety issues, such as immune response and infection, any remaining cell debris or fragments limit MSCs-CM application. Therefore, more effective and standardized extraction techniques for MSCs-Exos and MSCs-CM should be established. Notably, some drug and cosmetic companies are working closely with researchers in this area to promote MSCs' transformation and their related products.
Signal Pathways Related to Hair Regeneration
Wnt/β-catenin signaling pathway
The Wnt/β-catenin signaling pathway is crucial for HF regeneration. Several Wnt proteins that are released after activation of the Wnt/β-catenin signaling pathway, including Wnt1a, Wnt3a, and Wnt10a/b, affect the hair cycle and regeneration, thus repairing damaged HFs. Wnt4 can promote hair growth by increasing the expression of growth factors. Wnt5a regulates the morphology of HFs during early development [61].
β-catenin is a core signal transduction factor of the typical Wnt signaling pathway. However, Wnt can regulate the transcription of multiple genes by inhibiting β-catenin degradation. Nonetheless, Han et al. showed that β-catenin is excessively activated during the catagen phase, thus promoting the differentiation of CD34+DSCs and the overactivation of HF fibroblasts. This results in HF aging and the loss of potential in DPCs and their stem cells [62]. Other signaling pathways interact with the Wnt signaling pathway during this process to regulate hair cycle. For example, androgen can downregulate BMP2 and BMP4 in DPC spheroids, thereby affecting the accumulation of β-catenin and the activation of Wnt signaling pathways, resulting in HF-SC differentiation failure [63].
Studies have shown that a substance can stimulate hair regeneration and regulate hair cycle if it can activate the Wnt signaling pathway. KY19382 is one of the newly synthesized analogs of indirubin-3′-monoxime that can activate Wnt/β-catenin signaling in DPCs. KY19382 can increase alkaline phosphatase activity of DPCs and hair regeneration [64]. Recent studies have also shown that other compounds, such as Pyruvate Kinase M2 [65], Isoxazole 9 [66], and 5-Bromo-3,4-dihydroxybenzaldehyde [67], can affect the Wnt signaling pathway.
TGF-β/BMP signaling pathway
TGF-β family, including TGF-β, BMP, growth differentiation factor, anti-Mueller tube hormone, Activin, and Nodal, has attracted a lot of attention because of its numerous functions during cellular development and in many diseases, including cancer [68]. TGF-β plays a dual role in HF cell regulation, acting as an anti- or proapoptotic factor. However, TGF-β is influenced by BMP, TNF, and various HF growth cycle stages [69]. TGF-β1 is significantly overexpressed on the scalp of AGA patients. Besides, TGF-β1 can induce apoptosis of adjacent endothelial cells and vascular regression, leading to HF miniaturization [70], but TGF-β2 upregulates collagen, delays the aging of DPCs, and promotes the formation of spheroids in DPCs [71]. The glucocorticoid receiver and Forkhead box P3 costimulate the production of TGF-β3 by regulatory T cells, thereby activating Smad2/3 in HF-SCs and promoting HF-SCs' proliferation [72].
BMP is a multifunctional regulator of vertebrate development by binding to Smad1/5/8 in the nucleus. BMP also causes transcription of associated target genes, thus controlling the growth and differentiation of HF-SCs [2]. Smad4 is the intersection downstream of multiple TGF pathways, and significantly influences the growth and differentiation of HF. Calvo-Sánchez MI indicated that Endoglin (Eng) is a key coreceptor for BMP growth factors. The interaction between β-catenin and Smad4 can activate Eng promoter transcription, completing the switching mechanism of hair cycle [73].
However, the effect of TGF-β/BMP signaling pathway on hair is not unidirectional, necessitating further studies.
PI3K/AKT signaling pathway
PI3K
Mammalian target of rapamycin (mTOR) mainly targets serine/threonine protein kinase. mTOR complex 2 (mTORC2) controls HF-SCs' metabolism and the reversibility of ORS state through AKT. Glutamine metabolism and PI3K/AKT signaling pathways are inhibited when mTORC2 is absent, leading to failure in HF-SCs' niche reconstruction (HF regeneration defects) [78]. Furthermore, PI3K/AKT/mTOR/FOX01 signaling pathway is activated when HF-MSCs are stimulated by a micro current (50 μA), thus promoting the activity of DPCs' proliferation and migration [79]. In addition, BMP regulates the PI3K/AKT signaling pathway, thus inhibiting downstream PI3K/AKT signaling by activating Phosphatase and Tensin Homolog deleted on Chromosome 10 (PTEN). Sunkara et al. demonstrated that the signaling cascade of BMP-AKT-GSK3β can regulate the effect on the level of activity β-catenin through BMP signaling agonists [80].
Hedgehog signaling pathway
Hedgehog signaling reprograms HF niche fibroblasts to a hyperactive state that regulates DP fibroblasts. This state considerably speeds up hair growth and promotes HF proliferation in mice [81]. Hedgehog can regulate mesenchymal niche function in HF through SCUBE3/TGF-β mechanism when normal mice are injected with SCUBE3 protein (ligand for nonconventional TGF-β). Hedgehog signaling also induces new HFs in adjacent epithelial and stromal cells by driving Wnt/β-catenin signaling. In contrast, Hedgehog pathway activation alone in epithelial or stromal cells cannot result in new HF [82]. HF-SCs differentiate to stratum corneum with a large amount of reactive oxygen when mice are fed a high-fat diet. In this process, Hedgehog signaling is inhibited and autocrine/paracrine IL-1R signaling and NF-κB activate. Thus, miniaturized HF and lipid-rich HF-SCs form [83].
HF is a micro-organ with a complex regulatory network, including Wnt/β-catenin signaling pathway, TGF-β/BMP signaling pathway, Notch signaling pathway, and Hedgehog signaling pathway. There may be other pathways involved that we currently do not understand. During hair cycle and growth process, the Wnt signaling pathway is the main positive effect, while TGF-β is opposite. The studies on the interrelationships between various signal pathways are still in the initial stage and require more in-depth research.
New Methods and Materials of MSCs for Hair Treatment
The treatment of various alopecia diseases with MSCs and their derivatives has entered the clinical stage. Currently, the commonly used treatment methods include local medication or subcutaneous injection. In the clinical trials conducted by Gan et al. [84], MSC suspensions extracted from healthy HFs of AGA patients themselves were administered through injection. They observed an increase in the proportion of terminal hair and hair shaft diameter at 1 month, and enhanced miniaturized HF with hair shaft diameter less than 60 μm. In addition, many researchers have developed several new techniques, and materials, which have been developed based on that, can be used with MSCs.
Hydrogels are biomaterials that provide suitable niches for cell adhesion and survival. Xu H [85] constructed a functional injectable thermosensitive hydrogel for encapsulating UC-MSCs with CS, glycerophosphate sodium (GP), and cellulose nanocrystal (CNC). In addition to accelerating the gel speed, incorporating CNC in the CS/GP system enhanced the mechanical characteristics of the gel and slowed its rate of degradation. This brand-new hydrogel can be injected and does not cause toxic effects. When applied in the treatment of full-thickness skin wounds, it can accelerate wound closure, microcirculation, tissue remodeling, epithelial regeneration, and HF regeneration. It can also alleviate excessive inflammation in the central and surrounding wounds. A recent study investigated the incorporation of PDGF-BB and BMSCs into an injectable hydrogel made of sodium alginate and dextran. The formulation exhibited good biocompatibility and continuous release [86].
Silk fibroin in silk is regarded as a matrix that can be exploited for tissue regeneration, due to its high biocompatibility, changeable mechanical qualities, adjustable degradation rate, simplicity of modification into diverse various material forms, and limited inflammatory properties [87]. Li created a silk nanofiber hydrogel for BMSCs' loading to promote wound healing and hair regrowth [88]. A suitable environment for the functional repair of wounds was created by the synergistic interaction between MSCs and the silk-based composite hydrogel, which enhanced angiogenesis and the M1-M2 phenotypic conversion in macrophages. Furthermore, composite hydrogel with MSCs promoted rapid healing of intact HF without scarring.
Some scholars have established a system of improving drug delivery, which is a detachable microneedle patch [89], mainly composed of hair-derived keratin, for continuous delivery of MSC-derived secretions and a small-molecule drug, UK5099 (an activator of HF-SCs). The system improves the treatment efficacy and promotes pigmentation and hair regeneration with reduced doses. Yuan et al. [90] developed soluble microneedle patches that can produce less damaging mechanical stimulation, due to incorporation of MSCs-CM containing high levels of HIF-1α. For this reason, the AGA mouse model exhibits faster hair regeneration relative to minoxidil when the frequency of administration is low. Although this administration method is not as direct as intradermal injection and has high absorption efficiency, it can reduce the pain associated with scalp injection and increase compliance.
Experts have attempted to stimulate the proliferation of DP and the growth of HF by replicating the 3D microenvironment of HF. This has been achieved through methods such as the utilization of 3D DP spheres within a silk–gelatin microenvironment [91], the incorporation of MSCs within 3D bioprinting scaffolds [92], and the creation of 3D microgel models using a combination of poly (ethylene glycol) diacrylate and methacrylate gelatin [93]. For hair injuries caused by wounds, collagen chitosan scaffolds containing BMSCs with IGF 1 may speed up skin wound healing and successfully regenerate HF [94].
These pioneering materials and drug delivery techniques are the result of collaborative efforts across multiple disciplines. They represent significant advancements in the medical field, expanding the potential of stem cell therapy.
Summary
MSCs are increasingly applied in medicine and in the field of dermatology owing to their capacity to provide immunological control, improve repair processes, and promote autophagy.
The main breakthrough in hair-related research lies in the transition of hair cycle. Rapid and effective identification of hair and the characterized cell distribution in HF at different stages is the basis for future research. The effectiveness of MSCs in enhancing hair regeneration has been documented, but the signaling pathways responsible for this effect require further investigation. Exploring whether MSCs from various sources operate through similar or distinct signaling networks would also be an effort worth further research.
However, because of the complex microenvironment, research on the function of MSCs in hair and HFs is still in its foundational stage and faces several challenges, for example, the difficulty of extracting and culturing HF-MSCs; mouse hair cannot accurately simulate the growth laws and signaling pathways of human hair, source selection, and immunogenicity resolution of MSCs. Further discussion and resolution are needed regarding the administration methods and treatment efficiency of MSCs.
Footnotes
Author Disclosure Statement
The author declares that there are no competing financial interests.
Funding Information
This review was supported by grants to Xuegang Xu from the National Natural Science Foundation of China [81972956, 82173443], the Shenyang Youth Science and Technology Innovation Talent Support Program [RC220268], and the Dermatology and Immunology Laboratory of the First Affiliated Hospital of China Medical University. The authors thank Home for Researchers editorial team (