
Abstract
Feeder cells play a crucial role in maintaining the pluripotency of embryonic stem cells (ESCs) by secreting various extrinsic regulators, such as extracellular matrix (ECM) proteins and growth factors. Although primary mouse embryonic fibroblasts (MEFs) are the most widely used feeder cell type for the culture of ESCs, they have inevitable disadvantages such as batch-to-batch variation and labor-intensive isolation processes. Here, we revealed that the Sandoz inbred Swiss Mouse (SIM) thioguanine-resistant ouabain-resistant (STO) cell line, an immortalized cell line established from mouse SIM embryonic fibroblasts, can be used as a feeder layer for in vitro culture of authentic pig ESCs instead of primary MEFs. First, the expression of genes encoding ECM proteins and growth factors was analyzed to compare their secretory functions as feeder cells. Quantitative real-time polymerase chain reaction (qPCR) showed that the gene expression of these pluripotency-associated factors was downregulated in STO cells compared to primary MEFs of similar density. Therefore, subsequent optimization of the culture conditions was attempted using higher STO cell densities. Notably, pig ESCs cultured on STO cell density of 3 × (187,500 cells/cm2) exhibited the most similar pluripotent state to pig ESCs cultured on primary MEF density of 1 × (62,500 cells/cm2), as determined by alkaline phosphatase staining, qPCR, and immunocytochemistry. In addition, pig ESCs cultured on STO cell density of 3 × formed complex teratoma containing multiple types of tissues derived from all three germ layers. Our culture conditions using optimal STO cell density can be applied to fields requiring reproducible and scalable production of pig ESCs, such as preclinical research and cellular agriculture.
Introduction
Embryonic stem cells (ESCs) are pluripotent stem cells (PSCs) derived from the inner cell mass of blastocysts [1,2]. They are capable of unlimited self-renewal and have the ability to differentiate into various cell types from all three germ layers [3]. In addition to human ESCs, ESCs have also been derived from animals such as pigs, cows, and sheep [4,5]. Because pig ESCs have the potential for biomedical and agricultural applications [6], numerous studies have attempted to establish stable ESC lines from pigs [7 –11]. Recently, authentic pig ESCs were derived and maintained by defining the essential factors for pluripotency in pigs [12]. Derived pig ESCs showed single-layered and flattened colony morphology and expressed the pluripotency markers OCT4, SOX2, NANOG, SSEA1, SSEA4, TRA-1-60, and TRA-1-81.
Feeder cells play an important role in maintaining ESC pluripotency by secreting various extrinsic regulators, including extracellular matrix (ECM) proteins and growth factors [13]. ECM proteins create complex adhesive structures that affect cell survival, growth, proliferation, and differentiation. Major ECM proteins, including collagen, laminin, and fibronectin, have been used as substrates for ESC culture, showing that these ECM proteins support attachment and self-renewal of ESCs [14,15]. In addition, growth factors also support sustaining the undifferentiated state of ESCs. Since the basic fibroblast growth factor (bFGF) was found to be the first key growth factor essential for the pluripotency of human ESCs [16], it was revealed that signaling molecules such as transforming growth factor beta (TGFβ), and activin A (ActA) also serve as key growth factors [17]. Therefore, various combinations of bFGF, TGFβ, and ActA have been repeatedly supplemented to the culture media of human ESCs to enhance their undifferentiated proliferation [18,19]. Moreover, noggin, an antagonist of the bone morphogenetic protein (BMP) family, cooperates with bFGF to suppress the differentiation of human ESCs [20,21].
The most widely used feeder cell type for ESC culture is primary mouse embryonic fibroblasts (MEFs) isolated from mouse fetuses [22,23]. Because primary MEFs have limited proliferative ability [24], repetitive production is required to utilize them as feeder cells. Accordingly, batch-to-batch variations can hamper experiment reproducibility. Also, the primary culture process, in which they are isolated, is labor-intensive, which limits the scalable culture of ESCs. For this reason, Sandoz inbred Swiss Mouse (SIM) thioguanine-resistant ouabain-resistant (STO) cell line, an immortalized cell line established from mouse SIM embryonic fibroblasts [25], has been used as an alternative feeder cell line for ESC culture of various species [26 –28]. In this study, we attempted to develop reproducible and scalable culture conditions for pig ESCs using the STO cell line as a feeder layer.
Materials and Methods
Animal care
The care and experimental use of mice were approved by the Institutional Animal Care and Use Committee of Seoul National University (Approval No. SNU-191025-4-4 and SNU-201019-1-4). Pregnant Institute of Cancer Research (ICR) and athymic nude mice were purchased from Samtaco Bio (Korea) and OrientBio (Korea), respectively. Mice were maintained according to the standard protocol of the Institute of Laboratory Animal Resources at Seoul National University.
Culture of mouse feeder cells
Primary MEFs were isolated from ICR fetuses according to a previous study [29]. The STO cell line was purchased from the American Type Culture Collection. STO cells were cryopreserved after five times passaging. The thawed STO cells were used in experiments after two to three times subculture. Primary MEFs and STO cells were expanded in Dulbecco's modified Eagle's medium (DMEM; Welgene, Gyeongsan, Korea) supplemented with 10% fetal bovine serum (FBS; collected and processed in the USA; Genedepot, Katy, TX), 1 × GlutaMAX (Gibco, Gaithersburg, MD), 0.1 mM β-mercaptoethanol (Gibco), and 1 × antibiotic/antimycotic (Gibco). Primary MEFs and STO cells were mitotically inactivated by mitomycin C treatment (Roche, Basel, Switzerland) for 3 h and plated at a density of 62,500 cells/cm2 in the control group (1 × ).
Culture of pig ESCs
Pig ESCs were cultured in ESC medium as previously described [12]. ESC medium [DMEM/F12 containing 15% (v/v) KSR, 0.1% (v/v) LC, 1 × GlutaMAX, 0.1 mM β-mercaptoethanol, 1 × minimal essential medium (MEM) nonessential amino acids, and 1 × antibiotic-antimycotic; all from Gibco] supplemented with 20 ng/mL FGF2 (R&D Systems, Minneapolis, MN), 5 ng/mL ActA (R&D Systems), 1.5 μM CH (Cayman Chemical, Ahn Arbor, MI), and 2.5 μM IWR-1 (Sigma-Aldrich, St. Louis, MO) was changed every 24 h. Pig ESCs were cultured under humidified conditions in an atmosphere containing 5% CO2 at 37°C and subcultured every 4–5 days.
Quantitative real-time polymerase chain reaction
Total RNA was extracted from the cells using TRIzol® reagent (Invitrogen, Waltham, MA) according to the manufacturer's instructions. cDNA was synthesized using the High Capacity RNA-to-cDNA Kit (Applied Biosystems, Waltham, MA), producing a final volume of 20 μL. The derived cDNA samples were amplified with PowerSYBR® Green PCR Master Mix (Applied Biosystems) containing 0.5 pmol of each primer set (Table 1) in a 10 μL reaction volume. Amplification and detection were conducted using the ABI 7300 Real-Time PCR system (Applied Biosystems) under the following conditions: one cycle of 50°C for 2 min and 95°C for 10 min, followed by 40 cycles of denaturation at 95°C for 15 s and annealing/extension at 60°C for 1 min. The relative expression level was calculated by normalizing the threshold cycle (Ct) values of each gene to that of GAPDH via the Δ−Ct method [30].
Primer Sets Used for Quantitative Real-Time Polymerase Chain Reaction
Alkaline phosphatase staining
Before staining, all cell samples were fixed in 4% (w/v) paraformaldehyde for 15 min at 4°C. The fixed cells were stained with a solution containing nitro blue tetrazolium chloride and 5-bromo-4-chloro-3-indolyl phosphate toluidine salt stock solution (Roche) in a buffer solution for 30 min at room temperature (RT).
Immunocytochemistry
Before staining, all cell samples were fixed in 4% (w/v) paraformaldehyde for 15 min at 4°C. Fixed cells were permeabilized using 0.2% Triton-X100 (Sigma-Aldrich) for 15 min at RT and then blocked using 10% (v/v) goat serum in Dulbecco's phosphate-buffered saline for 2 h at RT. Serum-treated samples were incubated overnight at 4°C with primary antibodies: mouse anti-SSEA1 (1:200, MAB4304; Millipore, Burlington, MA), mouse anti-SSEA4 (1:200, MAB4301; Millipore), chicken anti-OCT4 (1:200, ab134218; Abcam, Cambridge, United Kingdom), rabbit anti-SOX2 (1:200, AB5603; Millipore), and rabbit anti-NANOG (1:200, 500-P236; PeproTech, Rocky Hill, NJ). Then, the cells were treated with Alexa Fluor-conjugated secondary antibodies for 2 h at RT. Nuclei were stained with Hoechst33342 (Molecular Probes, Eugene, OR). Images of the stained cells were captured using a TE2000-U inverted microscope (Nikon, Tokyo, Japan) and processed using ImageJ software.
Karyotyping
Karyotyping of cells using standard G-banding chromosome and cytogenetic analysis was performed at GenDix Laboratories (Korea).
Spontaneous differentiation using the embryonic body method
Pig ESCs were dissociated into single cells using TrypLE Express (Gibco) and cultured on ultralow-attachment plates (Sigma-Aldrich) with DMEM supplemented with 10% (v/v) FBS (Genedepot), 1 × GlutaMAX (Gibco), 0.1 mM β-mercaptoethanol (Gibco), and 1 × antibiotic/antimycotic (Gibco) for 5 days. After suspension culture, the dissociated cells aggregated and formed embryonic bodies (EBs), which were seeded on 0.1% (w/v) gelatin-coated plates and cultured for 7 days in the same medium.
Teratoma formation assay
Approximately 5–10 × 106 ESCs were resuspended in 200 μL of ESC culture medium containing 50% (v/v) BD Matrigel Matrix (BD Biosciences, Franklin Lakes, NJ). Next, resuspended pig ESCs were injected subcutaneously into 4-week-old athymic nude mice (Orient Bio). Three months after transplantation, the teratoma was collected, fixed in 4% (w/v) paraformaldehyde, embedded in paraffin, and stained with hematoxylin and eosin for light microscopic examination.
Statistical analysis
The data are presented as the mean ± the standard error of the mean and were analyzed using Prism 6 software (GraphPad Software, San Diego, CA). Significant differences among experimental groups were determined by one-way analysis of variance (ANOVA), followed by Fisher's least significant difference test, whereas unpaired t-tests were used for binomial data. Differences were considered significant at P < 0.05.
Results
Comparative analysis of primary MEFs and STO cells as feeder cells
Feeder cells create a suitable microenvironment for ESCs by secreting extrinsic regulators associated with pluripotency, including ECM proteins and growth factors. ECM proteins support the attachment and self-renewal of ESCs, and growth factors play an important role in sustaining the undifferentiated state of ESCs [17,31]. To compare their secretory functions as feeder cells, the gene expression of pluripotency-associated factors in primary MEFs and STO cells was analyzed. Col1a1, Col1a2, Fn1, Lama1, Lama2, and Lama4 were selected to compare the gene expression of ECM proteins, and all of these genes were downregulated in STO cells compared to primary MEFs (Fig. 1A). In particular, Lama1 and Lama2 showed much lower expression in STO cells compared to primary MEFs. Fgf2, Tgfb1, Inhba, and Nog were selected to compare the gene expression of growth factors. Similar to genes encoding ECM proteins, Fgf2 and Inhba were downregulated in STO cells compared to primary MEFs (Fig. 1B). However, the expression levels of Tgfb1 and Nog were similar in both feeder cell types. Overall, the expression patterns of pluripotency-associated factors were downregulated in STO cells compared to primary MEFs at a similar density.
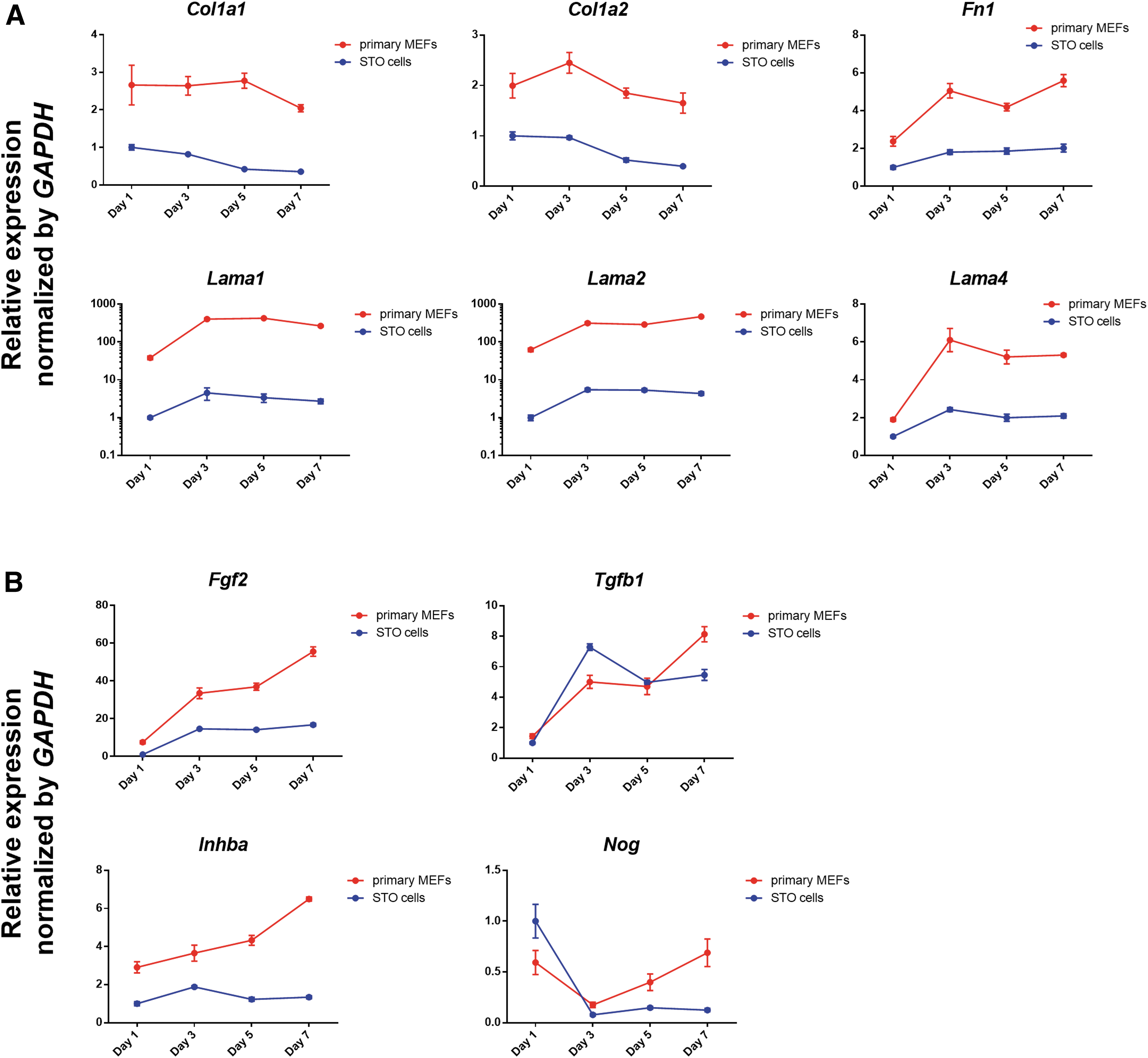
Comparative analysis of primary MEFs and STO cells.
Optimization of STO cell density for in vitro culture of pig ESCs
Based on these results, subsequent optimization of the culture conditions was attempted with higher STO cell densities. To identify optimal STO cell density, pig ESCs were cultured on STO cells at several densities and compared with pig ESCs cultured on primary MEF density of 1 × (MEF 1 × -pESCs). Pig ESCs cultured on STO cell density of 1 × (STO 1 × -pESCs) formed loose colonies with undefined boundaries that were markedly different from the colonies of MEF 1 × -pESCs (Fig. 2A). Interestingly, upon increasing the STO cell density, the colonies of pig ESCs became more compact and defined. In particular, the colony morphology of pig ESCs cultured on STO cell density of 3 × (STO 3 × -pESCs) was the most similar to that of MEF 1 × -pESCs. Pig ESCs cultured on STO cell density of 4 × formed relatively smaller colonies than MEF 1 × -pESCs, and the colony size did not surpass a certain value during the culture period. Consistent with the morphological observations, alkaline phosphatase (AP) activity was the lowest in STO 1 × -pESCs and the most similar to that in MEF 1 × -pESCs in STO 3 × -pESCs. The expression of pluripotency markers and three germ layer markers were analyzed using quantitative real-time polymerase chain reaction (qPCR) (Fig. 2B). In expressions of most genes, no considerable difference was observed among MEF 1 × -pESCs and pig ESCs cultured on STO cells at several densities. However, the BMP4 expression level was significantly upregulated in STO 1 × -pESCs, indicating mesodermal differentiation. In addition, the pluripotency marker OCT4, which promotes ESCs toward the mesodermal lineage [32], was mildly upregulated in STO 1 × -pESCs. Immunostaining revealed that most pluripotency markers were similarly expressed in MEF 1 × -pESCs and pig ESCs cultured on STO cells at several densities (Fig. 2C). However, the expression pattern of SSEA4 differed. The number of SSEA4-expressing cells was the lowest in STO 1 × -pESCs, and the growth of these cells was enhanced by increasing the density of STO cells. In particular, SSEA4 expression in STO 3 × -pESCs was the most similar to that in MEF 1 × -pESCs. Overall, STO 1 × -pESCs gradually lost their stemness, and STO 3 × -pESCs exhibited the most similar characteristics to MEF 1 × -pESCs.
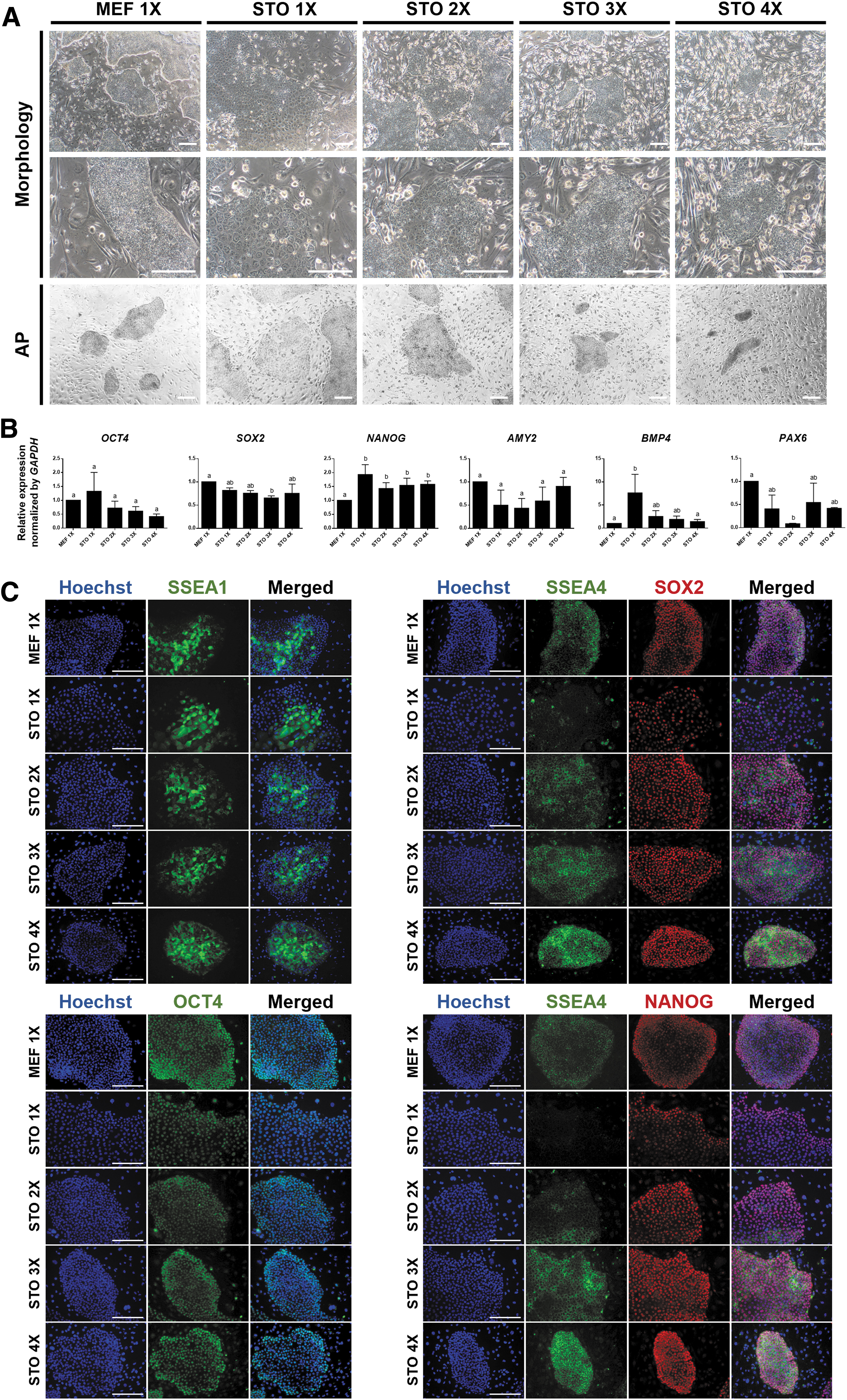
Optimization of STO cell density for pig ESCs. STO cells were plated at several densities, and then pig ESCs were cultured on STO cells at several densities. Pig ESCs cultured in primary MEF 1 × were used as the control group.
As a further study, the expression of additional mesoderm markers was analyzed to confirm mesodermal differentiation of STO 1 × -pESCs (Fig. 3A). Consistent with the above results, T was significantly upregulated in STO 1 × -pESCs. In addition, DES was slightly upregulated in STO 1 × -pESCs. Based on a previous study revealing that BMP signaling promotes mesodermal commitment of ESCs [33], the gene expression of BMP inhibitors in primary MEFs and STO cells was compared (Fig. 3B). Nog, Chrd, Grem1, and Grem2 were selected for comparison. Most BMP inhibitors, except Grem1, were significantly downregulated in STO cells. To validate whether BMP signaling induces mesodermal differentiation of pig ESCs, MEF 1 × -pESCs were treated with BMP4. The control group showed compact colonies of pig ESCs, whereas relatively loose colonies were observed in the BMP4 group (Fig. 3C). Furthermore, OCT4 and T were significantly upregulated in the BMP4 group (Fig. 3D). Next, STO 1 × -pESCs were treated with LDN193189, a potent BMP inhibitor, to investigate whether BMP signaling suppression can prevent mesodermal differentiation of pig ESCs. Although colony morphology was similar in both groups (Fig. 3E), OCT4 was significantly downregulated in the LDN group (Fig. 3F). In addition, T was mildly downregulated in the LDN group. These results indicate that the differentiation of STO 1 × -pESCs into the mesodermal lineage is possibly due to the insufficient secretion of BMP inhibitors.

Mesodermal differentiation of STO 1 × -pESCs.
Analysis of pluripotency and differentiation of STO 3 × -pESCs
Because STO 3 × -pESCs were the most similar to MEF 1 × -pESCs in various aspects, 3 × density was used as the standard STO cell density for further analysis. STO 3 × -pESCs formed compact colonies with high nuclear-to-cytoplasmic ratios and exhibited high AP activity (Fig. 4A). The expression of pluripotency markers in STO 3 × -pESCs was higher than pig embryonic fibroblasts (PEFs) and similar to that in MEF 1 × -pESCs (Fig. 4B). STO 3 × -pESCs stably maintained a normal karyotype (36+XX) (Fig. 4C). Immunostaining revealed that STO 3 × -pESCs expressed pluripotency markers at the protein level (Fig. 4D). To examine in vitro differentiation ability, EBs were generated from STO 3 × -pESCs (Fig. 5A). EBs spontaneously differentiated into multiple cell types, and during differentiation, three germ layer markers were upregulated, whereas pluripotency markers were downregulated (Fig. 5B). Next, to evaluate the in vivo differentiation ability, STO 3 × -pESCs were subcutaneously injected into an athymic nude mouse. STO 3 × -pESCs formed complex teratoma in mouse, and a teratoma with ∼0.5 cm was collected. Histological analysis showed that the teratoma contained multiple types of tissues derived from all three germ layers, including stratified nervous tissue (ectoderm), connective tissue (mesoderm), and respiratory epithelium with cilia (endoderm) (Fig. 5C). Taken together, we verified that an STO cell density of 3 × can maintain the pluripotent state and differentiation capacity of pig ESCs.

Analysis of the pluripotency of STO 3 × -pESCs.

Analysis of the differentiation of STO 3 × -pESCs.
Discussion
The role of feeder cells in maintaining pig pluripotency in vitro
Feeder cells play a crucial role in the maintenance of ESCs through the secretion of various paracrine factors and physical contact. The most widely used feeder cell type for ESC culture is primary MEFs isolated from mouse fetuses. Recently, authentic pig ESCs were derived using primary MEFs as feeder cells [12]. Because of their inherent properties related to primary culture, the use of primary MEFs as feeder layers can hamper the reproducible and scalable production of pig ESCs. Therefore, in this study, we demonstrated that the STO cell line can be used as a feeder layer for in vitro maintenance of authentic pig ESCs instead of primary MEFs.
Several studies have compared the expression and secretion of pluripotency-associated factors among various types of feeder cells and elucidated their respective characteristics [34 –36]. In our study, genes encoding pluripotency-associated factors, including Col1a1, Col1a2, Fn1, Lama1, Lama2, Lama4, Fgf2, and Inhba, were downregulated in STO cells compared to primary MEFs. Consequently, Pig ESCs cultured on STO cell density of 1 × (62,500 cells/cm2) differentiated into a mesodermal lineage. Consistent with these results, human ESCs cultured in culture medium (CM) derived from an STO cell density of 55,000 cells/cm2 underwent differentiation, whereas human ESCs cultured in CM derived from a similar density of primary MEFs were maintained [37]. These observations indicate that STO cell density of 1 × or less secrete insufficient amounts of pluripotency-associated factors and cannot sustain the undifferentiated state of ESCs.
BMPs prevent the differentiation of naive PSCs, including mouse ESCs, but stimulate the differentiation of primed PSCs, including human ESCs [33,38]. BMP signaling induces human ESCs into mesodermal derivatives [39 –42]. A previous study comparing primary MEFs and NIH/3T3 cells, which lack the ability to maintain human ESCs, revealed that primary MEFs express higher levels of BMP inhibitors, including noggin and gremlin [43]. Our results showed that genes encoding BMP antagonists were downregulated in STO cells compared to primary MEFs. Furthermore, when treated with a BMP inhibitor, genes involved in mesoderm development, including OCT4 and T, were downregulated in STO 1 × -pESCs. These findings suggest that BMP inhibitors secreted from feeder cells might play important roles in suppressing mesodermal differentiation during pig ESC culture.
Feeder-dependent/independent ESC culture for various applications
To avoid the use of primary MEFs, alternative feeder cells such as the STO cell line have been used to culture human ESCs [26,44]. However, similar to primary MEFs, STO cells contain murine factors that restrict the future clinical application of human ESCs [45]. Therefore, various types of human feeder cells, including fetal muscle and skin cells [46], neonatal foreskin fibroblasts [47], and adult marrow cells [48], have been used to eliminate xenogeneic contaminants. Because the use of a feeder layer hinders the elucidation of precise responses to various stimuli and raises concerns regarding unrecognized agents, feeder-free culture conditions for human ESCs have been developed [37,49,50].
Compared to humans, our understanding of the pluripotency network in pigs is relatively limited; thus, the development of xeno/feeder-free culture conditions for pig ESCs is still underway. Although PEFs have been used as feeder cells, in this case, pig ESCs are not maintained for longer than 11 passages [51]. Therefore, we used the STO cell line as the best alternative for replacing primary MEFs and found that an STO cell density of 3 × (187,500 cells/cm2) was the optimal density for maintaining the pluripotency of pig ESCs. At STO cell densities lower than 3 × , pig ESCs lost their stemness, possibly because of the factors mentioned above. Conversely, STO cell density higher than 3 × showed detrimental effects on the growth of colonies of pig ESCs. The reason for these effects may include rapid depletion or limited space for physical attachment [52].
The ultimate method for verifying ESC pluripotency is to confirm their in vivo developmental competence. The teratoma formation assay is considered the gold standard for assessing the differentiation potential of ESCs [53]. Here, STO 3 × -pESCs were capable of forming teratomas comprising all three germ layers. These results imply that our culture conditions reliably supported the pluripotency of pig ESCs. Taken together, we revealed that the STO cell line can be used as a feeder layer for in vitro culture of authentic pig ESCs instead of primary MEFs. Our culture conditions using optimal STO density can be applied to fields requiring reproducible and scalable production of pig ESCs, such as preclinical research and cellular agriculture.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the BK21 Four program, the Korea Evaluation Institute of Industrial Technology (KEIT) through the Alchemist project funded by the Ministry of Trade, Industry, and Energy (MOTIE; 20012411), and a National Research Foundation of Korea (NRF) grant funded by the government of the Republic of Korea (NRF-2023R1A2C1005026).