
Abstract
Neural stem/progenitor cells (NSPCs) are present in the mammalian brain throughout life and are involved in neurodevelopment and central nervous system repair. Although typical epigenetic signatures, including DNA methylation, histone modifications, and microRNAs, play a pivotal role in regulation of NSPCs, several of the epigenetic regulatory mechanisms of NSPCs remain unclear. Thus, defining a novel epigenetic feature of NSPCs is crucial for developing stem cell therapy to address neurologic disorders caused by injury. In this study, we aimed to define the R-loop, a three-stranded nucleic acid structure, as an epigenetic characteristic of NSPCs during neurodevelopment. Our results demonstrated that R-loop levels change dynamically throughout neurodevelopment. Cells with high levels of R-loops consistently decreased and were enriched in the area of neurogenesis. Additionally, these cells costained with SOX2 during neurodevelopment. Furthermore, these cells with high R-loop levels expressed Ki-67 and exhibited a high degree of overlap with the transcriptional activation markers, H3K4me3, ser5, and H3K27ac. These findings suggest that R-loops may serve as an epigenetic feature for transcriptional activation in NSPCs, indicating their role in gene expression regulation and neurogenesis.
Introduction
Neural stem/progenitor cells (NSPCs) persist in the mammalian brain throughout life. In the postnatal brain, NSPCs are spatially restricted to two specific regions: the subgranular zone (SGZ) within the dentate gyrus (DG) of the hippocampus and the subventricular zone (SVZ) in the lateral ventricles. 1 The emergence of a functional neural network necessitates NSPCs to produce an appropriate number of neurons and glial cells with the correct differentiation type at precise spatiotemporal locations, while malfunctioning of NSPCs can potentially result in the onset of various neurological disorders. 2
More importantly, recent studies indicate that NSPCs also participate in the repair and restoration of central nervous system (CNS) functions. 3 Thus, it becomes essential to comprehend the mechanisms that regulate proliferation and differentiation of NSPCs.
Epigenetics is imperative for regulating proliferation and differentiation of NSPCs by effectively and dynamically adjusting gene expression. 4,5 In addition to conventional mechanisms of epigenetics, such as DNA methylation, RNA interference, and histone modifications, recent studies have highlighted the significance of an epigenetic structure known as the R-loop in the regulation of transcriptional activity. 6,7
An R-loop is a three-stranded nucleic acid structure, comprising a DNA:RNA hybrid and the associated nontemplate single-stranded DNA. 8 It not only regulates gene transcription by interacting with DNA methylation enzymes and histone-modifying enzymes but also affects adjacent gene activity through spatial conformational changes. 9,10
Despite the R-loop playing substantial roles in transcriptional regulation, there exist differences in its functions among different cell types. The R-loop is associated with histone transcriptional activation markers, promoting transcriptional activation, while reports also indicate that R-loops can inhibit gene expression through various regulatory mechanisms. 11,12 In NSPCs, the R-loop exhibits enrichment at transcription start sites, gene bodies, and transcription end sites of actively transcribed genes, suggesting their involvement in regulatory processes. 13
Additionally, it is noteworthy that bulk R-loop levels escalate with age in Drosophila melanogaster photoreceptor neurons and this escalation is associated with a reduction in the expression of long highly expressed genes. 14 However, little is understood regarding the dynamic alterations in the R-loop during neural development.
In this study, we observed that NSPCs consistently display high levels of R-loops during brain development, and cells with high levels of R-loops colocalize with transcriptional activation and cell proliferation markers. This finding may provide new insights into the onset of neurodevelopment and offer an innovative epigenetic signature for discovering and characterizing key regulatory genes/regions for neurogenesis.
Materials and Methods
Animals
The C57BL/6N mouse strain was used for this study and all mouse experiments were approved by the Biomedical Ethics Committee of Health Science Center of Xi'an Jiaotong University (No. XJTUAE2022-613). This study conforms to the Declaration of Helsinki. The staff at the Laboratory Animal Center of Xi'an Jiaotong University is responsible for housing and daily maintenance.
Housing and environmental enrichment were according to general standards (12-h light–12-h dark cycle, food and water ad libitum, and at 20°C–24°C). All efforts were made to minimize animal suffering and keep the number of animals used to a minimum.
Tissue preparation
Embryonic day 18 (E18) and P5 mice were sacrificed by 65% CO2 exposure and their brains were dissected in cold phosphate-buffered saline (PBS) and then soaked in 4% paraformaldehyde (PFA) for fixation. P10, P15, P30, and P60 mice were anesthetized with 3.5% isoflurane, which was reduced to 2% for maintenance. The anesthesia machine was purchased from RWD Life Science (R640, China). Then, mice were transcardially perfused with normal saline, followed by 4% PFA.
All brains were dissected and postfixed by using 4% PFA for 5 days at 4°C, followed by paraffin embedding; 5-μM brain tissue serial slices were sectioned coronally using a microtome (RM2235; Leica, Germany), mounted onto adhesion microscope slides, and air-dried.
Immunofluorescence staining
The slides were deparaffinized with xylene, followed by rehydration in an EtOH gradient. Then, the slides were soaked in the antigen retrieval buffer (3 g of sodium citrate, 0.4 g of citric acid, and 1,000 mL of H2O, pH 6.0) by heating in a microwave oven at 95°C for 40 min. After washing with 0.01 M PBS (all washes were performed three times, 5 min each), slides were permeabilized with 0.3% Triton X-100 (X100-100ML; Sigma-Aldrich) for 15 min, rinsed, and then blocked in 5% bovine serum albumin (A8022; Sigma-Aldrich) with 5% normal goat serum (NS02L; Sigma-Aldrich).
The slides were incubated with primary antibodies (Supplementary Table S1) overnight at 4°C. The negative controls were incubated in a blocking buffer without the addition of primary antibodies. Following washing with 0.01 M PBS, samples were incubated with suitable secondary antibodies (Supplementary Table S1) for 2 h at room temperature.
The nuclei were visualized with a mounting medium, including DAPI (H-1200; Vector Laboratories). Images were captured using a Leica SP8 confocal microscope, and at least three animals were used for each experiment. Fluorescence intensity analysis and colocalization analysis were performed using ImageJ 5.0 software.
Results
High levels of R-loops in the neurogenesis area during mouse neurodevelopment
To characterize the dynamic alterations of R-loops during neurodevelopment, we collected mouse brain samples at various developmental stages: embryonic day 18 (E18), neonatal stage (postnatal days 5 and 10, P5 and P10), and postnatal days 15 and 30 (P15 and P30) in adolescence. Additionally, brain samples were collected from adult mice on postnatal day 60 (P60). Immunofluorescence staining was employed to assess R-loop levels and distribution.
The results showed a consistent reduction in the number of cells with high levels of R-loops during mouse neurodevelopment. Interestingly, there were consistently high levels of R-loops in cells present in both SVZ (Fig. 1A, C) and SGZ (Fig. 1B, D) during mouse neurodevelopment. Given that the majority of NSPCs, particularly in the adult brain, are located within the SVZ and SGZ, it is reasonable to postulate that NSPCs in mice manifest high levels of R-loops.
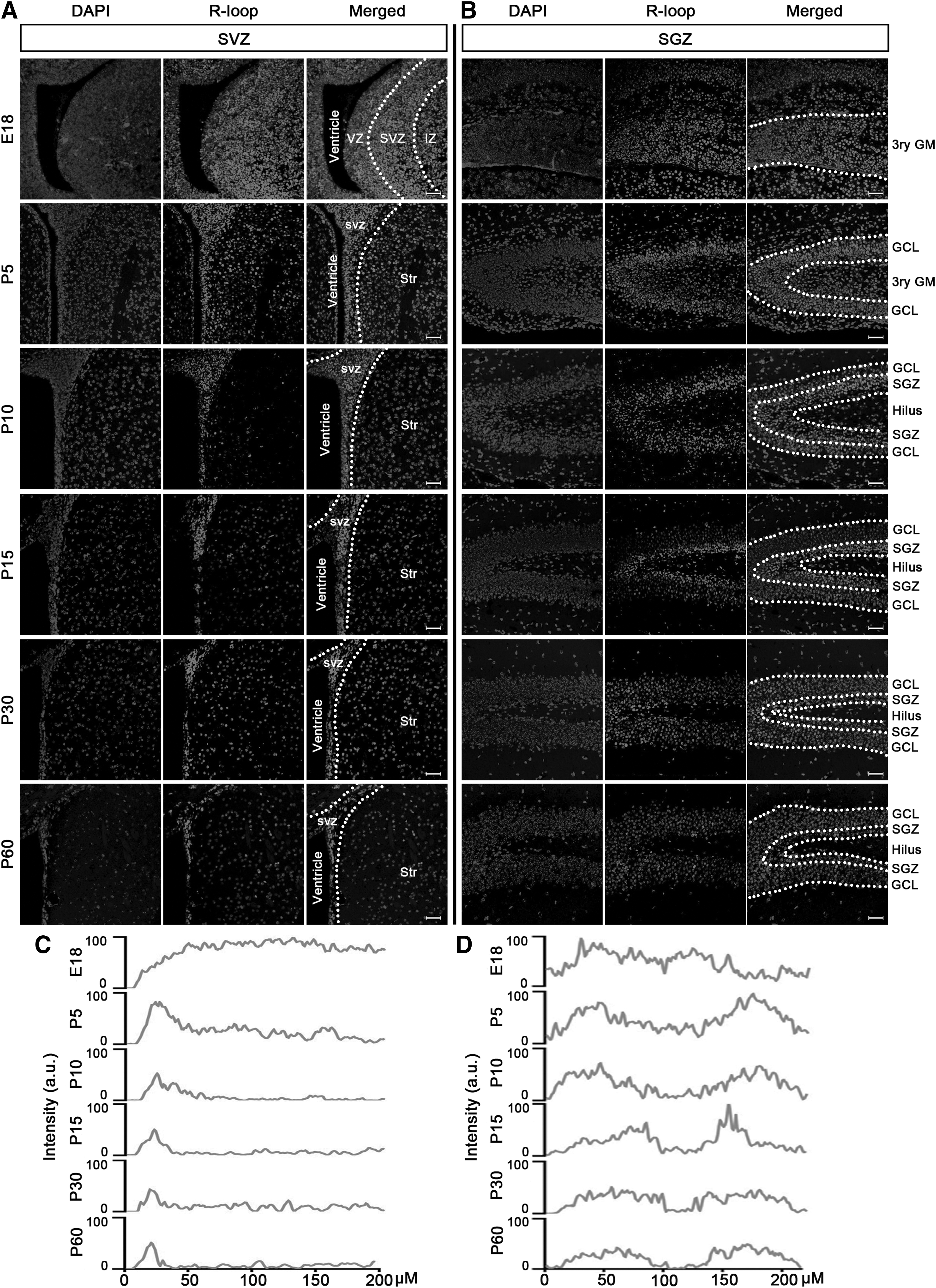
Dynamic changes in cells with high levels of R-loops in neurodevelopment. R-loop-positive cells were detected by immunofluorescence staining from E18 to P60 in SVZ
Cells with high levels of R-loops costain with SOX2 during mouse neurodevelopment
To further confirm that NSPCs are characterized by high levels of R-loops, costaining with the NSPC marker, SOX2, was performed. The immunostaining results in the SVZ showed that cells positive for both R-loops and SOX2 were enriched in the SVZ (Fig. 2C, E, G, I, K), with the exception of E18 (Fig. 2A). Notably, analysis of fluorescence intensity revealed that SOX2-positive cells exhibited high levels of R-loops across all time points (Fig. 2B, D, F, H, J), particularly in adulthood (Fig. 2L).

R-loops colocated with SOX2 during neurodevelopment in SVZ. Immunofluorescence staining revealed a high level of costaining between R-loops and SOX2 at E18
Subsequently, we examined the expression and distribution of R-loops and SOX2 in the SGZ. Similar to SVZ staining results, SOX2-positive cells with high levels of R-loops were consistently observed in the DG area during neurodevelopment. While minor variations in the distribution of R-loop and SOX2-positive cells were observed in the DG area at E18 and P5 (Fig. 3A–D), the occurrence of double-positive cells significantly increased in the SGZ from P10 to the adult stage (Fig. 3E–L).
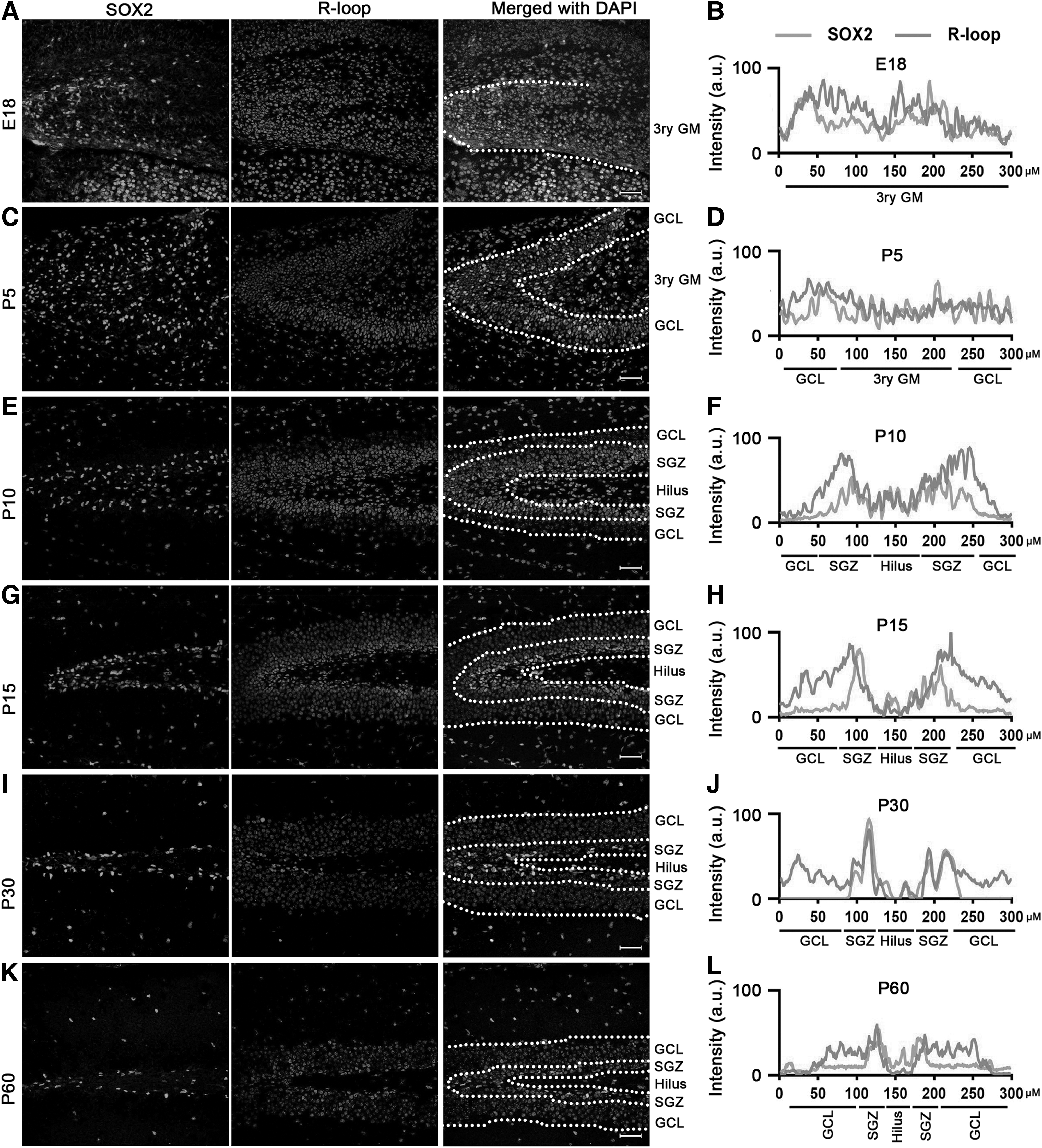
R-loops colocated with SOX2 during neurodevelopment in SGZ. Immunocytochemical double labeling revealed high levels of R-loops costained with SOX2 at E18
Based on the aforementioned results, we may conclude that NSPCs exhibit notably higher levels of R-loops compared with other cell types in the brain.
Cells with high levels of R-loops costain with H3K4me3 during mouse neurodevelopment
A careful inspection of the staining results demonstrates that cells exhibiting elevated R-loop levels were situated in the SVZ amid the ependymal cell layer and the striatal parenchyma, primarily at postnatal day 10 (P10). It is worth noting that our prior research indicated that H3K4me3, a marker associated with transcriptional activation, exhibited its strongest expression in the same anatomical region. 15 To validate the colocalization of R-loops and H3K4me3, we conducted double immunofluorescence staining.
As anticipated, at various stages of neurodevelopment, the vast majority of cells with heightened R-loop levels also exhibited costaining for H3K4me3 in both SVZ (Fig. 4A) and SGZ (Fig. 5A). Additionally, analysis of fluorescence intensity revealed a high degree of similarity between the fluorescence kurtosis plot of R-loops and that of H3K4me3, especially in the DG area (Figs. 4B and 5B). The similarity between the two fluorescence patterns was even more pronounced than that observed for SOX2.
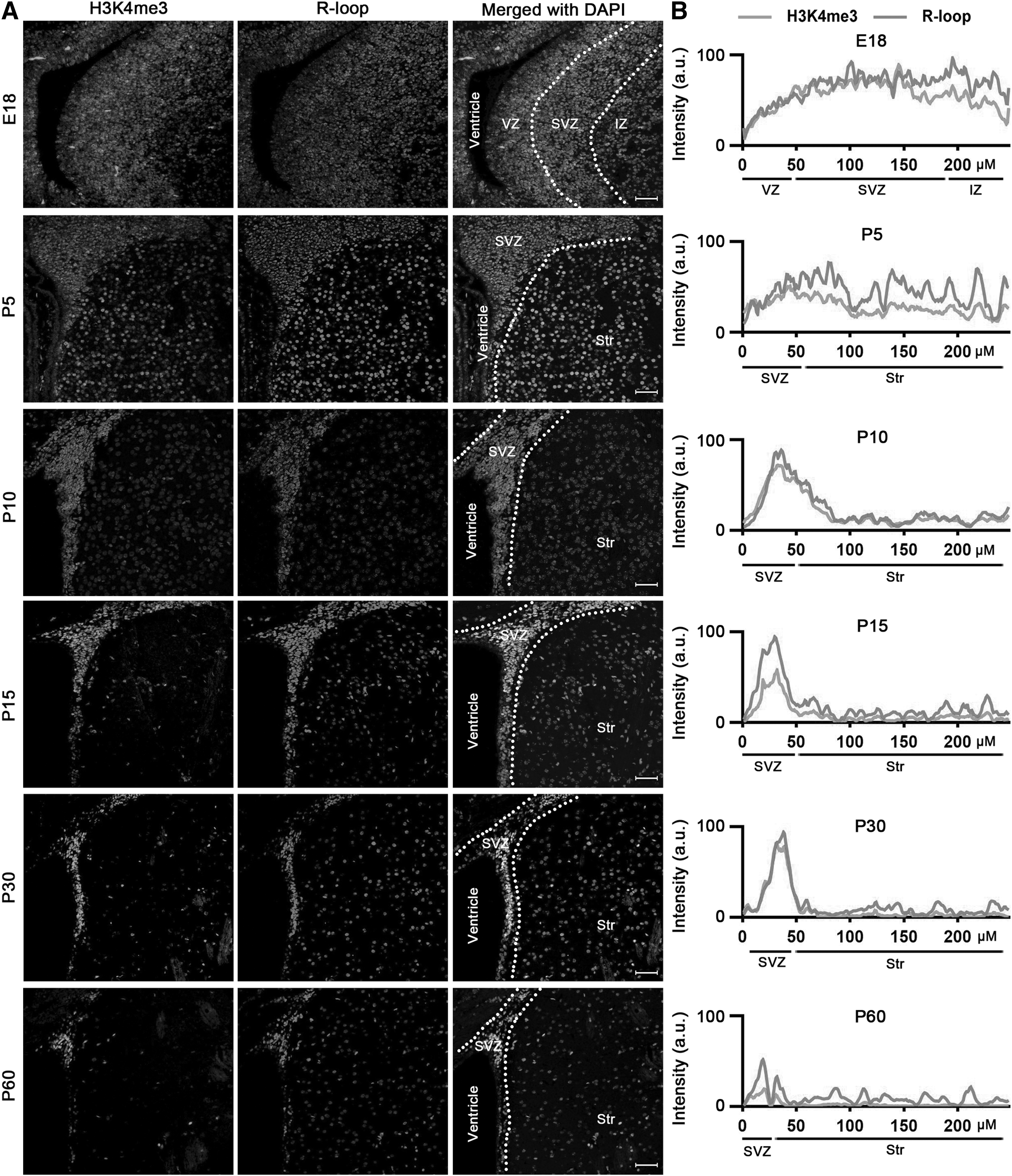
R-loops colocated with H3K4me3 during neurodevelopment in SVZ.
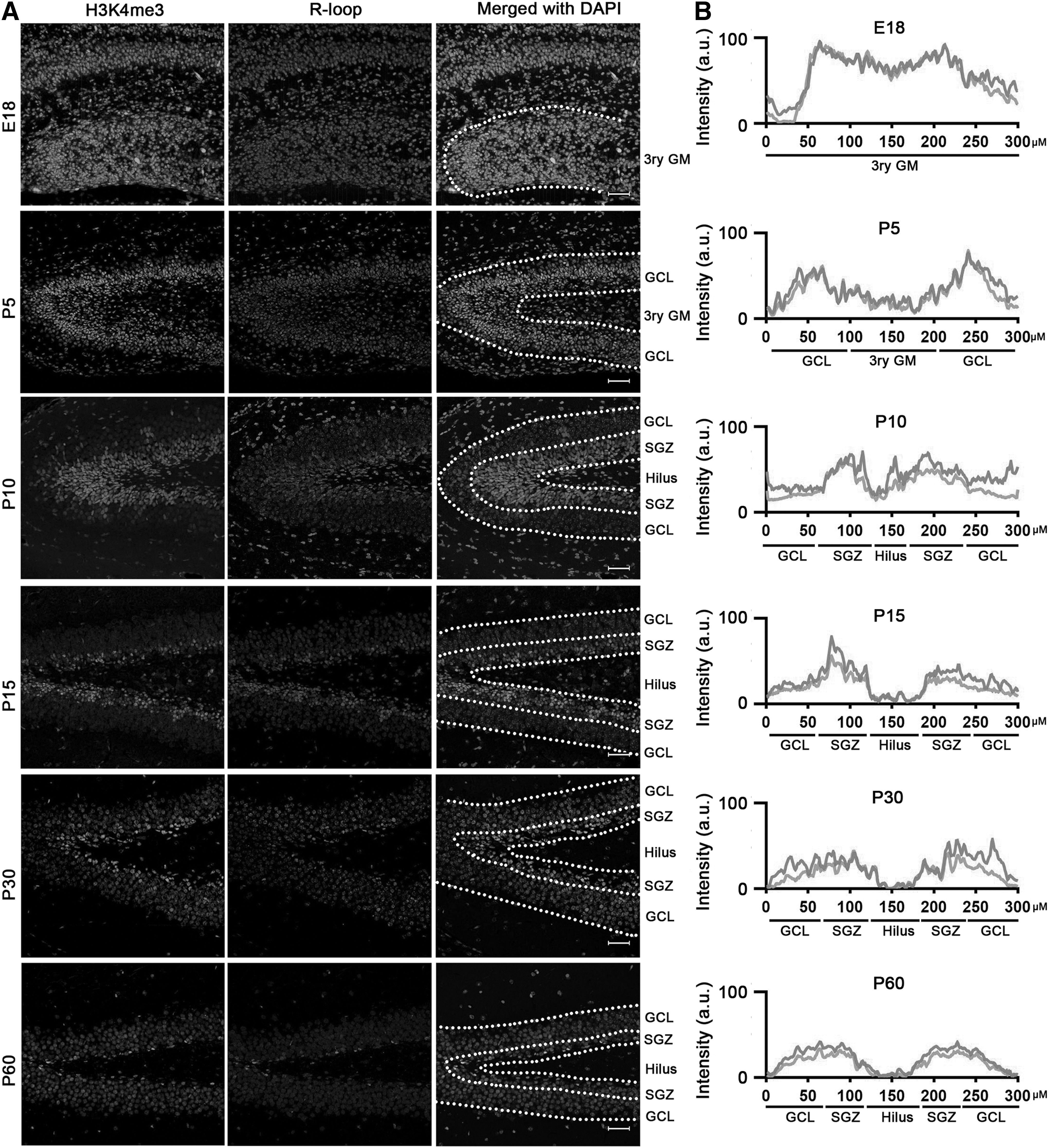
R-loops colocated with H3K4me3 during neurodevelopment in SGZ.
Our findings, in conjunction with prior research on the effects of R-loops, 16 confirm that the high level of R-loops is associated with regulation of gene expression and potentially serves as a marker for transcriptional activation in mouse NSPCs.
Cells with high levels of R-loops costain with transcriptional activation markers in both developing and mature brain tissues
To further substantiate the association between R-loops and transcriptional activation in the brain, we conducted double-label immunofluorescence staining to investigate the colocalization of R-loops with Ki-67 (a cell proliferation marker), serine 5 (ser5, an indicator of mRNA elongation), and H3K27ac (a marker of transcriptional activation). Immunocytochemical double labeling unveils Ki-67-positive cells showing high levels of R-loops in both SVZ and SGZ at P10 and P60 (Figs. 6A–D and 7A–D).
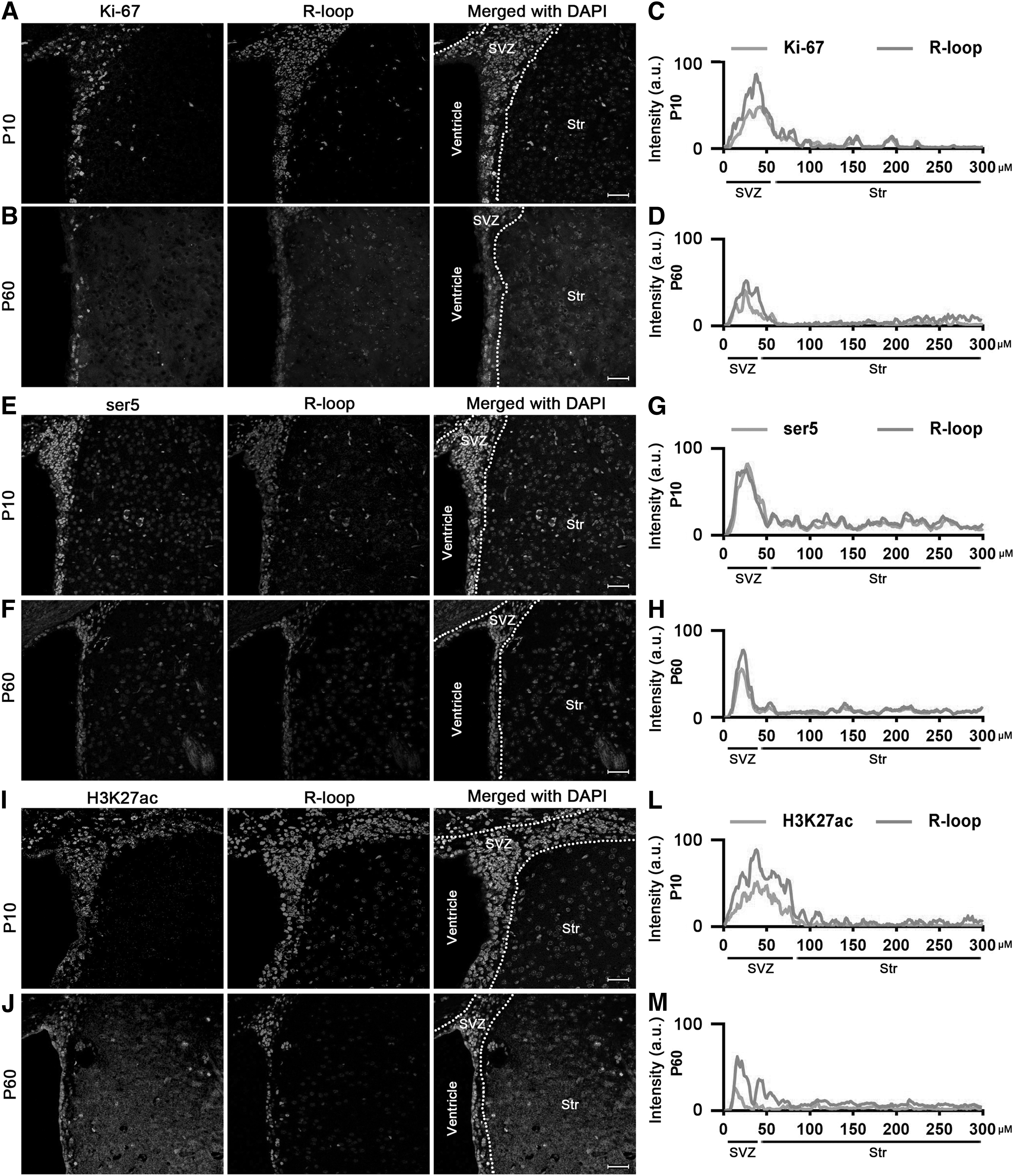
Ki-67, ser5, and H3K27ac costained with R-loops during neurodevelopment in SVZ at P10 and P60. Immunocytochemical double labeling revealed R-loops costained with Ki-67
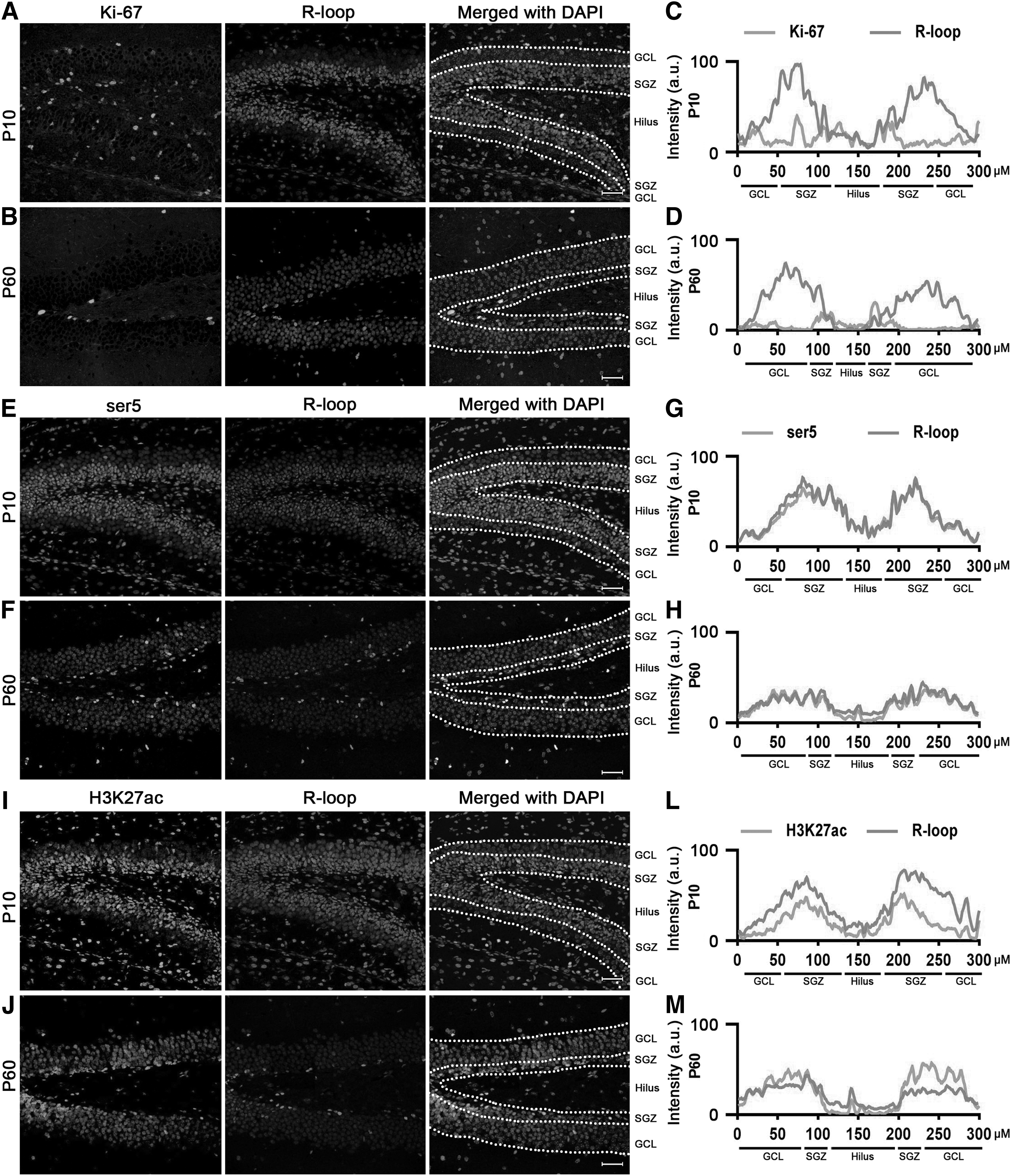
Ki-67, ser5, and H3K27ac costained with R-loops during neurodevelopment in SGZ at P10 and P60. Immunocytochemical double labeling showed high levels of R-loops in Ki-67
Moreover, consistent with H3K4me3 results, cells characterized by high levels of R-loops colocalized with ser5 (Fig. 6E, F) and H3K27ac (Fig. 6I, J) in the SVZ area at both P10 and P60. Notably, the fluorescence intensity analysis indicated that the fluorescence kurtosis plot of these transcriptional activation markers showed a strong overlap with R-loops (Fig. 6G, H, L, M).
In SGZ, we also observed colocalization between R-loops and ser5 (Fig. 7E–H) as well as H3K27ac (Fig. 7I–M). These findings collectively furnish robust evidence supporting a strong correlation between high levels of R-loops and transcription-activated cells during mouse neurodevelopment.
Discussion
Control of cell proliferation and directed differentiation are essential for the effectiveness of NSPC treatment in CNS diseases. While numerous studies have underscored the significance of epigenetic regulation in NSPC proliferation and differentiation, several aspects remain incompletely understood. 17,18 A novel perspective is to investigate the regulation of gene expression by R-loops.
Unlike typical epigenetic signatures, R-loops are three-stranded nucleic acid structures strongly associated with the regulation of gene expression, particularly during mammalian ontogeny. 19,20 During differentiation of embryonic stem cells (ESCs), distinct cell-type-specific R-loops are present in each lineage, with corresponding cotranscriptional formation at cell-type-specific genes.
Specifically, after differentiation of ESCs into NSPCs, R-loops mainly exist within neural specification genes such as PAX6, DLL1, and POU3F2. These R-loops exhibit a positive correlation with an increase in the active chromatin state and decrease in repressive chromatin marks. 21 These phenomena suggest that gene expression is tightly linked to changes in the levels of R-loops.
In this study, we observed that the levels of R-loop dynamics changed during neurodevelopment. Specifically, cells with high levels of R-loops are primarily localized within the neurogenesis area, concomitant with a reduction in the population of cells exhibiting heightened R-loop levels throughout neurodevelopment. Furthermore, these cells with high R-loop levels colocalize with SOX2 and exhibit a high degree of overlap with transcriptional activation markers, including H3K4me3, H3K27ac, and ser5.
Our finding suggests that R-loop may play a critical role in regulation of NSPC proliferation and differentiation. This study may offer a novel perspective on a therapeutic strategy for CNS diseases and provide a potential epigenetic feature for screening and identifying key therapeutic genes for such conditions.
SOX2, a transcription factor, plays a pivotal role in regulating development and self-renewal of NSPCs, influencing the maintenance of stemness properties in these cells. 22,23 A high SOX2 expression level is linked with preservation of NSPC stemness, promoting self-renewal and proliferation, while inhibiting NSPC differentiation into specific cell types. 24
Conversely, a decrease in SOX2 expression triggers the loss of stemness in NSPCs, inciting the differentiation of NSPCs into mature neurons or glial cells and limiting self-renewal capacity. 25 In this study, SVZ staining results revealed that cells with a high expression level of SOX2 were predominantly situated within the ependymal cell layer. Although cells with high R-loop levels costained with SOX2, most of them did not have high expression of SOX2.
Previous research has identified four major types of NSPCs within the SVZ niche, including type E cells (ependyma-like stem NSPCs), type B cells (quiescent or dormant NSPCs), type C cells (transit-amplifying progenitors), and type A cells (migrating neuronal precursors). 26,27 Type B cells are located between type A cells and the underlying striatal parenchyma, as well as between type A cells and the ependymal cells, whereas type C cells are positioned around type A cells. 28
Our findings indicated that cells with high levels of R-loops were primarily present between the ependymal cell layer and the striatal parenchyma. In addition, the fluorescence intensity analysis showed a significant resemblance between the fluorescence kurtosis plots of R-loops and H3K4me3 (a specific type C/A cell marker). Therefore, it is reasonable to speculate that type C/A cells are represented by high levels of R-loops. However, further research is required to validate our hypothesis.
The staining results reveal a notable concentration of cells with heightened R-loop levels within the neurogenesis area at time points ranging from P10 to the adult stage. Conversely, cells positive for R-loops are widely dispersed throughout the brain, showing consistent levels from E18 to P5. This discrepancy may arise from distinctions between embryonic and adult NSPCs in terms of their abundance and differentiation potential.
Embryonic NSPCs undergo asymmetric division, giving rise to neurons directly or through intermediate progenitor cells, as well as oligodendrocytes. For example, embryonic NSPCs experience apical detachment and subsequently differentiate into astrocytes. In contrast, adult NSPCs maintain the capacity to generate neurons and oligodendrocytes, but lack the propensity to differentiate into astrocytes. 29
Moreover, embryonic NSPCs exhibit robust proliferation and self-renewal capacity, enabling significant expansion, whereas adult NSPCs demonstrate limited proliferation and self-renewal ability, resulting in a slower turnover rate and lower cell numbers compared with their embryonic counterparts. 30 Our finding suggested that R-loops might play a more critical role in regulation of NSPC proliferation and differentiation than during embryonic phases.
Conclusions
The R-loop, a recently identified, novel, epigenetic regulatory mechanism, is highly dynamic and constantly reshaped during neurodevelopment. 31 It can exert influence on DNA methylation and histone modifications, which are fundamental mechanisms of epigenetic regulation. 32,33 On the one hand, methylation of CpG sites near R-loop-forming regions stabilizes the R-loop structure, while demethylation decreases stability and promotes proper DNA processing 34 ; and on the other hand, R-loops can recruit histone-modifying enzymes to specific genomic regions, leading to changes in histone acetylation or methylation. 35,36
Our study suggested that the levels of R-loops decreased during development of NSPCs and costained with transcriptional activation markers. The findings of this study provide novel perspectives on initiation of neurodevelopment and present an epigenetic signature that may be utilized for identification and characterization of pivotal regulatory genes involved in neurogenesis.
Nonetheless, additional research, particularly whole epigenome analysis and R-loop profiling, is required to gain a more comprehensive understanding of the function of R-loops in neurodevelopment.
Footnotes
Acknowledgment
The authors thank the Laboratory Animal Center of Xi'an Jiaotong University for mouse husbandry.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the National Natural Science Foundation of China (No. 82271200); Fundamental Research Funds for the Central Universities (xzy012022035); Key Research and Development Projects of Shaanxi Province (2023-YBSF-303); and Natural Science Basic Research Program of Shaanxi Province (S2022-JC-QN-1370).
Supplementary Material
Supplementary Table S1