
Abstract
Abstract
Background:
Although much attention is currently directed to studying microbial biofilms on a variety of surfaces, few studies are designed to study bacterial growth on surgical suture. The purpose of this study was to compare the kinetic development of Staphylococcus aureus and Enterococcus faecalis on five surgical suture materials and to clarify factors that might influence this growth.
Methods:
Pure cultures of S. aureus and E. faecalis were incubated with five types of suture for four days using either tissue culture medium or a bacterial growth medium. Suture-associated bacteria were quantified daily. In selected experiments, the bacterial growth medium was supplemented with heparin, a substance known to promote S. aureus biofilm formation. The ultrastructure of S. aureus biofilm developing on braided suture was studied with scanning electron microscopy.
Results:
Staphylococcus aureus and E. faecalis were recovered in greater numbers (typically p < 0.01) from braided than from monofilament suture, and the numbers of bacteria were greater (often p < 0.01) on sutures incubated in bacterial growth medium rather than tissue culture medium. Addition of heparin 1,000 U/mL to silk or braided polyglactin 910 suture incubated three days with S. aureus resulted in greater numbers of bacteria on day one but not on subsequent days. Scanning electron microscopy showed a maturing
Conclusions:
Bacterial growth was favored on braided vs. monofilament suture, and heparin enhanced bacterial adherence after day one, but not at subsequent times. Staphylococcus aureus adhered to suture material and formed a structure consistent with a bacterial biofilm.
The impact of SSIs on health care in the U.S. is substantial. Summarizing data from 90% of all U.S. hospitals in 2005, de Lissovoy et al. [4] reported that SSIs occur in ∼2% of surgical procedures and account for ∼20% of all health-care-associated infections, resulting in almost one million additional inpatient hospital days and $1.6 billion in excess cost annually. A single SSI increases the length of stay an average of 9.7 days and the cost by $27,288. Collection of these data relied on a single diagnostic code number to identify cases [4], and thus the data have low sensitivity although high specificity. They reflect approximately 125,000 SSIs per year, a figure substantially less than the 274,000 cases per year that are estimated by the U.S. Centers for Disease Control and Prevention using more inclusive criteria [1]. Nonetheless, it is evident that SSIs have a substantial impact on patient morbidity and on health care costs in the U.S.
Surgical suture can become contaminated by bacteria from the environment or from the patient's normal flora, and substantial effort has been targeted at identifying risk factors as well as preventative measures, such as preoperative skin preparation, perioperative oxygen inspiration, intranasal decontamination (to eradicate S. aureus), and antimicrobial prophylaxis [2,5–7]. It is accepted widely that suture material provides a nidus for microbial adherence and wound contamination. Early literature documented that bacterial adherence to suture depends on the microbial species and the suture composition and structure, with braided sutures more prone to colonization than monofilament ones [8–10].
There is increasing awareness that bacteria colonize surfaces as communities of organisms within a biofilm, consisting of a population of bacteria enclosed in an extracellular polymeric substance composed of proteins, lipids, polysaccharides, and extracellular DNA [11,12]. In the clinical setting, bacterial biofilms contaminate indwelling medical devices such as catheters, orthopedic implants, and artificial heart valves. These infections are notoriously difficult to eradicate, and resolution often requires removal of infected tissue or the device. There has been some effort to decrease microbial contamination of surgical suture by coating the material with an antimicrobial agent [13,14], but these technologies have not gained widespread acceptance.
Using an in vitro model system, experiments were designed to clarify the effect of several dependent variables on the growth of S. aureus and E. faecalis as biofilms on suture materials. Dependent variables included the type of suture and the nutrient environment. Taking advantage of developments in high-resolution scanning electron microcopy (SEM), the developing ultrastructure of S. aureus biofilm formation on suture material also was investigated.
Materials and Methods
Bacterial cultivation on surgical suture
Staphylococcus aureus RN6390 is a virulent strain that has been used to study interactions on silicone elastomer (Silastic®) catheters [15] and in cultured human cells [16–18]. Enterococcus faecalis OG1RF is a plasmid-free strain, often used as the parent strain for genetic manipulations of this species [19]. Enterococcus faecalis VA1128 is a clinical isolate.
Suture material was purchased from Ethicon, Inc., Johnson & Johnson (Somerville, NJ) and included 3-0 silk (black braided), absorbable, clear, monofilament polydioxanone (PDS*II®), 3-0 blue monofilament polypropylene (Prolene®), and 3-0 and 4-0 undyed, braided polyglactin 910 (Vicryl®). Suture was handled aseptically, cut into 1-cm segments, placed in six-well dishes containing 3 mL of growth medium per well, and incubated for as long as four days at 37°C with gentle rotation. Growth medium was either a standard tissue culture medium consisting of Dulbecco Modified Eagle Medium supplemented with 15% fetal bovine serum and 4 mM L-glutamine or a biofilm growth medium containing 66% tryptic soy broth supplemented with 0.2% glucose (TSB/glu) [20]. In some experiments, the medium was supplemented at 1,000 U/mL with heparin sodium from porcine intestinal mucosa (grade 1-A; Sigma-Aldrich Catalog No. H3393, St. Louis, MO).
The growth medium was inoculated with 5 × 107 bacteria from a pure culture that had been cultivated overnight in tryptic soy broth, washed, and resuspended to the appropriate concentration in 100 microliters of Hank's Balanced Salt Solution. The growth medium was changed daily. Immediately after fresh medium was added to a given well, 100 microliters of the fluid was cultured quantitatively to determine the concentration of residual planktonic bacteria (not associated with the biofilm) bathing the sutures in the well, and a suture was subsequently removed. The numbers of planktonic bacteria verified that the numbers of these bacteria transferred inadvertently in the medium bathing a suture did not contribute significantly to the numbers of viable bacteria recovered from the suture itself after sonication (data not shown). On removal from the well, each suture was placed in 3 mL of sterile saline, sonicated (∼50 J at 100% amplitude for 5 sec) using a VibraCell 130 W 20 kHz Ultrasonic Processor (Sonics and Materials, Newtown, CT), then quantitatively cultured on tryptic soy agar supplemented with 5% sheep red blood cells.
Data are reported as the log10 viable bacteria/cm of suture. In selected experiments, bacterial suspensions from sonicated suture were examined by epifluorescent microscopy after staining with BacLight™, a viability stain (Molecular Probes, Eugene, OR).
Statistical analysis
Two groups were compared by the unpaired Student t-test, and more than two groups were examined by one-way analysis of variance with Fisher post hoc testing. Significance was set at p < 0.05.
Scanning electron microscopy
Staphylococcus aureus was incubated with 3-0 polyglactin 910 suture in biofilm growth medium for four days as described above. At daily intervals, two or three sutures were processed for scanning electron microscopy (SEM) as described [21] with minor modifications, with all aldehyde fixatives and buffers obtained from Electron Microscopy Sciences (Hatfield, PA). Briefly, suture segments were rinsed; fixed overnight in a mixture of 2% glutaraldehyde, 2% paraformaldehyde, 4% sucrose, and 0.15% alcian blue in 0.15 M sodium cacodylate buffer; washed in cacodylate buffer; postfixed in 1% osmium tetroxide and 1.5% potassium ferricyanide in cacodylate buffer; rinsed; and dehydrated through a graded ethanol series, followed by critical point drying with CO2. Samples were coated with 1 to 2 nm of platinum with an argon ion beam coater (Denton DV-502, Denton Vacuum, LLC, Moorestown, NJ) and viewed with a Hitachi S-4700 field emission scanning electron microscope (Tokyo, Japan) operated at 2 to 3 kV. Images were collected using Quartz PCI software (Quartz Imaging Corp., Vancouver, BC, Canada) and stored in TIFF format.
Results
Bacterial colonization of suture materials
As noted in Figure 1, the cultivation medium influenced the growth/survival of both S. aureus and E. faecalis on the five types of suture, with greater numbers of suture-associated bacteria generally being recovered from samples incubated in TSB/glu than those grown in tissue culture medium. Also, greater numbers of bacteria were recovered consistently from braided than from monofilament suture. Curiously, there were many situations in which the numbers of suture-associated bacteria stayed relatively constant over the four-day duration of the experiment; e.g., monofilament suture incubated with S. aureus in tissue culture medium and with E. faecalis OG1RF in TSB/glu (Fig. 1). Subsequent experiments used only S. aureus because this is the organism most frequently involved in SSIs [2] and because results with this organism were most consistent in the two growth media.

Numbers of viable bacteria adherent to five suture materials (3-0 silk, 3-0 PDS*II, 3-0 polypropylene [Prolene®], 3-0 braided polyglactin 910 [Vicryl®], and 4-0 Vicryl®) after inoculation with Staphylococcus aureus (
To assess bacterial viability in suture-associated biofilms, bacterial suspensions from sonicated 3-0 silk and 3-0 braided polyglactin 910 sutures incubated with TSB/glu were stained with BacLight™ (Invitrogen Corp., Carlsbad, CA), with viability assessed daily (Table 1). Bacterial viability did not vary significantly (62% to 72%) over the three days of the experiment.
Average of three series of 100 contiguous bacteria, where the series did not differ by >15%.
Effect of heparin on colonization of braided suture with S. aureus
Preliminary experiments indicated that adding heparin to biofilm growth medium enhanced bacterial clumping in broth alone and on braided polyglactin 910 suture incubated in broth (Fig. 2). Thus, heparin 1,000 U/mL was added to suture incubated with S. aureus for three days, and the numbers of suture-associated bacteria were counted daily (Fig. 3). Heparin had no significant effect on the numbers of S. aureus recovered from 3-0 silk suture incubated in tissue culture medium, although heparin was associated with greater numbers of bacteria after the first day, but not subsequent days, of incubation with 3-0 silk and 3-0 braided polyglactin 910 in TSB/glu (Fig. 3).

Staphylococcus aureus cultivated overnight in 66% tryptic soy broth supplemented with 0.2% glucose in a six-well dish at 37°C with gentle rotation, showing the gross morphologic effect of heparin 1,000 U/mL on growth. The dishes in the upper photographs contain only S. aureus and medium, where unsupplemented broth has a uniform turbidity, and heparin supplementation fosters visible bacterial clumping. Lower photographs show three pieces of 3-0 braided polyglactin 910 suture added to each well, with markedly clumped bacteria obscuring the sutures in medium supplemented with heparin.
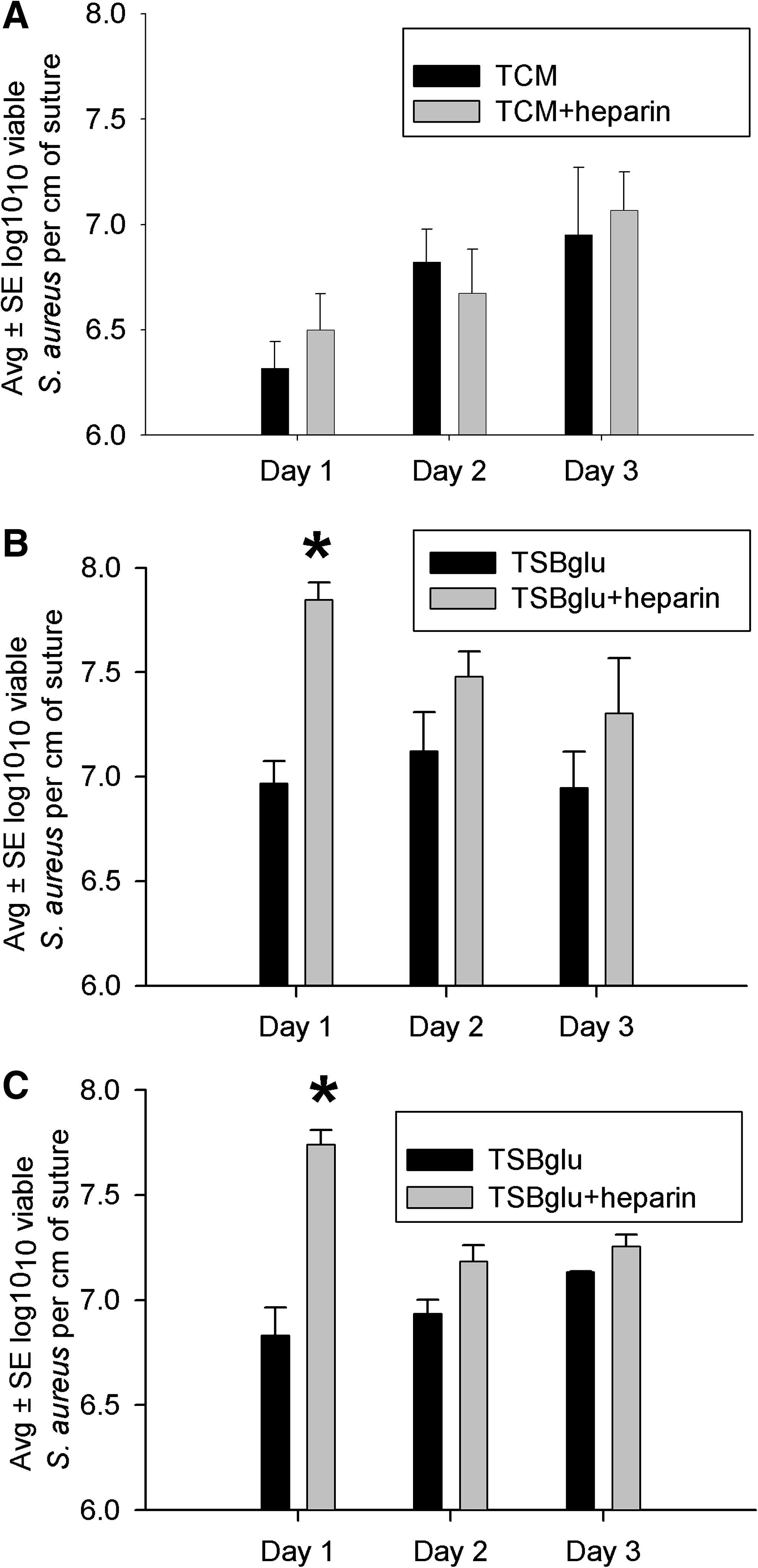
Effect of heparin 1,000 U/mL on Staphylococcus aureus biofilms cultivated three days on silk suture in serum-supplemented tissue culture medium (
Scanning electron microscopy of S. aureus biofilms on braided polyglactin 910 suture
The ultrastructure of S. aureus biofilm development on braided polyglactin 910 suture is presented in Figure 4. After 24 h of incubation, clusters of cocci were seen among amorphous material, and suture-adherent bacteria often were connected by fibrillar strands (Fig. 4A). Some bacterial cells appeared to cluster between individual suture strands, making these cells difficult to resolve by SEM (Fig. 4B). After two days, the biofilm appeared more developed and was characterized by larger clusters of bacteria embedded in amorphous and fibrillar material (Fig. 4C), and some bacteria appeared to acquire a denser coating of extracellular material (Fig. 4D). After three or four days, the biofilm appeared more consolidated, and it seems reasonable to suggest that many bacterial cells were deeply embedded in the extracellular material (Fig. 4E, F).
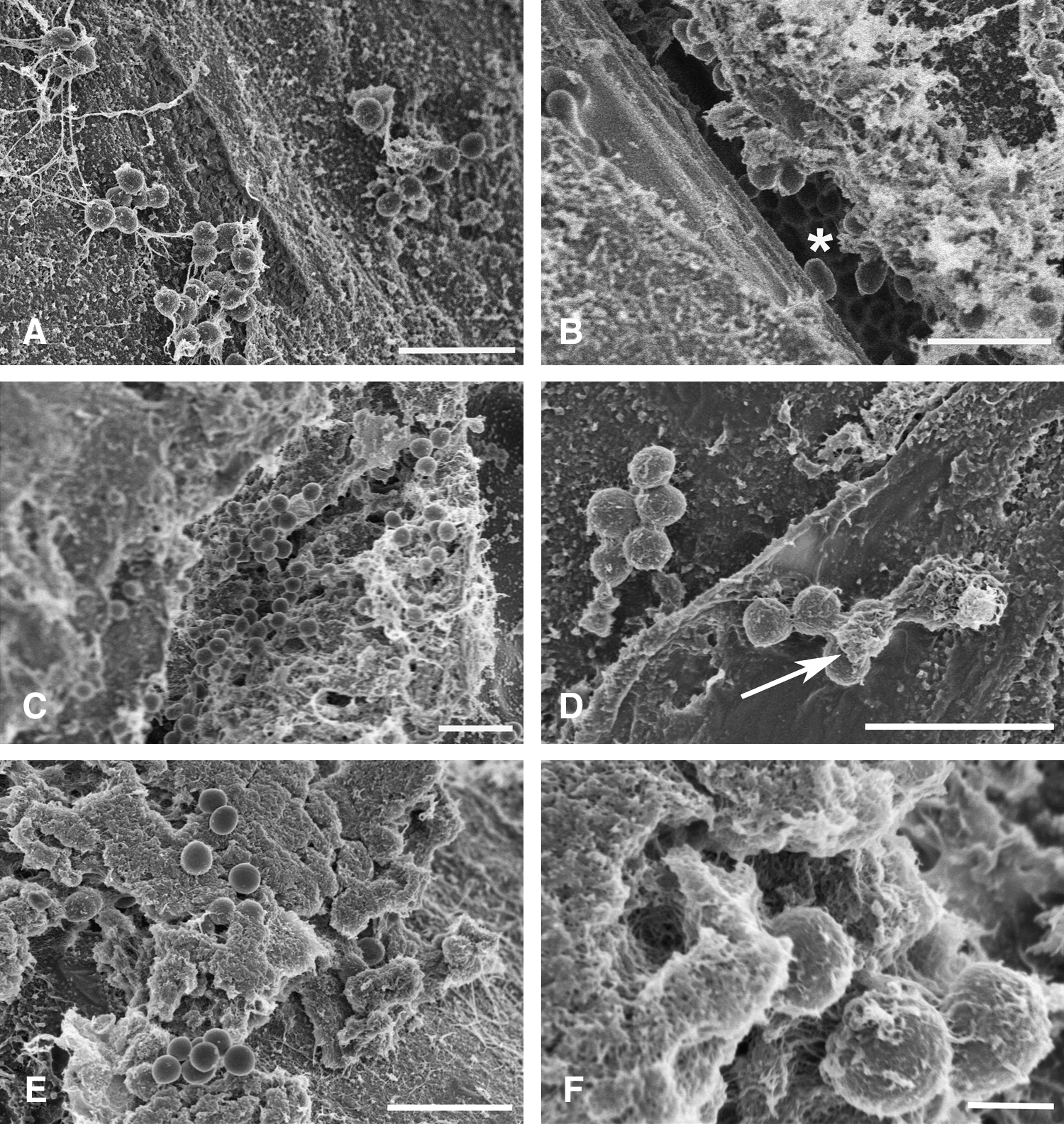
Scanning electron microscopy of 3-0 braided polyglactin 910 suture incubated in 66% tryptic soy broth supplemented with 0.2% glucose with Staphylococcus aureus for one day (
Discussion
Bacterial biofilms are attracting more attention in clinical medicine. Numerous disease states have been associated with bacterial growth as a biofilm, such as urinary tract infections, catheter-related infections, middle ear infections, dental infections, endocarditis, infections in cystic fibrosis patients, and infections of indwelling devices such as joint prostheses and heart valves [11]. It has long been accepted that the presence of suture material increases the risk of infection, and SSIs, often associated with suture materials, are likely associated with bacterial growth as a biofilm.
Chu and Williams [10] studied adherence of radiolabeled S. aureus and Escherichia coli to ten suture materials. These authors summarized their results by saying that bacterial adherence depended on a number of factors, including filament configuration (monofilament vs. braided) and the chemical nature of the suture, as well as the suture coating material, with the coating likely to be more influential (with absorbable but not nonabsorbable suture) than physical configuration. In the present study, S. aureus and E. faecalis appeared to adhere preferentially to braided rather than monofilament suture, and this effect was more noticeable in experiments conducted in TSB/glu rather than serum-supplemented tissue culture medium, indicating that the nutrient composition plays a role in bacterial adherence to suture. It is tempting to speculate that braided suture is more prone to bacterial adherence because of its larger, more complex surface area, but further study will be needed to confirm that conclusion.
We had expected that biofilm growth would increase steadily over time, but that was not always the case; e.g., S. aureus cultivated on braided suture in TSB/glu. Here, bacterial viability staining indicated that, although the majority of suture-adherent S. aureus cells were viable, a relatively consistent proportion (∼30–40%) were not. Thus, changes in the proportion of nonviable and viable bacteria did not appear to explain the recovery of consistent numbers of S. aureus from braided suture over time. The growth medium was changed daily, and this medium likely contained bacteria that had separated from the suture-associated biofilm. Chu and Williams [10] noted that bacterial adherence to suture is a dynamic process, and it is known that bacteria (either as single cells or clusters) can actively (or passively) leave the biofilm by a process termed “dispersion” or “dissolution.” This activity is thought to represent the last step in biofilm development, as bacteria return to the planktonic state. Little is known about the mechanisms involved in dispersion, and these mechanisms likely differ with the bacterial species, the environment, etc. [22]. A caveat to the experiments herein is the fact that the in vivo environment contains a variety of substances, such as albumin, fibrinogen, fibronectin, etc., that may coat the suture and influence bacterial adherence in ways that were not relevant to these in vitro studies.
There is evidence that heparin can stimulate formation of S. aureus biofilms [20]. We noted that heparin increased adherence of S. aureus to suture materials after one day of incubation in TSB/glu, but not at later time points. The mechanism of this effect is not clear and deserves further study; it may be related to pH changes with bacterial growth in the presence of a fermentable sugar.
Ultrastructural observations of bacterial biofilms on suture materials are rare. More than 25 years ago, Chu and Williams [10] used SEM to observe sutures contaminated for 60 min with S. aureus or Escherichia coli and described the presence of an extracellular “adhesive medium that has the appearance of tangled polymeric fibrils of polysaccharides or branching sugar molecules that extend from the bacterial surface and form a felt-like glycocalyx surrounding an individual cell or colony of cells and is secreted by bacteria.” Edmiston et al. [13] viewed S. aureus and E. coli on a variety of suture materials and noted that bacterial adherence was easily visible. We have extended these findings and observed that early bacterial adherence was accompanied by noticeable amounts of amorphous material and relatively long fibrillar strands that often appeared to connect the bacterial elements. As the biofilm developed over time, the extracellular material appeared to consolidate and to encase the bacteria in a thick blanket of extracellular material. Assuming this dense material exists in vivo, antimicrobial agents might not penetrate it easily, thus contributing to the greater antibiotic resistance often noted with biofilm-associated infections.
Conclusion
Bacterial growth on suture material appeared to have the characteristics of biofilm formation. Further study of these cells within the biofilm may shed light on the relative antibiotic resistance of biofilm-encased bacteria.
Footnotes
Author Disclosure Statement
This work was supported in part by U.S. National Institutes of Health Grant R01 AI058134 (to GD) and in part by funds from the Department of Surgery, University of Minnesota, Minneapolis (to DH). No competing financial interests exist.
Parts of this work were carried out in the Institute of Technology Characterization Facility, University of Minnesota, which receives partial support from the National Science Foundation through the National Nanotechnology Infrastructure Network program.