
Abstract
Direct conversion is an innovative new technology that involves the conversion of somatic cells to target cells without passing through a pluripotent state. Forced expression alone or in combination with transcription factors (TFs), which are critical for the generation of target cells, is important for successful direct conversion. However, most somatic cells are unable to directly convert into target cells even with forced expression. We herein demonstrated that epithelial–mesenchymal transition (EMT) is advantageous for the direct conversion of somatic cells. We previously reported that mouse keratinocytes converted into neural crest cells (NCCs) following the forced expression of the NCC specifier Sox10 in combination with expression of the TFs Snail1, Slug, Twist1, and Tcfap2a (4 TFs). 4 TFs induced EMT in keratinocytes; therefore, EMT was considered to be advantageous for direct conversion. The direct conversion of mouse mammary gland epithelial cells (NMuMG cells) into NCCs was not observed with the forced expression of Sox10, but was detected with the expression of Sox10 following the induction of EMT by 4 TFs. Furthermore, TGF-β1-induced EMT and Sox10 expression directly converted NMuMG cells into NCCs. These results suggest that the induction of EMT in somatic cells is advantageous for direct conversion.
Introduction
The introduction of genes was recently shown to induce a shift in the developmental fate of cells without passing through stem cell intermediate states. 1 This innovative technology, direct conversion, has been utilized to convert somatic cells into other types of somatic cells. The induction of a combination of cell type-specific genes, mainly transcription factors (TFs), directly converts some cell types, typically fibroblasts, into other types. The genes induced are fundamentally selected from TFs that constitute the gene regulatory network in the development of “target” differentiated cells. For example, mouse embryonic fibroblasts (MEFs) were directly converted into hepatocytes after the induction of the following TFs for hepatocyte development: Hnf4a, Foxa1, Foxa2, Foxa3, Gata6, and Cdx2, 2,3 or into cardiomyocytes by the forced expression of critical TFs for cardiomyocyte generation: Gata4, Mef2c, and Tbx5. 4 We previously demonstrated that MEFs were directly converted into neural crest cells (NCCs) with the forced expression of the NCC specifier TF, Sox10. 5
Fibroblasts, a typical mesenchymal cell, are mainly utilized in direct conversion because they are readily available and easy to culture. 6 Furthermore, fibroblasts may be converted into cells in different germ layers other than the mesoderm. In the aforementioned direct conversion, mesoderm fibroblasts are directly converted into endoderm hepatocytes, ectoderm cardiomyocytes, and NCCs. However, few cells besides fibroblasts are directly converted into other cell types, except for descendants derived from the same progenitor (e.g., glial cells are converted into neuronal cells), or into the same cell type (e.g., pancreatic exocrine cells are converted into pancreatic β cells). Moreover, cells other than fibroblasts are rarely converted to completely different cells in different germ layers. We previously showed that keratinocytes (XB2 cells) were not converted into NCCs with the forced expression of Sox10, by which MEFs are converted into NCCs. 7 However, with the application of direct conversion technology to regenerative medicine, target cells will not always be fibroblasts. Therefore, the direct conversion of various cell types will become necessary for the application of this technology to medicine.
The present study aimed to directly convert various somatic cell types. We previously demonstrated that the direct conversion of XB2 keratinocytes into NCCs was not observed with the overexpression of Sox10 alone, but was detected with the overexpression of Sox10 in combination with the expression of Snail1, Slug, Twist1, and Tcfap2a. 7 Since Snail1, Slug, Twist1, and Tcfap2a function in the induction of epithelial–mesenchymal transition (EMT), 8,9 EMT may be advantageous for the direct conversion of cells. Therefore, we herein performed the direct conversion of mouse mammary gland epithelial cells (NMuMG cells) into NCCs with EMT.
Materials and Methods
Cell culture
NMuMG cells (purchased from Public Health England) were cultured in DMEM high glucose medium (Wako Pure Chemical Co., Osaka, Japan) supplemented with 10% fetal calf serum (FCS) and 10 µg/mL insulin (Wako Pure Chemical Co.) at 37°C and 5% CO2. To induce TGF-β1, recombinant human TGF-β1 (PeproTech Cranbury, NJ, USA) was used at 5 ng/mL.
Retroviral transfection and NCC generation
The retrovirus vectors pMXs-GW and pMYs-GW for Snail1, Slug, Twist1, Tcfap2a, and Sox10 were a gift from the human proteome expression resource (HuPEX) library (HuPEX clones in Human Gene and Protein Database (HGPD), http://www.HGPD.jp/, the National Institute of Advanced Industrial Science and Technology, Japan). Retrovirus preparation and infection were described in detail in our previous study. 7 A total of 1.0 × 104 cells/cm2 of NMuMG cells were infected with the retrovirus mixture for Snail1, Slug, Twist1, and Tcfap2a. Twenty-four hours later, cells were maintained under NCC conversion conditions: α-MEM (GIBCO, NY, USA) supplemented with 10% FCS, 10−7 M dexamethasone (Sigma-Aldrich, St. Louis, MO, USA), 20 pM bFGF (R&D Systems Inc., Minneapolis, MN), 10 pM cholera toxin (Sigma-Aldrich), and 100 ng/mL human recombinant endothelin-3 (EDN3; Peptide Institute, Inc., Osaka, Japan) at 37°C and 5% CO2. After 3 days, NMuMG cells were infected with the retrovirus for Sox10.
Flow cytometry and cell sorting
Cell preparation for the analysis and isolation of P75+ cells was described in detail in our previous study. 7 Anti-P75 antibodies (ab8875; Abcam, Cambridge, UK) and DyLight 649-conjugated anti-Rabbit IgG (BioLegend, San Diego, CA, USA) were used to stain infected cells. Cell sorting and analyses were performed using a FACS Aria flow cytometer (Becton Dickinson, Franklin Lakes, NJ, USA).
In the analysis of E-cadherin, dissociated cells were fixed in 4% paraformaldehyde (PFA) in phosphate-buffered saline (PBS) for 8 min and made permeable with 0.1% Triton X-100 (Sigma-Aldrich) in 5% bovine serum albumin (BSA) PBS for 15 min. Cells were then stained with Alexa Fluor 647-conjugated anti-mouse E-cadherin (BioLegend, San Diego, CA, USA). E-cadherin expression was assessed by measuring the total mean fluorescence intensity of cells using FACS Diva (Becton Dickinson).
Maintenance and differentiation of P75+ cells
Isolated P75+ cells were cultured under hypoxic conditions with 5% O2 and 5% CO2 at 37°C under the NCC culture conditions described by Morrison et al. 10 ; a 5:3 mixture of DMEM-low: neurobasal medium (GIBCO) supplemented with 15% Chick Embryo Extract (CEE, US Biological, Salem, MA, USA), 1% N2 (GIBCO), 2% B27 (GIBCO), 50 µM 2-mercaptoethanol (Sigma-Aldrich), 35 ng/mL all-trans retinoic acid (Sigma-Aldrich), 20 ng/mL IGF-1 (R&D systems), 100 ng/mL EDN3 (Peptide Institute, Inc.), and 20 ng/mL bFGF (R&D Systems). After 6 days, P75+ cells were differentiated into NC derivatives using NCC differentiation medium. NCC differentiation medium had the same components as the medium used for NCC culture conditions, except that it contained 1% CEE and 10 ng/mL bFGF. 10 Regarding specific neural cell differentiation or glial cell differentiation, we incubated P75+ cells in NCC differentiation medium supplemented with 50 ng/mL BMP-2 (R&D Systems) or 1 nM forskolin (Sigma-Aldrich) and 1 nM Nrg-1 (R&D Systems), respectively. All differentiation cultures were performed under hypoxic conditions at 37°C.
Immunohistochemical analysis
Colony fixation, permeabilization, and blocking were performed as previously. 11 The primary antibodies were as follows: mouse anti-mouse neuronal class III β-tubulin (1:500; TuJ-1; COVANCE, Princeton, NJ, USA), rabbit anti-mouse glial fibrillary acidic protein (GFAP, 1:500; Z0334; Dako Cytomation, Denmark), and anti-mouse α-smooth muscle actin (SMA, 1:500; 1A4; Sigma-Aldrich). The secondary antibodies were as follows: Texas Red-conjugated anti-mouse IgG (1:500; Molecular Probes, Life Technology) and Alexa Fluor 488-conjugated anti-rabbit IgG (1:500; Molecular Probes, Life Technology). Nuclei were stained with Hoechst 33258 (Sigma-Aldrich).
RT-PCR and quantitative PCR analysis
Total RNA was purified using Isogen (Nippon Gene, Tokyo, Japan), and first-strand cDNA synthesis was conducted with Superscript III (Invitrogen, Carlsbad, CA, USA). Quantitative PCR (q-PCR) was performed with the Thermal Cycler Dice Real Time System (TAKARA, Ohtsu, Japan) and SYBR premix Ex taq (TAKARA). Data were analyzed by the ΔΔCt method. Experiments were performed in triplicate, and expression levels relative to β-actin are shown. Reverse transcriptase-PCR (RT-PCR) and q-PCR primers are described in Supplementary Data S1.
Results
Four TFs generate EMT in XB2 keratinocytes
We previously reported that mouse keratinocytes XB2 were converted into NCCs by the forced expression of Sox10 in combination with the expression of Snail1, Slug, Twist1, and Tcfap2a (4 TFs). 8,9 Therefore, we hypothesized that 4 TFs induced EMT in keratinocytes, which was advantageous for their direct conversion. We noted a marked change from the tightly associated morphology of XB2 cells to a spindle-shaped migratory morphology, which is typical of mesenchymal cells after the forced expression of 4 TFs (Fig. 1A). 12 Flow cytometry showed the downregulated expression of E-cadherin at 3 days after the forced expression of 4 TFs (Fig. 1B). To investigate whether 4 TFs induced EMT in XB2 cells, we compared the expression of the epithelial marker E-cadherin and keratinocyte marker Keratin 5 in XB2 cells before and after the expression of 4 TFs using q-PCR. Three days after the expression of 4 TFs, E-cadherin and Keratin 5 expression levels were lower than those in intact XB2 cells (Fig. 1C). Therefore, the forced expression of 4 TFs induced EMT in keratinocytes.
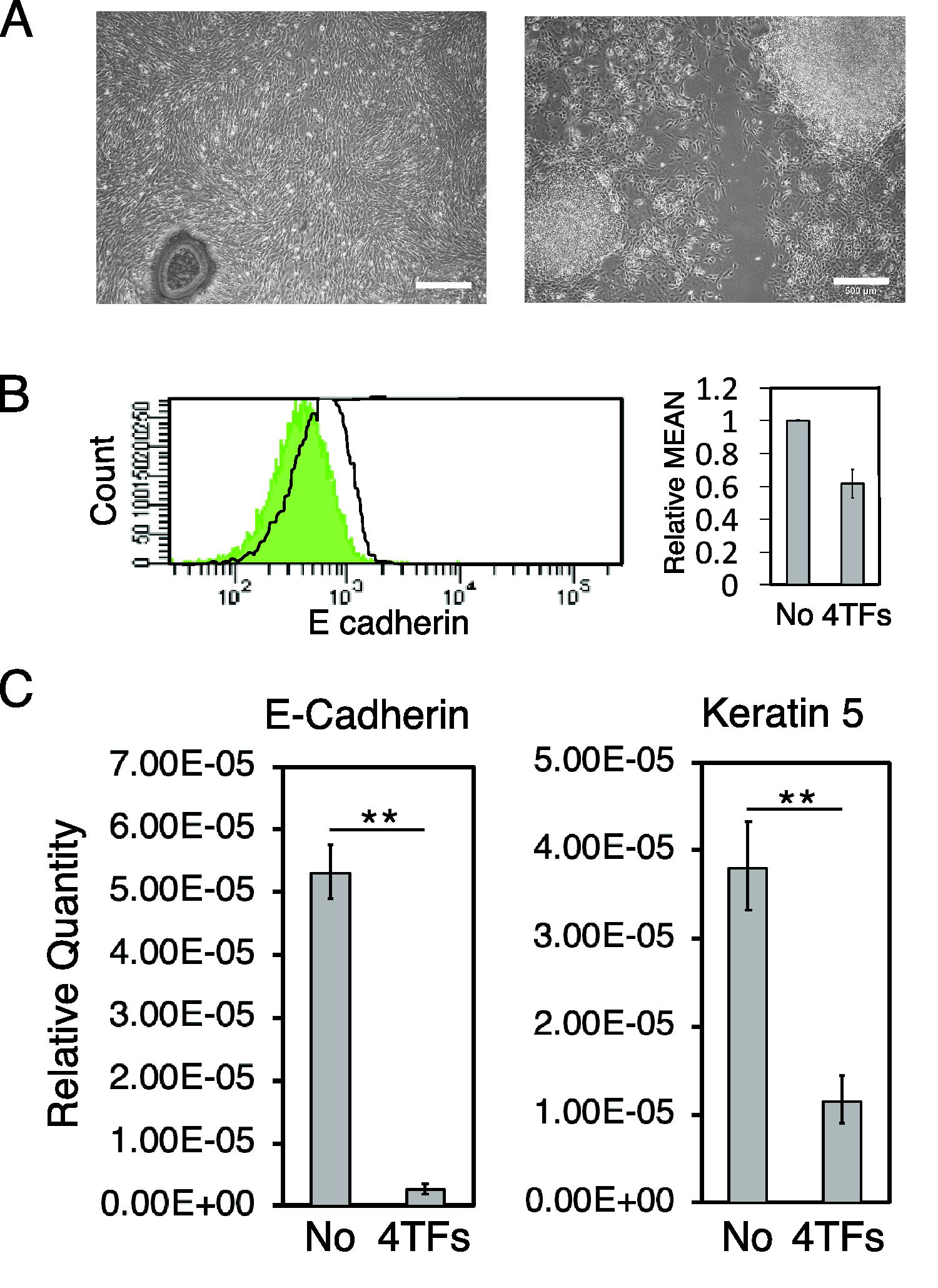
4 TFs induce EMT in keratinocytes.
4 TFs generate EMT in mammary gland epithelial cells
We confirmed that NMuMG cells were not directly converted into P75+ NCCs after a 10-day culture with the forced expression of Sox10 (Fig. 2A). Therefore, we investigated whether the forced expression of 4 TFs induced EMT in NMuMG cells in the same manner as in XB2 keratinocytes. After the forced expression of 4 TFs, NMuMG cells changed their shape into a spindle-like migratory morphology (Fig. 2B). Flow cytometry showed the downregulated expression of E-cadherin at least 2 days after the forced expression of 4 TFs, which peaked after 3 days and remained unchanged thereafter (Fig. 2C, Supplementary Fig. S1A). To investigate whether EMT was induced in NMuMG cells by 4 TFs, we compared the expression of E-cadherin and the mesenchymal cell marker Vimentin in NMuMG cells before and after the expression of 4 TFs using q-PCR. Three days after the expression of 4 TFs, the expression level of Vimentin was slightly higher than in intact NMuMG cells (Fig. 2D, Supplementary Fig. S1B), whereas that of E-cadherin was lower (Fig. 2D, Supplementary Fig. S1B). Therefore, the forced expression of 4 TFs appeared to induce EMT in NMuMG cells.
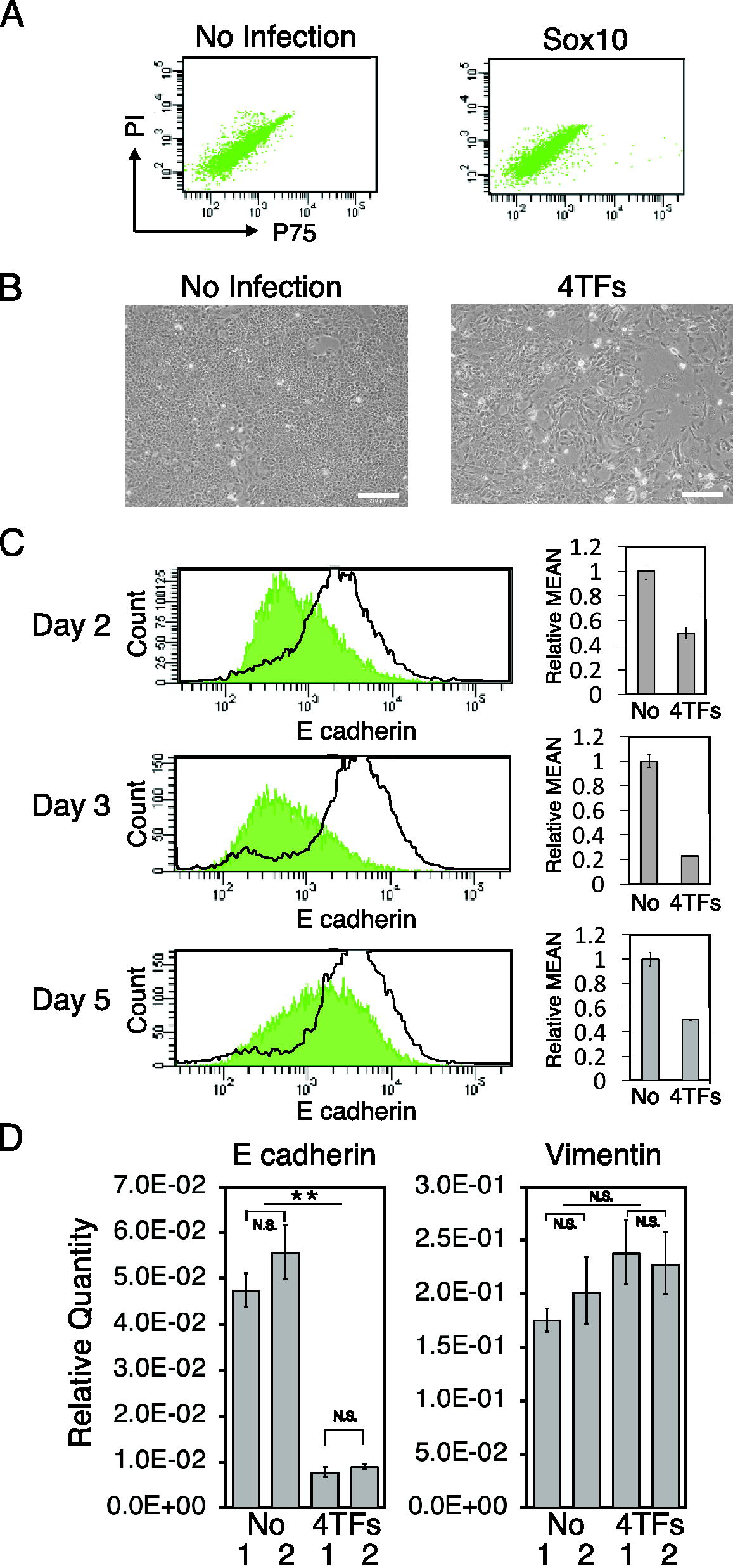
4 TFs induce EMT in mammary gland epithelial cells.
Sox10 in combination with EMT by 4 TFs converts NMuMG cells into NCCs
We investigated whether the forced expression of Sox10 in combination with EMT by 4 TFs converted NMuMG cells into NCCs in the same manner as XB2 keratinocytes. Three days after the forced expression of 4 TFs, NMuMG cells overexpressed Sox10 and were cultured under NCC conversion conditions (Fig. 3A). Flow cytometry showed that NMuMG cells generated P75+ cells on day 10 of the culture after the forced expression of Sox10 (Fig. 3B). RT-PCR revealed that the P75+ cells generated expressed the following NCC marker genes: Sox10, Foxd3, P75, Pax3, and Id2 (Fig. 3C). We isolated P75+ and P75− cells and cultured them under the NCC culture conditions previously established by Morrison et al. 10 After 6 days, P75+ cells were maintained, proliferated, and generated colonies under these culture conditions. Colonies were large colonies from which many cells migrated, suggesting the strong proliferation and migratory potencies of P75+ cells (Fig. 3D). On the other hand, P75− cells were not or were only slightly maintained and formed a few very small clusters in which cells tightly adhered to each other under NCC culture conditions (Fig. 3D). To investigate the differentiation potencies of the P75+ cells generated, the medium after 6 days of cultivation under NCC culture conditions was changed to NCC differentiation medium (Fig. 3A). After 21 days of culture in NCC differentiation medium, colonies generated from P75+ cells contained TuJ-1+ neuronal cells (TuJ1+ N cells) and GFAP+ glial cells (GFAP+ G cells) (Fig. 3E–G, Supplementary Fig. S2A, B). Some colonies also contained αSMA+ smooth muscle cells (αSMA+ SM cells) (Fig. 3H). On the other hand, the cluster formed from P75− cells did not generate TuJ-1+ N cells, GFAP+ G cells, or αSMA+ SM cells (data not shown). RT-PCR showed that the neuronal cell marker genes TuJ-1, Mash-1, and Peripherin and glial cell marker genes Gfap, S100β, and B-fabp were expressed in P75+ cells cultured in a differentiation medium (Fig. 3I). These results suggest that Sox10 overexpression after the induction of EMT by 4 TFs converted NMuMG cells into multipotent NCCs.

Sox10 in combination with 4 TFs converted mammary gland epithelial cells into P75+ NCCs.
TGF-β1-induced EMT is advantageous for the direct conversion of NMuMG cells into NCCs
NMuMG cells have been shown to undergo EMT in response to TGF-β1. 13 We investigated whether TGF-β1-induced EMT also directly converted NMuMG cells into NCCs with Sox10 expression. After the TGF-β1 treatment, NMuMG cells changed their shape into a spindle-like migratory morphology (Fig. 4A). Flow cytometry showed the downregulated expression of E-cadherin after the TGF-β1 treatment (Fig. 4B, Supplementary Fig. S3A), which peaked after 3 days (Fig. 4B, Supplementary Fig. S3A). In comparisons with intact NMuMG cells, q-PCR revealed the downregulated expression of E-cadherin, the upregulated expression of N-cadherin, and no change in the expression of Vimentin (Fig. 4C, Supplementary Fig. S3B), showing the induction of EMT in TGF-β1-treated NMuMG cells.

TGF-β1 induced EMT in mammary gland epithelial cells.
Three days after the TGF-β1 treatment, NMuMG cells overexpressed Sox10 and were cultured under NCC conversion conditions (Fig. 5A). Flow cytometry showed that NMuMG cells generated P75+ cells on day 10 of the culture after Sox10 overexpression (Fig. 5B). We isolated P75+ and P75− cells and cultured them under NCC culture conditions. P75+ cells proliferated and generated colonies under these conditions (data not shown). P75− cells were not maintained or only formed a few very small clusters under NCC culture conditions (data not shown). The colonies generated from P75+ cells expressed the NCC marker genes Sox10, Foxd3, P75, Pax3, and Id2 (Fig. 5C). In the differentiation culture, GFAP+ G cells and TuJ-1+ N cells were generated in the colonies formed from P75+ cells after 21 days of culture (Fig. 5D). The cluster formed from P75− cells did not generate GFAP+ G cells or TuJ-1+ N cells (data not shown). RT-PCR showed that the N cell marker genes Mash1, TuJ-1, Nestin, and Peripherin and the G cell marker genes Gfap, S100β, and B-fabp were expressed in P75+ cells cultured in differentiation medium (Fig. 5E). These results demonstrated that Sox10 overexpression after TGF-β1-induced EMT converted NMuMG cells into multipotent NCCs, suggesting that the induction of EMT was advantageous for the direct conversion of NMuMG cells.
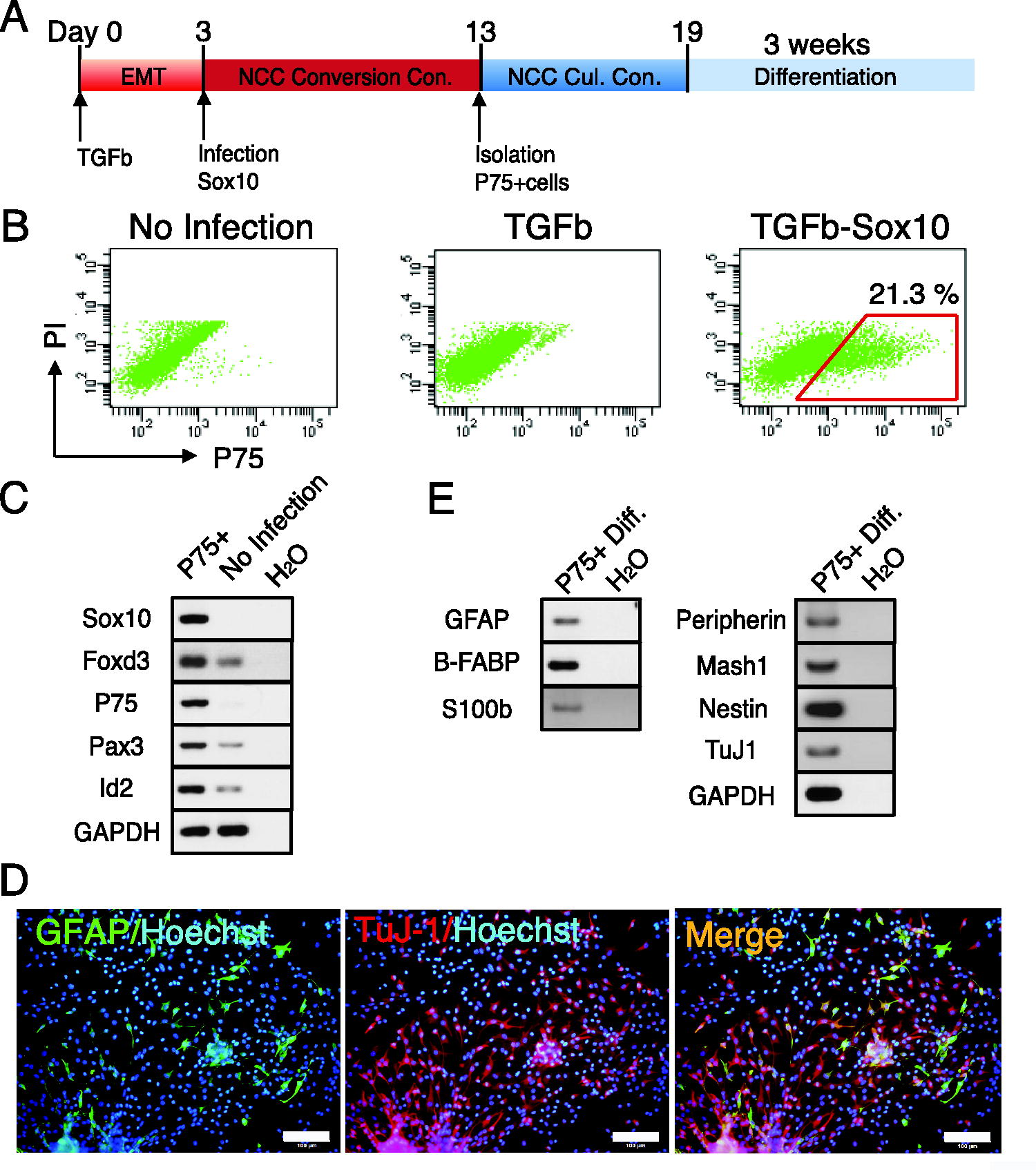
The forced expression of Sox10 after the TGF-β1 treatment converted mammary gland epithelial cells into multipotent P75+ NCCs.
Discussion
Direct conversion is an innovative new technology by which somatic cells shift into other types of somatic cells with the induction of genes without passing through stem cell intermediate states. Fibroblasts, a typical mesenchymal cell, are mainly utilized in direct conversion because they are readily available and easy to culture. On the other hand, other types of somatic cells are seldom used for direct conversion, and even when used, they rarely convert into completely different cells in different germ layers. We recently showed that MEFs and mouse bone marrow-derived ST2 mesenchymal stroma cells were directly converted into NCCs with the forced expression of Sox10, a master specifier for the development of NCCs; however, keratinocytes were not converted into NCCs in the same manner. 5,7 We also demonstrated that keratinocytes were directly converted into NCCs with the expression of Sox10 in combination with the expression of Snail1, Slug, Twist1, and Tcfap2a. 7 Snail1, Slug, Twist1, and Tcfap2a are TFs relevant to NCC specification, 9 whereas Snail, Slug, and Twist1 are major inducers of EMT, 8 and Tcfap2a is associated with EMT. 9 Based on the results obtained on the direct conversion of keratinocytes, we hypothesize that the induction of EMT is advantageous for direct conversion.
EMT has been reported in various embryonic development events, including gastrulation, mesoderm formation, and NCC development, and has been associated with physiological and pathological responses, wound healing, fibrosis, and cancer progression. 12,14 A series of TFs regulate the induction of EMT by repressing epithelial genes while activating mesenchymal genes. We confirmed that Snail1, Slug, Twist1, and Tcfap2a repressed the expression of E-cadherin and induced a spindle-shaped migratory morphology, which is typical of mesenchymal cells in XB2 keratinocytes and NMuMG cells, showing that 4 TFs induced EMT. After the induction of EMT, NMuMG cells were directly converted into NCCs by the forced expression of Sox10. Furthermore, NMuMG cells were directly converted into NCCs by the forced expression of Sox10 after TGF-β1-induced EMT. These results suggest that EMT is favorable for direct conversion.
An advantageous condition for direct conversion is considered to involve two states: one in which the master TFs for direct conversion easily bind to their target sites, and another in which cells themselves are highly susceptible to changes caused by direct conversion. 15 The former state may be induced by the epigenetic modulation of the genome in the “initial” somatic cells. EMT-TFs directly regulate chromatin-remodeling enzymes or induce them by physical interactions, and this interplay plays a pivotal role in EMT. 16 Twist and Snail family proteins interact with Polycomb Repressive Complex 1 (PRC1) and recruit it to the CDH1 promoter in order to induce EMT. 16 –20 Snail1 and Slug also interact with PRC2 through its SNAG domain to repress CHD1. 16,17,21 In addition to the gene suppressive effects of EMT-TFs, TGF-β-induced EMT in hepatocytes caused a global reduction in heterochromatin marks and induced an open chromatin structure. 16,22 Although the relationship between EMT and chromatin states induced by epigenetic changes warrants further study, EMT may generate open chromatin states that are favorable for accessing the master TFs for direct reprogramming.
Another state may be induced by the acquisition of cellular plasticity in the “initial” somatic cells. Through EMT, epithelial cells remodel their adhesive junctions and cytoskeleton to acquire highly migratory and invasive properties. The phenotypic changes induced by EMT are reversible in many cases, and this reversibility is responsible for cell plasticity. 16 The induction of EMT was recently reported to confer plasticity and stemness to cells. The induction of EMT by the forced expression of Snail1 or Twist1 converted mature human mammary epithelial cells into stem-like cells with mammosphere-forming potency, self-renewal activity, and the ability to differentiate into epithelial cells. 23 Twist, Zeb, and Six1 transform mammary epithelial cell traits and endow stem cell-like characteristics together with the induction of EMT. 24,25 Although the precise mechanisms underlying the acquisition of stemness through EMT remain unclear, the induction of EMT is considered to induce plasticity in the genetic state of differentiated cells. EMT does not strictly switch from an epithelial to mesenchymal phenotype, it is an intermediate state that temporarily retains epithelial and mesenchymal features (intermediate/partial EMT). 26 The biphasic properties generated by EMT induce plasticity and may contribute to stemness and facilitate direct conversion, as demonstrated herein.
Although the molecular mechanisms by which EMT facilitates direct cellular conversion remain unclear; further studies may provide novel insights into these mechanisms. Furthermore, the direct conversion of various cell types will contribute to the application of direct conversion technology to regenerative medicine.
Footnotes
Acknowledgments
The authors thank Dr. Naoki Goshima (Molecular Profiling Research Center for Drug Discovery, National Institute of Advanced Industrial Science and Technology) for providing the retrovirus vector, Dr. Howard Green (Department of Cell Biology, Harvard Medical School) for providing XB2 cells, and other members of our laboratory for their discussions and critical reading of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Gifu University Graduate School of Medicine Research Grant Program and by a grant from the program Grants-in-Aid for Scientific Research (C) from the Japan Society for Promotion for Science (22K06808).
Supplementary Material
Supplementary Data S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3