
Abstract
Fasting regimens have shown profound impact on pro-longevity and tissue regeneration in diverse species. Physiological events can induce a regenerative response in adult stem cells. However, little is known about signaling and activation of adult stem cells which are modulated by fasting. This study analyzed the presence of hematopoietic stem/progenitor cells (HSPCs) and their circulation in the peripheral blood (PB) of healthy male adults practicing Ramadan fasting. Ten healthy male volunteers were enrolled in this prospective observational study. PB samples were collected twice daily on days 0, 10, 20, and 30 of Ramadan fasting (RF). Populations of stem cells and serum soluble factors were analyzed by flow cytometry. As a response to RF, we report an increase in the average absolute count of circulating of HSPCs, defined as LIN−CD45− and LIN−CD45+ cell subsets expressing the stem markers, CD34 and CD133. Changes in the number of HSPCs subsets reflected changes in the peripheral concentration of chemoattractant soluble factors during fasting. A chemotaxis assay showed a migratory property of HSPCs towards plasma, collected at D30 of fasting that contained a higher concentration of SCF and G-CSF. The relationship between RF and an increase in the number of circulating HSPCs in part, describes a regenerative response to the physiological changes during fasting and may open opportunities to define the role of dietary intervention in the stem cell therapy.
Introduction
The existence of a physiological process by which hematopoietic stem/progenitor cells (HSPCs) egress out of the bone marrow (BM) into the peripheral circulation has come a long way since its discovery over four decades ago and the term “mobilization” was coined. 1,2 HSPCs, known as precursors to all immune, blood, and tissue cell types continue to migrate, during steady-state homeostasis, via the bloodstream throughout adulthood at a very low frequency representing ∼0.01%–0.0001% of the mononuclear fraction in peripheral blood (PB). 3,4 It has been speculated that the migratory potential is essential for the continuous regenerative program of the BM niche itself, and potentially of other tissues. 5 Moreover, these cells have been described as “stem cells on patrol,” due to their ability to survey peripheral organs and stimulate the local production of tissue-resident innate immune and progenitor cells under both steady-state conditions and in response to inflammatory or regeneration signals. 6 Cellular and molecular mechanisms underlying the circulation of HSPCs and their physiological significance have remained relatively obscure but it is very likely that the process of mobilization makes use of physiological molecular pathways driving HSPCs into circulation. Studies by Borkowska and colleagues showed a tight process of circadian rhythm-regulated mobilization of primitive stem cells from BM into PB. 7,8 The Circadian oscillations of circulating HSPCs, especially CD34+ cells, are reported to be significantly higher in the evening than morning. 9 In addition to their physiological migration, studies showed an increased mobilization of these cells from their BM niches and a robust increase in their number in peripheral circulation in response to a variety of systemic “stressors.” 10 –14 Although their contribution to tissue repair and regeneration is uncertain, programmed increases of HSPCs due to a combination of paracrine effects, cytokines release, and direct cellular differentiation have been shown. Moreover, mobilization of HSPCs has been reported in several pathological conditions, 15 –17 and has been shown to correlate positively with the clinical resolution. For example, the SDF-1/CXCR4 cytokine-receptor axis has been described for HSPCs mobilization and recruitment to injured tissue. Despite its demonstrated importance, the exact mechanism(s) by which various growth factors cause the increased presence of circulating HSPCs is incompletely understood.
Acute fasting regimens have become popular and have shown profound impact on pro-longevity and tissue regeneration in diverse species. It has been proposed that diet-induced physiological cues likely influence stem cell biology. Caloric restriction and intermittent fasting positively affect diverse populations of HSPCs, notably by reversing age-related phenotypes and enhancing regeneration in various tissue-derived stem cells. 18 For example, short-term fasting augments intestinal stem cell function to form intestinal organoids, in young and aged mice, by inducing a fatty acid oxidation program. This enhancement of intestinal stem cell function was positively correlated with the duration of the fast. 19 Similarly, a restoration in the expression of critical transcription factors for self-renewal, was reported in murine muscle stem cells after multiple rounds of a short-term calorically restrictive diet designed to mimic fasting. 20 In blood, a fasting-mimicking diet (FMD) was also shown to rescue the decline in mesenchymal stem and progenitor cell number within BM. 20 Likewise, intermittent fasting or FMD promotes self-renewal of hematopoietic stem cells (HSCs) and induces protection against cytotoxic agents used in chemotherapy. 21 Moreover, studies using repeated rounds of FMD showed an increased in the number and proliferation ability of mesenchymal stem and progenitor cells in aged animals as well as rebalanced outputs from HSCs in aged mice. 21 After prolonged fasting in mice, results from cell-cycle analyses showed a significant induction of cell-cycle entry, of different populations of HSCs, indicating an induced increase in HSCs self-renewal proliferation without an increase in apoptosis. 21 In human FMD subjects, the percentage of LIN−CD45−CD184+ mesenchymal stem cells in PB showed a proportional increase at the end of FMD, with a subsequent return to baseline levels after refeeding. 20 However, because studies of HSCs have shown that induction of proliferation may sometimes be accompanied by an increased apoptosis, 22 it is important to note that this was not observed in prolonged fasting induced stem cells self-renewal proliferation. Future research studies on human subjects should be performed to compare the impact of fasting on stem cell biology and correlate this with observations in animal models. In humans, fasting is achieved by ingesting no or minimal amounts of food and caloric beverages for periods that typically range from 12 h to 3 weeks. Many religious groups incorporate periods of fasting into their rituals including Muslims who fast from dawn until dusk during the month of Ramadan, and Christians, Jews, Buddhists, and Hindus who traditionally fast on designated days of the calendar each year. Ramadan fasting (RF) constitutes a daily refrain from food and drink consumption, smoking, and oral administration of medications for a duration of 29–30 days, every year. The duration of fasting ranges from <12 h to as much as 18 h each day, depending on the season that Ramadan falls. This makes RF a form of prolonged intermittent fasting where both the quantity and quality of food eaten are altered. Several studies showed that the month of Ramadan can precipitate dramatic changes in sleep patterns and circadian rhythms. Moreover, RF can be associated with favorable physiological changes among healthy individuals, such as decreased body weight, significant changes in lipid profile, reduced oxidative stress and inflammation, and disease prevention. 23 –25
The intersection between RF and HSPCs biology has remained unclear and not well studied in human subjects. Therefore, we conducted, for the first time, an observational study aimed at evaluating HSPCs subpopulations in circulation in human PB during RF. Examining the influence of fasting on the physiological peripheral circulation of stem cells could improve the understanding of their biological role during this stressed period. These observations provide opportunities to define the role of dietary intervention in stem cell therapy. In the current study, circulating HSPCs were defined by the lack of expression of lineage (LIN−), mild to nonexpression of CD45, and the expression of the hematopoietic stem-cell marker CD34 and/or expression of the early hematopoietic stem-cell marker CD133.
Materials and Methods
Patient recruitment
This is a prospective observational study, conducted in the month of Ramadan during April–May 2022 in the city of Abu Dhabi, United Arab Emirates. Ten male volunteers were recruited via direct invitation to
Ethics approval for human samples and consent to participate
This study was approved by the Research Ethics Committee of ADSCC, Abu Dhabi, in accordance with the Helsinki Declaration. 26 Written informed consent was obtained from each participant in agreement with the rules of the Ethics Committee. The information was protected under the principles of confidentiality without revealing participants’ identities.
Circulating HSPCs isolation
Stem cell processing was performed as previously described, 27 with minor modifications. Briefly, PB-derived mononuclear cells (MNCs) and erythrocytes layers were separated, by Ficoll density gradient centrifugation (Ficoll-Hypaque; Sigma-Aldrich, ρ = 1.077 g/mL) at 800g, for 30 min, 21°C. PB-MNCs layer, at the interphase, was diluted 1:3 (v/v) with RPMI 1640, and centrifuged at 1,000g, for 10 min, 21°C, without brake. The erythrocytes layer was first diluted in 1:5 (v/v) with lysis solution (VersaLyse Lysing Solution, Beckman Coulter) for 15 min at room temperature (RT). Subsequently, the top layer was collected by centrifugation at 1,000g for 10 min, 21°C. This process was repeated two times until erythrocytes were completely removed. The pellet obtained after centrifugation at 1,000g for 10 min, 21°C was added to the pellet obtained during MNCs layer processing.
Flow cytometry
Circulating HSPCs were evaluated using a panel of monoclonal antibodies; Pacific Blue (P-B) conjugated-antihuman Lineage Cocktail (CD3, CD14, CD16, CD19, CD20, CD56) (LIN) (Biolegend, USA), phycoerythrin texas Red (ECD) conjugated-CD34 (Beckman Coulter, USA), allophycocyanin conjugated-CD133 (Beckman Coulter, USA), Krome Orange conjugated-CD45 (Beckman Coulter, USA), and 7-AAD. Briefly, the cells were suspended in phosphate buffer saline (PBS) and incubated with 10 µL fluorochrome‐conjugated antibodies and incubated for 30 min in the dark at 4°C. After incubation and washing, the cells were resuspended in PBS. Flow cytometric data were collected on a Navios EX device and analyzed with Kaluza C V.1.1 software (Beckman Coulter, USA). Fifty thousand events per sample were analyzed and compared with an antihuman IgG isotype-matched negative control. Viable HSPCs were defined as cells forming a cluster with low side scatter, low Lin staining, low-to-intermediate CD45 staining, and positive for CD34 and/or CD133. The number of HSPCs was expressed as absolute count using flow count fluorospheres (Beckman Coulter, USA) (Fig. 1).
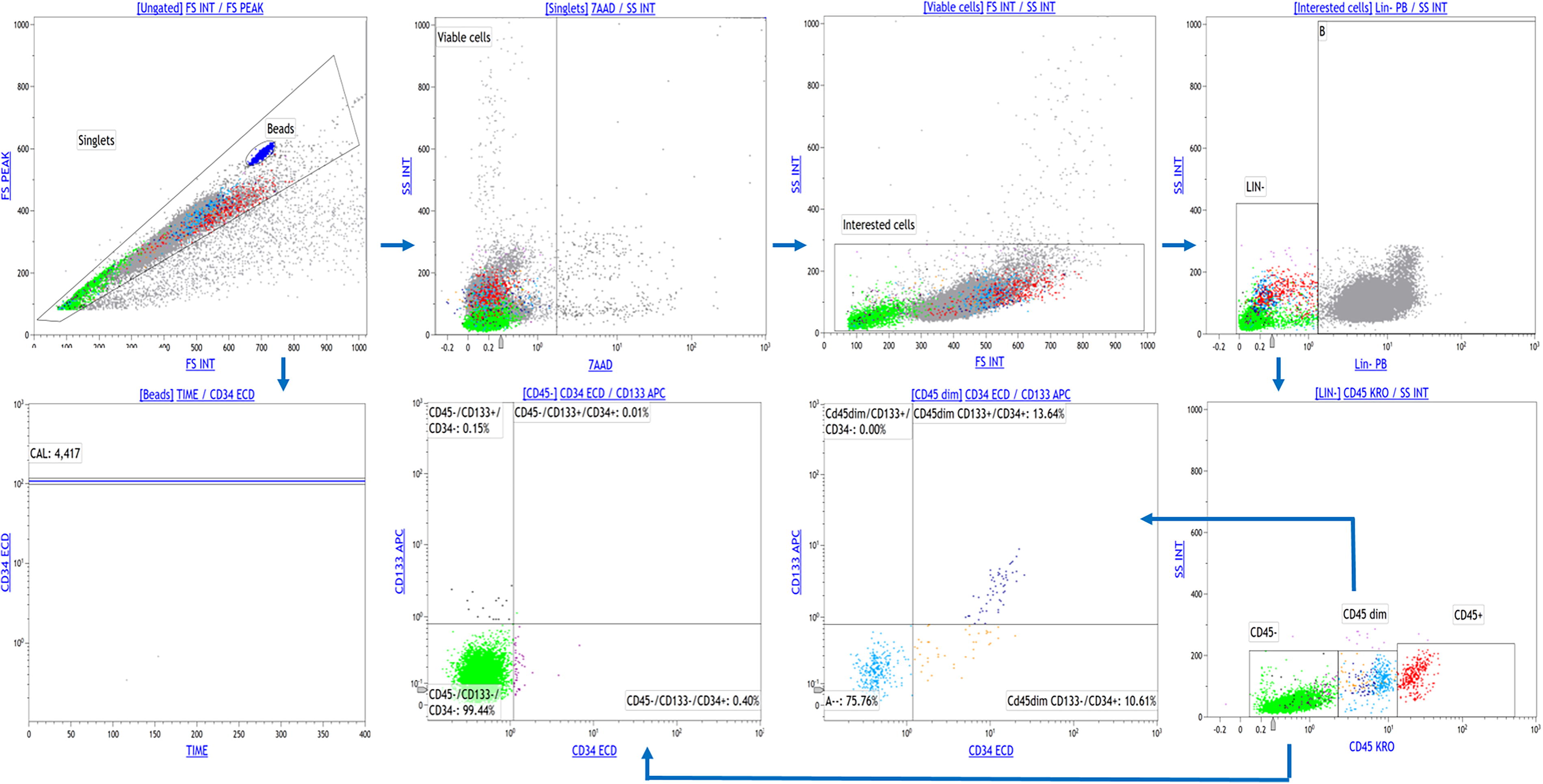
Gating strategy used for HSPCs analysis. HSPCs were defined as cells forming a cluster with low side scatter, low Lin staining, low-to-intermediate CD45 staining, and positive for CD34 and/or CD133. Flow cytometric data were collected on a Navios EX device and analyzed with Kaluza C V.1.1 software. Fifty thousand events per sample were analyzed. The number of HSPCs was expressed as absolute count using flow count fluorospheres. HSPCs, hematopoietic stem/progenitor cells.
Measurement of human serum growth factors
For serum human growth factors measurements, an aliquot (5 mL) of PB was collected in a vacutainer without anticoagulant. Sample tubes were left at RT for 30–45 min, and then were centrifuged at 2,000g for 15 min, 21°C. The obtained serum was aliquoted and stored at −80° C until use without repeated freeze–thaw cycles. A simultaneous quantification of 13 human growth factors: Angiopoietin-2 (Ang-2), Epidermal growth factor (EGF), Erythropoietin (EPO), Fibroblast growth factor (FGF), Granulocyte-colony stimulating factor (G-CSF), Granulocyte macrophage-colony stimulating factor (GM-CSF), Hepatocyte growth factor (HGF), macrophage-colony stimulating factor (M-CSF), Platelet derived growth factor AA (PDGF-AA), platelet derived growth factor BB (PDGF-BB), Stem cell factor (SCF), Transforming growth factor alpha (TGF-α), and vascular endothelial growth factor (VEGF) was determined, in triplicate, by using a 13-plex bead-based multiplex assay kit (LEGENDplex™ Human Growth Factor Panel), according to manufacturer’s instruction (Biolegend, USA). The serum measurement’s linearity was first checked on two samples of D0 and their respective at D10, D20, and D30, using three different dilutions (1:1, 1:2, and 1:4). All samples were diluted 1:2 and known standard concentrations of each analyte were run in parallel with experimental samples. The median fluorescence intensity for each of the analytes in the samples was measured using a Navios EX cytometer and then calculated by LEGENDplex V.8.0 software using the results of the sigmoidal standard curve as recommended by the manufacturer.
Immunofluorescence staining
Freshly isolated HSPCs cells were fixed in 4% paraformaldehyde (Sigma-Aldrich) for 20 min, permeabilized, and blocked with 0.3% Triton X-100, 1% BSA, and 10% normal donkey serum in 1X PBS for 45 min at RT. Subsequently, cells were incubated with primary antibodies diluted with 1% BSA in PBS overnight at 4°C. The primary antibodies used include the surface markers: fluorescein Isothiocyanate (FITC) conjugated-CD45 (IM0782U, Beckman coulter, 1:20), phycoerythrin (PE) conjugated-CXCR4 (A07409, Beckman coulter, 1:20), and the nuclear markers from the Human Pluripotent Stem Cell Marker Antibody panel plus (R&D Systems, Cat. No. SC009) including goat Immunoglobulin G (IgG) anti-hNANOG and anti-hOct-3/4, mouse IgG anti-hSOX2 and anti-h/mSSEA-4 used at a final concentration of 10 µg/mL. Secondary antibodies included Northern Lights antigoat IgG NL557 (R&D Systems, 1:200) and antimouse IgG NL493 (R&D Systems, 1:200) in PBS containing 1% BSA for 1 h incubation at RT. Finally, nuclei were counterstained with 10 µg/mL Hoechst 33342 solution (Sigma-Aldrich, USA). All steps were carefully performed to avoid washing away the cells in suspension. Images were obtained using a Lionheart FX automated microscope (Biotek, Agilent). Representative cells were acquired with the following settings: DAPI (377/447; Ex/Em), GFP (469/525; Ex/Em), Texas Red (586/647; Ex/Em), and the bright field channels, using 20× objective with optical zoom 3–6.
Chemotaxis assay
Migration of cells toward human plasma was assessed using a transwell migration assay. A total of 1 mL of RPMI containing 10% and 50% of pooled normal human plasma (obtained at D0 and D30) was placed in the lower chamber of a 24-well plate equipped with a 3 µm Transwell insert. Then 105 of pooled cells from D0 and D30 were treated with or without 50 ng/mL Mitomycin-C (Sigma-Aldrich, USA), suspended in 300 µL of RPMI medium, and loaded into the upper chamber. The plate was incubated at 37°C and 5% CO2 for 24 h. Since cells are nonadherent, then the migrated cells will drop into the media in the lower chamber. No presence of migrated cells on the bottom surface of the Transwell has been observed. Migrated cells were collected from the lower chamber and stained with PB conjugated-LIN and FITC-CD45, for 30 min in the dark at 4°C, and counted using Navios EX flow cytometry with Kaluza C V1.1 software. Ten thousand events were analyzed, and antihuman IgG isotype-matched negative control was used with each sample.
Statistical analysis
The Shapiro–Wilk normality test was used to determine the normal distribution of peripheral populations and soluble factor concentrations. Statistical significance between the groups of two-time points, for example, morning versus evening or D0 versus fasting days, were evaluated using t-tests and the Wilcoxon test, for paired values tests, when data passed or failed the normality test, respectively. The Kruskal–Wallis nonparametric analysis was performed to evaluate independent variables corresponding to the cell migration assay. Post hoc comparisons with Dunn’s test were applied. The Spearman correlation coefficient was used to estimate the correlation between cell subpopulations and soluble factors. All these statistical analyses were performed using GraphPad® Prism v8.4.3 (GraphPad® Software Inc., USA). The statistical data were considered significant if P < 0.05.
Results
Increase in the absolute counts of LIN− cell subsets during diurnal RF
First, we aimed to evaluate the effect of diurnal RF on the absolute counts of circulating LIN− cell populations. Distribution of LIN− cell subsets according CD45 expression from the MNCs cell populations of PB collected on D0-morning and D0-evening are represented (Fig. 2). The average absolute counts of LIN−CD45−, LIN−CD45dim, and LIN−CD45+ were significantly higher (47-, 4-, and 1.7-fold, respectively) on D0-evening than morning. Of note, LIN−CD45− was found to be the highest responsive cell subset to diurnal RF. No changes in the LIN− subsets of the control group have been significantly observed (data not shown).
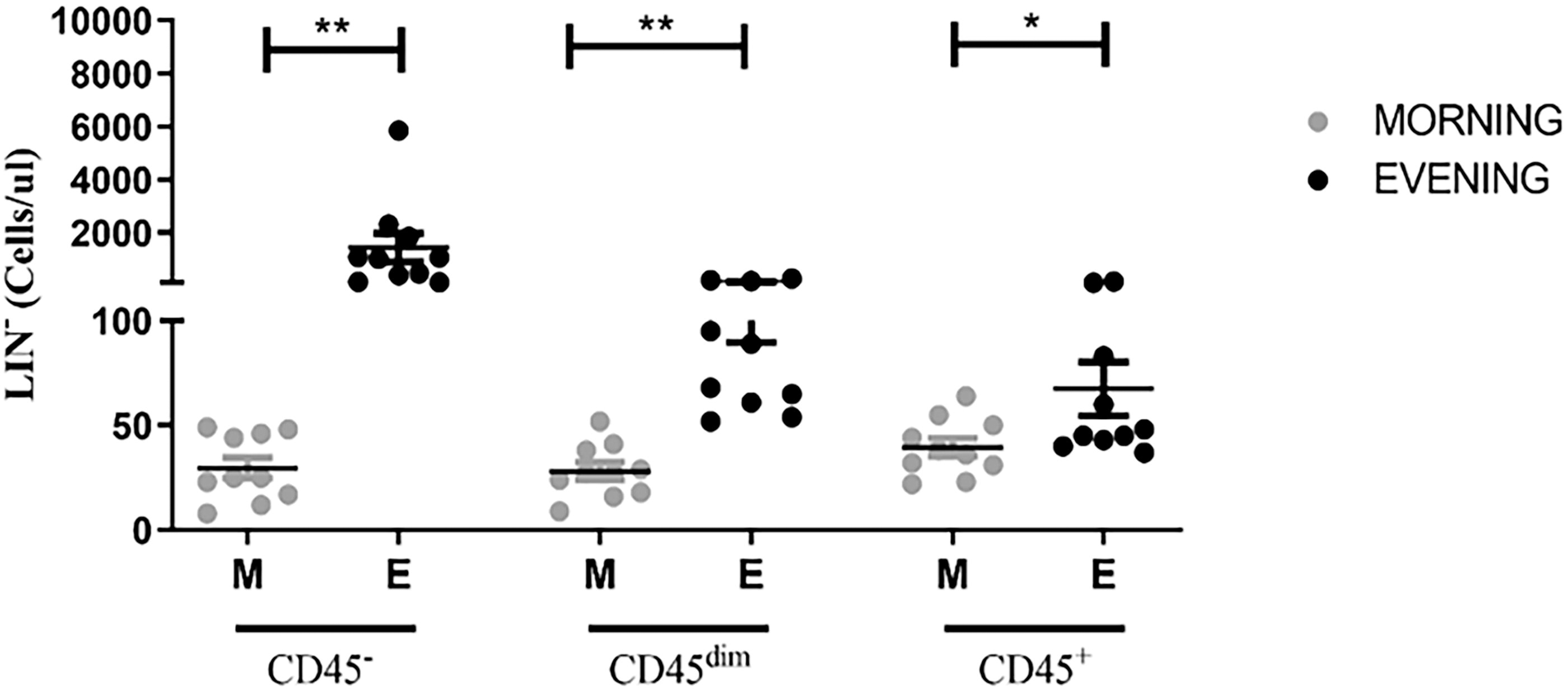
Baseline absolute counts of LIN− cell subpopulations according to CD45 expression. The asterisks indicate statistically significant differences among the groups (*P < 0.05; **P < 0.01); using the Wilcoxon test or paired t-test.
Cumulative effect of RF on LIN− cell subsets counts
We then proceeded to evaluate a possible cumulative effect of RF on the peripheral absolute counts of LIN− cells only for fasting group. LIN− average absolute counts were found significantly greater in evening than morning collection, accounting for >10-fold at each investigated fasting days D0, D10, D20, and D30 (Table 1). An overall increasing trend of LIN− cells through fasting in samples longitudinally collected on morning and evening was observed. A sharper increase was found on D30-morning (sixfold, P < 0.005) and D30-evening (eightfold, P < 0.0001) as compared with D0 average absolute counts. When CD45 expression was evaluated, the average absolute counts of LIN−CD45− and LIN−CD45dim were significantly higher in the evening than morning values at all investigated days. On the other hand, LIN−CD45+ average absolute counts were similar between both collections. Of note, LIN−CD45− cell subset was predominant, and the average absolute counts show longitudinal increases through fasting in both morning and evening collections with most intense amplitude on D30 morning and evening. LIN−CD45dim absolute counts tended to be higher through fasting, but the difference was statistically significant only on D30-evening. No significant variations were observed in morning and evening absolute counts of LIN−CD45+ (Table 1). In the control group, the average difference in the absolute counts of different LIN− cell subsets between D0 versus D30, on morning and evening samples was <10% (data not shown) which, is the acceptable threshold for error and has no significance. Based on these results, we exclude the control group in our further designed experiments in this study.
Absolute Counts of LIN− Cell Subsets of Both Morning and Evening Collection Times During Fasting Days
The asterisks indicate statistically significant differences among the groups (*P < 0.05; **P < 0.01) using a Wilcoxon test.
LIN−CD45− and LIN−CD45dim stem/progenitor cell subsets circulation during RF
To further characterize the primitive profile, we examined CD34 and CD133 stem/progenitor cell markers of LIN−CD45− and LIN−CD45dim subsets, for both collection times and during fasting days. Average absolute counts of LIN− cell subsets according CD45, CD34, and CD133 combined expression are listed in Table 2. Baseline average absolute counts (D0-morning) show LIN−CD45−CD34−CD133− and LIN−CD45dimCD34−CD133− as main cell subsets, whereas LIN−CD45−CD34+CD133+, LIN−CD45−CD34−CD133+, and LIN−CD45dimCD34−CD133+ were undetectable. However, a persistent significant increase between D0-morning and D0-evening average absolute counts was observed in all detectable subsets. Of note, LIN−CD45−CD34−CD133+, LIN−CD45−CD34+CD133+, and LIN−CD45dimCD34−CD133+ subsets tended to be higher, but the difference was not statistically significant. When compared D30 average absolute counts, a persistent increase was observed in evening values versus the morning ones (Fig. 3A, B). Interestingly, a sharp increase in D30-morning absolute counts has been similarly observed for LIN−CD45−CD34−CD133+, LIN−CD45−CD34−CD133−, and LIN−CD45−CD34+CD133+ subsets. Of relevance, LIN−CD45dimCD34−CD133+ showed a sudden raise to a high value at D30-evening only. Furthermore, morning- and evening-absolute counts of LIN−CD45−CD34+CD133− and LIN−CD45dimCD34+CD133− were proportionally increased with fasting days. In contrast, no cumulative effect of RF on LIN−CD45dimCD34+CD133+ and LIN−CD45dimCD34−CD133− was observed.
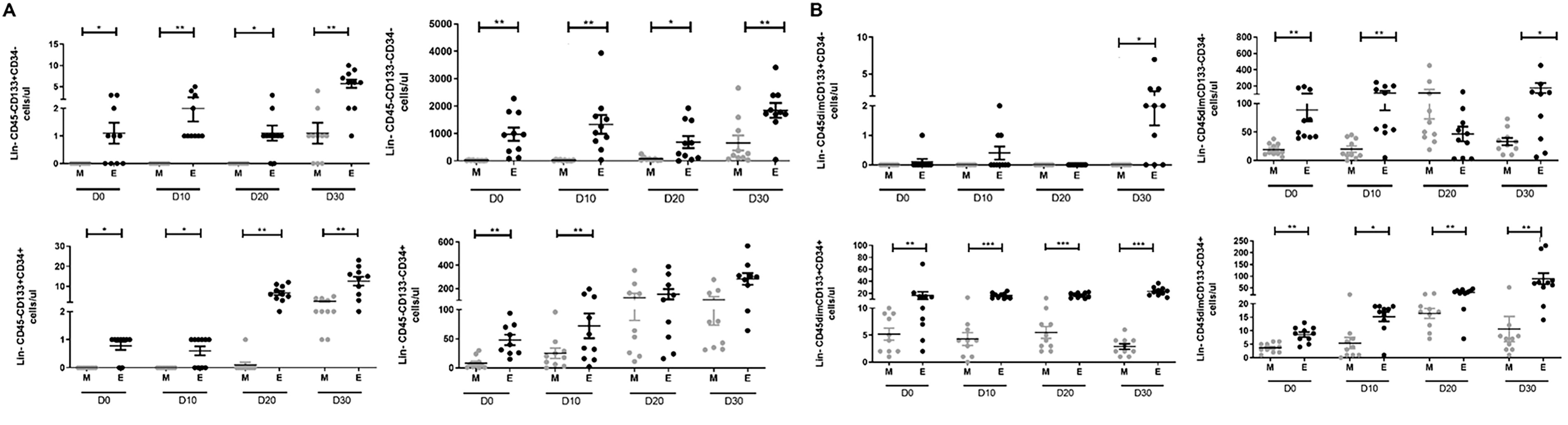
Changes in (Lin−CD45−/dim) cell subpopulations absolute counts for both morning and evening times during fasting days. Absolute counts (cells/µL) were obtained by flow cytometry analysis on
The Cumulative Ramadan Fasting Effect on LIN−CD45− and LIN−CD45dim Cell Subpopulations on D0 and D30 Time Points
The asterisks indicate statistically significant differences among the groups (*P < 0.05; **P < 0.01); using the (a) Wilcoxon test or (b) paired t-test.
RF-induced variations in the serum concentration of soluble factors
We measured the effect of a daily and cumulative RF on the circulating soluble factors levels in the serum of the enrolled healthy volunteers. Serum SCF and G-CSF levels were significantly upregulated in the evening time, as compared with the morning concentration in each investigated fasting day (D0; P < 0.005, D10; P < 0.005, D20; P < 0.005, D30; P < 0.0005, paired t-test) for SCF and P < 0.0001 (paired t-test) for G-CSF (Fig. 4A, B). In addition, a cumulative effect was progressively observed during fasting days, on average there was a two- and threefold increase in SCF and 1.5-fold increase in G-CSF levels at D30-morning and evening as compared with the basal level in D0, SCF; P < 0.005 (Wilcoxon test) and P < 0.0001(paired t-test), respectively, and G-CSF; P < 0.0001 and P < 0.0001 (paired t-test), respectively (Table 3). In contrast, EGF and ANG-2 levels were decreased between morning and evening of each investigated day (Fig. 4C, D) and showed the lowest concentrations in both timings at D30 as compared with the basal levels of D0, EGF; P < 0.005 and P < 0.005 (Wilcoxon test), respectively and ANG2; P < 0.0001 and P < 0.0001 (paired t-test), respectively (Table 3). Serum VEGF levels showed an oscillation pattern during fasting (Fig. 4E).

Soluble factors levels during fasting days. Serum concentration (pg/µL) obtained by Legendplex analysis on
The Cumulative Ramadan Fasting Effect on Soluble Factors on D0 and D30 Time Points
The asterisks indicate statistically significant differences among the groups (**P < 0.01; ***P < 0.001) using the (a) Wilcoxon test or (b) paired t-test.
VEGF, vascular endothelial growth factor.
Correlations between serum soluble factors and LIN− cell subsets during RF
Associations between soluble factors SCF, G-CSF, EGF, VEGF, ANG2, and both LIN−CD45− and LIN−CD45dim subpopulations were assessed between morning and evening time points on D0 and D30 (Fig. 5). Serum concentrations of ANG2 and SCF were positively correlated on D0-morning (r = 0.8659, P < 0.005). In contrast, between ANG2 and LIN−CD45dim, a negative correlation was observed (r = −0.6505, P < 0.05). In the evening, positive correlations were observed between SCF and ANG2 (r = 0.6951, P < 0.05), SCF and LIN−CD45− (r = 0.6869, P = 0.05), and LIN−CD45dim and LIN−CD45− (r = 0.8061, P < 0.01). Furthermore, at D30 morning, positive correlations were observed between SCF and LIN−CD45dim (r = 0.5719, P = 0.089) and SCF and G-CSF (r = 0.7295, P < 0.05), whereas negative correlations were observed between EGF and ANG2 (r = −0.6342, P < 0.01) and EGF and G-SCF (r = 0.6727, P < 0.05) on evening time.
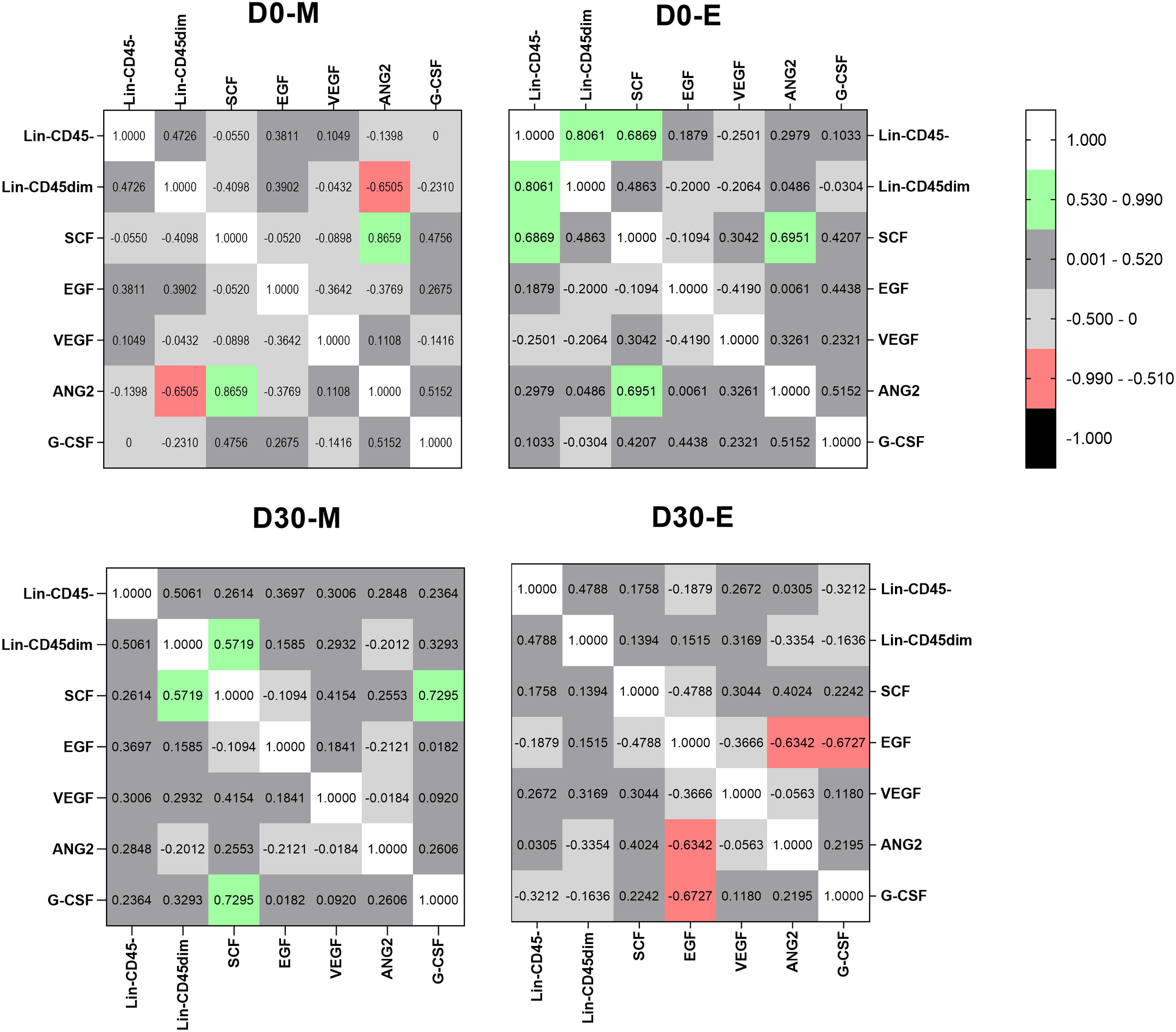
Correlations among soluble factors and LIN− cell subpopulations on D0 and D30. Correlation coefficient and statistically significant associations are highlighted in green (positive correlation) and in red (negative correlation). r = as Spearman Correlation coefficient. Statistical data were considered significant if P < 0.05.
RF stimulates LIN− cell subsets migration expressing CD184
Then, we evaluated the in vitro chemotactic activity of pooled serum obtained in D0-evening and D30-evening, on PB-derived LIN−CD45− and LIN−CD45dim cells of D30-evening. Pooled serum-induced cell migration, in a concentration-dependent manner (Fig. 6A). Cell migration ratio was observed to be stronger using serum (50%) obtained on D30 (0.35 and 0.31, for LIN−CD45− and LIN−CD45dim, respectively), as compared with that obtained on D0 (0.11 and 0.12 for LIN−CD45− and LIN−CD45dim, respectively). To evaluate whether the chemotactic activity was due to enhanced migration or increased proliferation, we repeated the assays in the presence of mitomycin C, an inhibitor of proliferation. Treatment with mitomycin C did not affect the plasma-stimulated cell migration of either the LIN−CD45− or LIN−CD45dim, indicating that the observed effect was not due to increased proliferation of the cells (Fig. 6B). Immunofluorescence staining of CD184 was evaluated and showed an intense plasma membrane staining in LIN−CD45− and LIN−CD45dim migrated cells (Fig. 6C). Furthermore, migrated cells showed a positive expression for SOX-2 and NANOG pluripotent stem markers but negative for SSEA-4 and Oct 3/4 (data not shown) (Fig. 6D).

Effect of RF on (LIN−) cell migration.
Discussion
Emerging evidence suggests that adult stem cells respond to cues from their environment, such as circadian rhythm, changes in metabolism, dietary signals, exercise, aging, infection, and disease, by altering their functional and biological properties. This allows adult stem cells to play a precise role in orchestrating and regulating not only short-range signals that maintain homeostasis within their tissue of origin but also long-range signals that integrate stem cell function with systemic physiology. Herein, we conducted the first human study that aimed at defining mechanistic insights into how dawn-to-dusk, dry, intermittent fasting, for 30 consecutive days can affect the differential kinetics related to stem cell populations in the PB of 10 healthy subjects practicing RF.
Data collected from numerous studies have indicated that the most primitive hematopoietic cells, do not express a variety of surface markers that are associated with the terminal maturation of specific blood cell types and reside in a lineage-negative (LIN−) subpopulation. Our data clearly show an increased number of circulating LIN− cells by the evening of the first day of fasting, and this tendency was continuously observed at each time point of this study. Interestingly, LIN−CD45− represents the highest mean absolute count values followed by LIN−CD45dim. To better understand if both populations include stem cells, we extend our study in defining different subpopulations by expressing and/or co-expressing two putative stem/progenitor markers, CD34 and CD133. Our LIN−CD45− and LIN−CD45dim fractions were composed of heterogeneous populations. We first identified a main population lacking the expression of lineage markers, CD45, CD34, and CD133, defined as LIN−CD45−CD34−CD133− and show a sustained increase during fasting. Although, detailed investigations on the molecular and functional characterization of this circulating population are rare, human circulating LIN−CD45−CD34−CD133− cell population has been described by Ciraci and colleagues, as hemangioblast-like cells endowed with developmental potential to differentiate into both hematopoietic and endothelial lineages in vitro and in vivo. 28 Indeed, this population has shown the ability to generate a significant number of CD45+ progeny in vivo, implicating their role in long-term immune cells maintenance. 28 The increase in the LIN−CD45−CD34−CD133− cell fraction observed in this study may be a response to compensate for changes in hematopoiesis during RF. It is well known that total number of leukocytes, granulocytes, lymphocytes, and monocytes were significantly decreased during Ramadan in comparison to pre-Ramadan. 29,30 In addition, cycles of diet that mimics fasting (FMD) are shown to be effective in promoting increases in mesenchymal stem and progenitor cells (MSPCs), which are CD45, CD34, and CD133 negative and likely contribute to the regeneration of various committed cells. 31 Brandhorst and colleagues reported an increase of LIN−CD45−CD184+ MSPCs in the PB of human FMD subjects. The percentage of MSPC in the PB-derived MNC population showed a trend to increase sevenfold at the end of FMD, with a subsequent return to baseline levels after re-feeding. 20 Consistent with this notion, our immunofluorescence results remarkably showed LIN−CD45−CD34−CD133− fraction positive for CXCR4 or CD184. Thus, we propose that LIN−CD45−CD34−CD133− should include an MSPCs subpopulation LIN−CD45−CD34−CD133−CXCR4+ that shows a similar tendency to increase during the fasting day and accompanied subsequently by a decrease return to baseline levels after re-feeding, shown in the morning values. Phenotypic characterization of the most widely accepted antigen panel and quantification of circulating MSPCs-derived MNCs population are needed for further investigations. 32
Our study clearly shows an increased response to fasting in LIN−CD45−/dim CD34+, CD133+, and CD34+CD133+ double positive cell fractions in the PB-derived MNCs population. Several studies have documented the existence of an inverse relation between blood glucose levels and the numerical and functional alterations of circulating CD34+CD133+ stem cells. 33 It was reported that hyperglycemia contributes to a 40% mean reduction in circulating progenitor stem cells levels, CD45dim CD34+, CD45dim CD133+, and CD45dim CD34+ CD133+ double-positive cells in PB. 34,35 In addition, it is widely accepted that stable hyperglycemia impairs CD45−/dim CD34+ cells from entering circulation from the BM to PB. 36 In line with these findings, a reduction of plasma glucose levels was followed by an increase of circulating CD45−/dimCD34+ cells and insulin supplementation was reported to mobilize these populations from the BM. However, whether the increase in circulating CD45−/dim CD34+ cell levels was due to decreased blood glucose levels rather than insulin itself is not clear. 37 Under this prospective and considering that fasting for longer duration shows a decrease in plasma glucose, due to an increase in glucose uptake into peripheral tissues and thereby increasing in the insulin sensitivity. 38 –41 We speculate that these reported changes in glucose metabolism during RF upregulate the levels of circulating progenitor cell fractions that we observed herein. Interestingly, loss of body weight due to caloric restriction from intermittent fasting has been shown to be positively correlated to the absolute numbers of circulating CD34+ cells. 42 Indeed, a meta-analysis that used data from 35 publications showed that RF caused a statistically significant decrease in weight (−1.24 kg for healthy male) by the end of Ramadan. 43 The difference in findings observed in this study between the late increase response of CD45−CD34+CD133+ and CD45dimCD34−CD133+ versus the progressive increase response observed in the CD45−/dimCD34+ subpopulations during RF could be explained by the effect of weight reduction in individuals that have fasted for 30 days. This may also explain the increase in the subpopulations, CD45−CD34+CD133+ and CD45dimCD34−CD133+ observed at the later time points. Unfortunately, we have not conclusively shown that regulation of these subpopulations during fasting is a direct result of lower glucose or weight loss. Knowledge about HSPCs circulation and the regulatory processes governing physiological protection of organs during fasting is scarce. However, it is tempting to speculate that increases in circulating CD45−/dimCD34+ cells in response to fasting is a precautionary response to protect various tissues and organs from metabolic insult, triggered by stimulating these cells to maintain tissue plasticity in the face of low glucose due to fasting. 44 One noteworthy finding of this study was that the responsive cell population LIN−CD45−/dim CD34+CD133+ to RF, feature some phenotypic characteristics of endogenous human very small embryonic-like (hVSEL) stem cells reported by Ratajzack and colleagues since 2006. 45 hVSELs have been described as small cells of 3–5 µm in size, spherical in shape with high nucleo-cytoplasmic ratio and open type euchromatin. At present, hVSELs have been reported in various human tissues as a resident pool of primitive and quiescent stem cells that could actively mobilized from the BM into the PB under stress conditions. 46 A supportive study to our data showed that caloric restriction without malnutrition, positively affects the number of BM- and spleen-residing as well as PB-circulating VSELs and HSPCs. 47 These findings provide corroboration for our experimental data showing that LIN−CD45−/dimCD34+CD133+ may be identified as this rare circulating VSELs that may contribute to regenerative responses or homeostasis regaining by migrating into PB. Interestingly, our immunofluorescence data on LIN−CD45−/dim, show the expression of two main pluripotent markers well expressed in hVSELs, SOX-2 and Nanog. However, imageStream analysis, confocal microscopy, and quantitative RT-PCR to further characterize our experimental cell populations are needed to better identify the identity of cell populations involved in this study.
Furthermore, there is compelling evidence for the role of the circadian rhythm in regulating properties and function of adult stem cells, 48 including circulating stem cells. 49 Indeed, based on circadian rhythmicity dictated by signals from the sympathetic nervous systems, several clinical research studies were designed to optimize the collection time that coincides with the peak of CD34+ egress into the circulation. 9 In humans, circulating LIN−CD45−CD34+ and their progenitors exhibit robust oscillatory behavior, peaking 5 h after the initiation of light and reaching a nadir 5 h after darkness. 46 Consistent with these findings, we consider the changes shown at D0 may be the circadian oscillation of HSPCs but we cannot exclude a complementary effect of a 1-day fasting specifically, that no significance changes have been observed in the control group.
Hematopoietic and HSPCs development and function involves a complex array of cytokines and growth factors. In previous reports, it has been reported that the mobilization of HSPCs populations into the circulation followed the kinetics of plasma elevation of several chemo/cytokines such as SDF1, SCF, VEGF, and G-CSF. Interestingly, the present study showed a progressive increase response in SCF and G-CSF serum levels across the fasting time course. This suggests that RF may be a stronger stimulus for increasing these two chemo-attractants levels in circulation. Moreover, in our present study, the serum levels of SCF were found to be positively correlated to LIN−CD45− and LIN−CD45dim absolute counts at D0 and D30 of fasting. This is presumably a consequence of an increased level of SCF and G-CSF that can promote the mobilization of LIN− HSPCs. Previous studies reported SCF plasma levels were directly correlated to progenitor cell mobilization. 50 Additionally, both SCF and G-CSF were found positively correlated at D30 fasting when LIN− HSPCs content had already peaked in the circulation. Another explanation for the relative increase in HSPCs could be attributed to a combination effect of SCF and G-CSF in promoting cell mobilization. Indeed, several clinical trials have proven a synergistic action between these cytokines in increasing the mobilization of PB progenitor cells for transplantation over that seen with G-CSF alone. 51,52 Interestingly, the present study showed a significant progressive decrease in VEGF serum level. However, no significant correlation was observed between circulating VEGF level and the increase absolute counts of LIN− HSPCs subsets in the peripheral circulation. Hence, we hypothesize no significant role for VEGF in the HSPCs recruitment. The reduction in the EGF and ANG-2 levels during RF could be attributed to the decreased oxidative stress during Ramadan 53 and the decreased reactive oxygen species that mediates the functions of these cytokines, which are responsible for the production of pro-inflammatory cytokines including IL-6, IL-8, and GRO-α. Moreover, studies have demonstrated that EGF is an important biomarker in obesity 54 and may also have metabolic and weight-regulating effects. 55
This study has a few limitations that need to be considered. It is an observational longitudinal study with small sample size. This could have reduced the power of the study, but not the novel finding on the changes in circulating HSPCs levels during RF. Further multicenter studies with larger cohorts are needed to perform in‐depth evaluations on the here‐described correlation of circulating HSPCs levels and circulating growth factors with RF. Notwithstanding our sample size, we were able to collect PB from the same donor, at two different time points on each of the study days and at each successive 10-day period of fasting. The fact that we performed flow cytometry on fresh sample, not subjected to freezing-thawing to avoid the negative effects of cell damage was strongly considered. This strengthens our results about the effect of a daily and cumulative fast on these cell populations. The in-vitro functionality of investigated stem cell subsets, for example, self-renewal ability, colonies formation, differentiation has not been done yet. Finally, we were unable to fully characterize the SDF1-CXCR4 axis to ensure its role in triggering the circulation of stem cell subsets to the PB. Answering these questions in future studies will help to determine whether circulating HSPCs subsets could play a potential role and identify new mechanisms responsible for the changes in stem cell physiology during RF.
Conclusion
The current study shows an enhanced presence of primitive stem cell subsets in the peripheral circulation during RF. We report persistent changes in the level in PB during a daily fast and across the time points within the 30-day fasting period. In addition, our data show a differential in the kinetic response of stem cells subsets as well as a positive association with some growth factors implicated in mobilization to PB. We speculate that a change in the mobilized HSPCs populations is contributing to tissue-specific fasting-induced adaptations. These findings are novel; we are the first to report this association and it raises intriguing questions about the role of circulating HSPCs and fasting-related changes of cellular and metabolic pathways. Defining the fasting-related mechanisms leading to changes in circulating HSPCs and their multisystemic effects should be the focus of additional future work.
Footnotes
Author Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This work received funding from the Abu Dhabi Stem Cells Center.