
Abstract
Abstract
Background:
Neutrophil dysfunction has been documented after injury in animals and human beings. This review evaluates the relative effects of the hormonal and endotoxin response to injury on immune resistance.
Method:
Review of the pertinent English-language literature.
Results:
In volunteers given total parenteral nutrition, neutrophils demonstrate a robust response to leukotriene B4 but none to zymosan/activated serum or the bacterial metabolite formyl-methionyl-leucyl-phenylalanine (FMLP). This finding suggests subclinical exposure to activated complement and FMLP that does not occur during enteral feeding. Additional evidence of neutrophil activation is the release of lactoferrin to the same degree with the two routes of feeding. When normal volunteers are challenged with endotoxin, uniform impairment of the neutrophil response to chemotactic stimuli except LTB4 is demonstrated. Epinephrine increases the total circulating neutrophil pool for a few hours, whereas when cortisol is administered, the neutrophil counts continue to increase through 6 h. A combined epinephrine and cortisol infusion extends the half-life of neutrophils. The role of genomic and central nervous system control through the vagus nerve also is reviewed.
Conclusion:
Normal volunteers have provided insight into the stress response to infection that is understood only partially.
In 1968, as a response to the increasing clinical interest in acute respiratory failure, the National Science Foundation sponsored a symposium on pulmonary insufficiency following nonthoracic injury [6,7]. Several events allowed posttraumatic lung dysfunction to gain widespread clinical recognition. These included the better management of “acute renal failure” (now called acute kidney injury), along with the development of technology that allows accurate measurement of the partial pressure of gases in the blood. As a consequence of improvements in the evacuation of severely injured soldiers from the rice paddies of Vietnam to sophisticated resuscitation hospitals, a large population of otherwise-healthy patients with “shock lung” provided a challenging clinical problem and created a need to improve survival. Many fatally injured patients survived the initial hours and days after their injury, only to die several weeks later from complications of multiple organ dysfunction syndrome, often as a result of nosocomial infection and overwhelming sepsis [8].
Metabolism and Nutrition
The complex relations among nutrition, metabolism, and host responsiveness were a puzzle with many moving pieces. Data from individuals suffering from chronic malnutrition during World War II revealed higher rates of bronchopneumonia than in normal-weight subjects [9]. Similarly, patients with peptic ulcer disease suffering from chronic gastric outlet obstruction leading to substantial weight loss showed the operative mortality rate to increase in proportion to the percentage of preoperative weight loss [10]. These data indicated an important relation between nutrition and the host's ability to fight infection. Clinical studies of delayed hypersensitivity reactions likewise showed a correlation between nutrition and survival, but these studies failed to identify a mechanistic defect relating the malnutrition to susceptibility to infection.
In 1975, Doctor G. Tom Shires moved to New York from Seattle and became Chairman of the Department of Surgery at what is now the Weill Medical College of Cornell University. The Department had had eight chairmen or acting chairs of surgery over the preceding decade. Research was a dark and receding memory, because a previous chairman had attempted to develop basic research in the Department but instead brought notoriety to the institution and a weak clinical program for the surgeons under his supervision. Doctor Shires made many personnel changes so rapidly at so many levels that upon JMD's return to the institution after a two-year fellowship at the National Institutes of Health (NIH), he found the department almost unrecognizable. These staff additions, made to re-create a research-oriented faculty of competent scientists and meticulous surgeons, were crucial to dispelling the skepticism left by Shires' distant but well-remembered predecessor. Not only was the Department enlarged to accommodate clinically skilled surgeon–educators (Doctors Peter C. Canizaro, Malcolm O. Perry, and P. William Curreri) and a new group of academically oriented residents (Doctors Roger W. Yurt, Philip S. Barie, and David N. Herndon), but new clinical programs were established. A burn unit was opened that became busy instantly, caring for the many victims of apartment fires in New York City. This unit was supported by an NIH Center grant. Doctor Curreri, who provided clinical resources and supported the awarding of an NIH burn center grant in 1979, became the first Principal Investigator. The history of the burn center has been reviewed recently [11].
The clinical interest in the metabolic demands of burn patients sparked a productive line of inquiry by Doctor David Herndon, who had just completed a burn fellowship at Brooke Army Medical Center, Fort Sam Houston, Texas, under Doctor Basil Pruitt, Jr. Herndon had developed an animal model to assess metabolic demand, providing insight into the importance of metabolism in the surgical care of these patients [12–14]. This interest in the metabolic demands of burn patients focused many of us on this clinical problem, including Doctor Stephen F. Lowry. Interest in metabolism evolved with the purification and identification of tumor necrosis factor-alpha (TNFα) in 1985, which appeared to be a crucial cytokine mediator of these changes [15]. Lowry was arguably the first to recognize the importance of this molecule and its relevance to surgical problems and infections.
Because malnourished individuals were believed to be more susceptible to infection than well-nourished subjects, nutritional support, whether intravenous or enteral, was thought of as a panacea for the host's ability to fight infection. Moreover, the metabolic response to injury was known to create such a high nutritional requirement that a trauma victim could easily deplete his or her nutritional reserve, leading to a state similar to what was seen in World War II. Total parenteral nutrition (TPN) had been developed at the University of Pennsylvania by a number of workers starting with Doctor Jonathan Rhoads in the 1950s [16,17], culminating in 1968 with a paper by Doctor Stanley J. Dudrick demonstrating its effectiveness in the growth and development of litter-mate dogs [18]. This was an important advance in treating the hypermetabolic, critically injured patient. However, studies were beginning to be published supporting the differential benefit of enteral feeding over TPN.
Normal Volunteer Model
Whereas most laboratories interested in the metabolic response to injury were using animal models [19], the study of normal volunteers provided unique insights. The subjects were recruited by public advertisements under an approved Institutional Review Board protocol. They were healthy males ranging in age from 18 to 30 years and within 10% of ideal body weight. After a thorough clinical evaluation and admission to the Clinical Research Center (CRC), the volunteers were fed a standard isocaloric diet (either enteral or parenteral) calculated to maintain their body weight (Fig. 1). A bolus of endotoxin from a uniform lot, used for more than 20 years in similar types of experiments, was administered over a 5-min period, which resulted in a classical response to endotoxin, with malaise, fever, and, in some instances, nausea and vomiting [20,21]. Lowry saw the value of this model and adapted it quickly to address the clinical questions posed by the identification of TNFα [20].
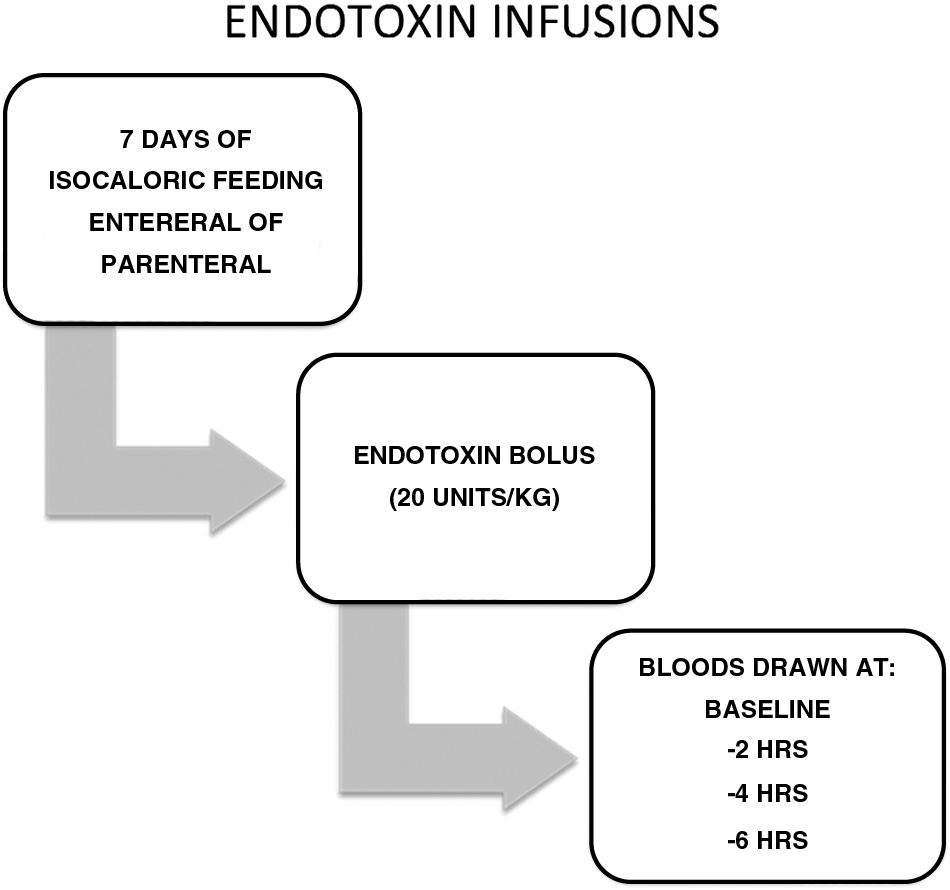
Protocol for experimental endotoxin infusion.
Route of Feeding and Neutrophil Function
Neutrophil function (complement activation, peripheral cell counts, and chemotactic responsiveness) was measured at baseline and then at 2, 4, and 6 h after endotoxin infusion given one week after the volunteers were admitted to the CRC and started on isocaloric nutrition with either enteral feeding or TPN. Chemotaxis was used because this is a complex activity employing both cell-surface receptors and intracellular integrity to generate movement toward a stimulus. Three chemoattractants were tested. The first was zymosan, consisting of protein–carbohydrate complexes derived from yeast cell walls, combined with activated serum (ZAS) [22], which contains activated complement (C3a and C5a). The second was a small amino acid, formyl-methionyl-leucyl-phenylalanine (FMLP) [23], and the third was leukotriene B4 (LTB4) [24]. The FMLP molecule is a potent chemoattractive by-product of metabolism that can be found in the supernatant fluids of cultures of many bacterial species, whereas LTB4 is a lipid mediator derived from cell membranes and generated by stimulation of neutrophils. It can enhance recruitment of cells despite an injury that already has attracted neutrophils.
The most important observation in this study was that in the TPN group, neutrophils demonstrated a robust response to LTB4 but none to ZAS or FMLP [25]. This suggested that during feeding, the patients given TPN had subclinical exposure to activated complement and to FMLP that did not occur during enteral feeding.
A number of hypotheses can be offered addressing why this occurs. The first possibility is that the intravenous catheters of patients fed parenterally were contaminated, causing bacterial inoculation and complement activation. Another possibility is loss or disuse atrophy of enterocytes, allowing the establishment of gut bacteria in the circulation (e.g., by translocation) and resulting in activation of complement and production of FMLP. These possibilities are not mutually exclusive [26].
Additional evidence of neutrophil activation was the release of the enzyme lactoferrin from intracellular granules (Fig. 2). This release while the cells were still in the circulation is part of a priming process. There was no difference in lactoferrin release with the two routes of feeding. However, simple tests looking at neutrophil numbers support the hypothesis of exposure to stimuli during parenteral nutrition. When the volunteers were stimulated with endotoxin, there was no evidence of complement activation by serum measures in the TPN group. Neither was there neutrophilia in response to this stimulus, which is consistent with our knowledge of bone marrow recruitment, as the complement by-products are potent mediators of bone marrow recruitment to the circulation (Fig. 3).

Plasma lactoferrin more than doubled after endotoxin infusion.
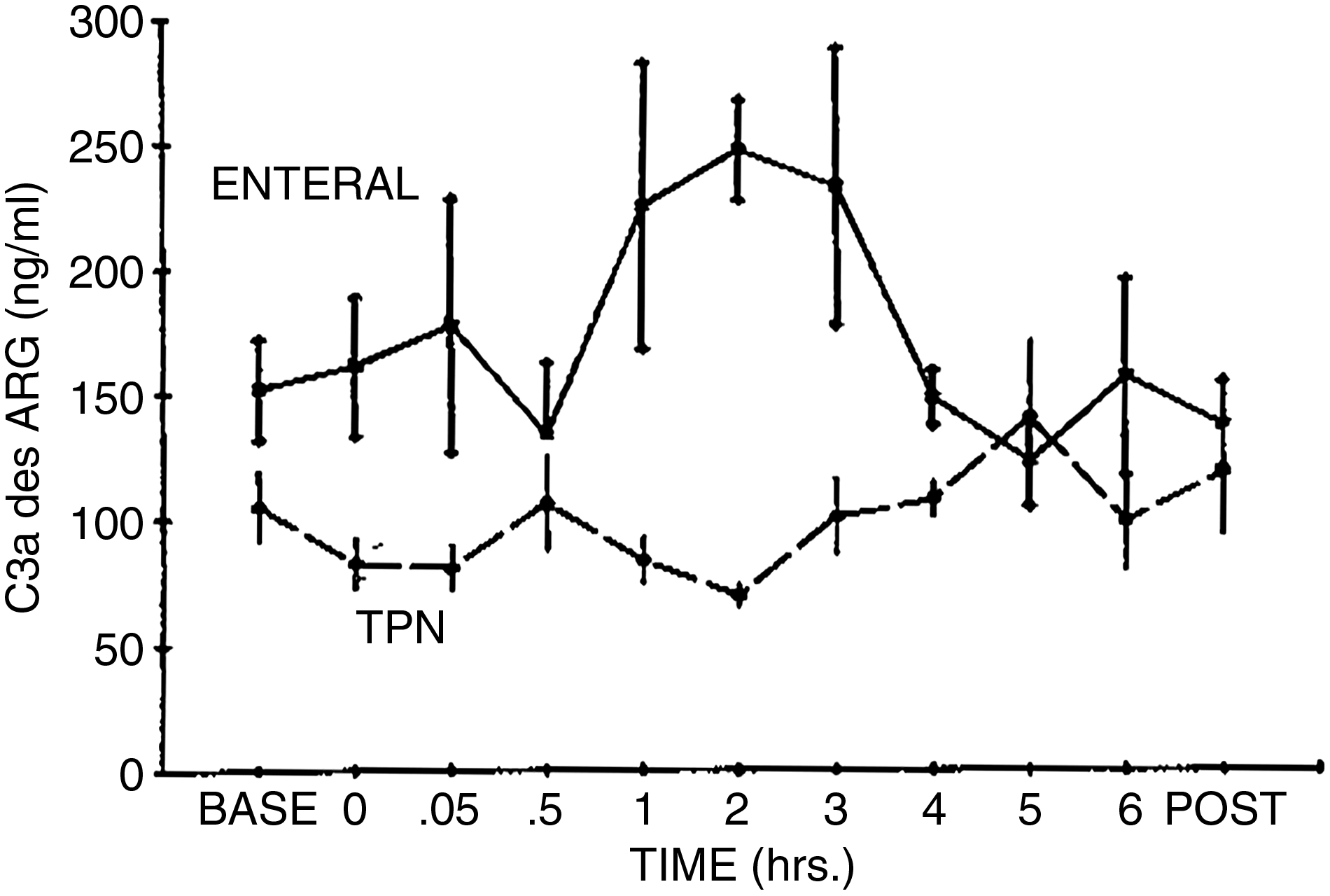
Changes in plasma complement factor 3a desARG concentrations after endotoxin infusion in enterally and parenterally fed volunteers. The C3a concentrations were significantly higher in the enterally fed group prior to infusion (p<0.02) and from hour 1 through hour 4 after the infusion compared with the TPN group (p<0.01). (Reprinted with permission from reference 28.)
This series of tests in normal volunteers described a differential activation pattern of neutrophils that was not appreciated fully. The “kamikaze” cell had different signaling mechanisms that could modulate intravascular activation. Subsequent study showed that the neutrophils had multiple membrane receptors that could cause up- or down-regulation as the cell was stimulated.
When normal volunteers were challenged with endotoxin [27], uniform impairment of the response to chemotactic stimuli except with LTB4 was demonstrated. Furthermore, neutrophil generation of LTB4 was increased at 2 h after infusion. In vitro incubation of cells from normal subjects exerted similar effects, reducing the chemotactic response to all stimuli except LTB4. Prior incubation with TNF had a facilitatory effect on the synthesis of LTB4.
Effect of Epinephrine and Cortisol on Neutrophils
A second group of normal volunteers was studied to evaluate the effects of “stress” hormones. These individuals were admitted to the CRC for a two-day isocaloric diet and then stimulated with epinephrine, cortisol, or both [28]. Normally, circulating neutrophils from healthy adults are in equilibrium with neutrophils adherent to the vascular endothelium. Additional neutrophils are 10- to 14-fold more abundant in the bone marrow than in the circulation. Consequently, adequate reserves of “troops” to respond to “an invading army” of infectious agents always are present at the time of injury. The recruitment of bone marrow is stimulated by a number of signals, including complement. It also is altered by the release of the stress hormones. The neutrophils then leave the circulation by rolling along the endothelial wall, becoming adherent, and then entering the tissues by squeezing between endothelial cells.
Epinephrine can demarginate all circulating neutrophils and thereby increase the total circulating neutrophil pool. The epinephrine response occurs without affecting the host's ability to eliminate damaged cells from the circulation or to recruit new cells from the bone marrow. To conduct this study [28], the volunteers received a 6-h infusion of epinephrine at 3 ng/kg/min (Fig. 4). Blood samples were harvested every 2 h. Neutrophil numbers increased steadily so that by the 4-h point of the infusion, the neutrophil count had reached its maximum. When cortisol was administered to volunteers at a rate of 3 mcg/kg/min for 6 h, the neutrophil counts continued to increase throughout this time. Cortisol increases the half-life of neutrophils by reducing cell–cell interaction (adhesion).

Protocol for epinephrine and cortisol infusions.
A final group of volunteers had a 6-h infusion of epinephrine followed by a combined epinephrine and cortisol infusion [28]. Using the formula for half-life (decay), the impact of cortisol on the lifespan of the neutrophil in the circulation was calculated. The total neutrophil pool reached after epinephrine infusion consists of circulating and marginated neutrophils. After the 6-h infusion, the number of neutrophils in the circulation and the number recruited from the bone marrow were calculated using the following formula: A=A0 e−0.693(t)/t1//2. The half-life of neutrophils was extended from 6.6 h to 10.4 h by cortisol. This is similar to the half-life calculated using radiolabeled cells [29].
Summary and Conclusions
Current thinking about neutrophil action maintains that the basics of neutrophil responsiveness include the classic steps of rolling, adhesion, and transmigration [30]. Within the past decade, these steps have been updated to include enhanced knowledge of how neutrophils become attached to endothelial cells, arrest at the site of inflammation, and transmigrate by one of two means: Classic paracellular (through the gaps between cells) and transcellular (through the endothelial cell itself) (Fig. 5) (see below). Attachment (or tethering) depends on the dynamics of integrins, which can quickly (<1 sec) switch from “inactive” to “active” conformations or from “active” to inactive” [31]. These integrins then interact with their endothelial cell-bound ligands such as intercellular adhesion molecule 1 and vascular cell adhesion molecule 1. Neutrophil surface molecules interact with platelet endothelial cell adhesion molecule 1 on the endothelial cell surface to mediate evasion by the neutrophil [32]. The neutrophil will be activated by macrophage-1 antigen and in some cases will transmigrate by either paracellular or transcellular action [30].
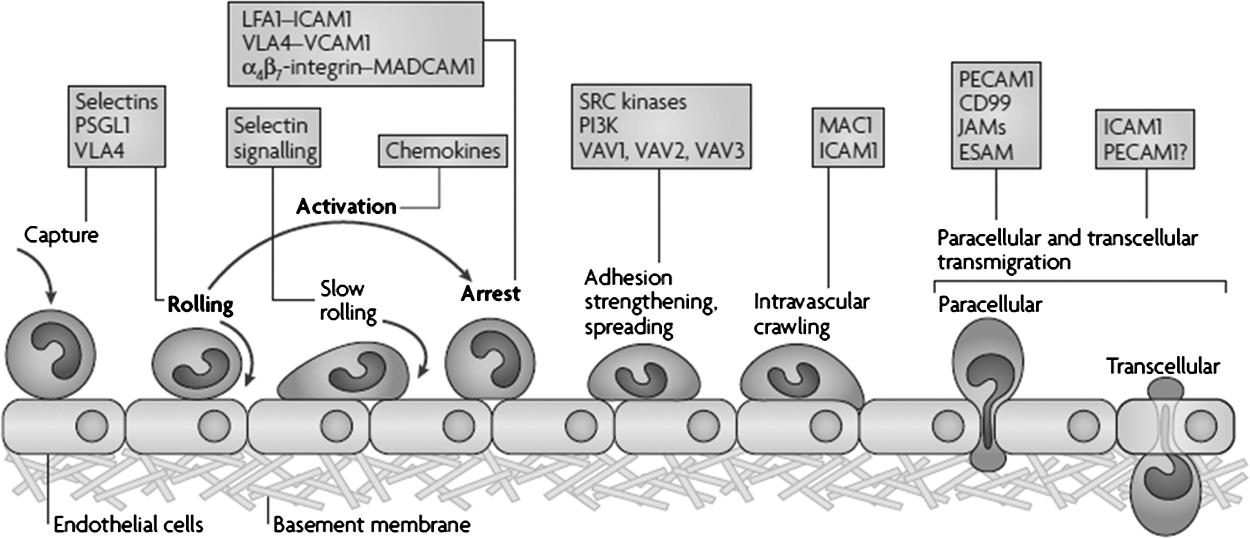
Updated leukocyte adhesion cascade. The original three steps are shown in boldface type: Rolling, mediated by selectins; activation, mediated by chemokines; and arrest, mediated by integrins. Progress has been made in defining additional steps: Capture (or tethering), slow rolling, adhesion strengthening and spreading, intravascular crawling, and paracellular and transcellular migration. Key molecules involved in each step are indicated in boxes. ESAM=endothelial cell-selective adhesion molecule; ICAM=intracellular adhesion molecule 1; JAM=junctional adhesion molecule; LFA1=lymphocyte function-associated antigen 1 (also known as α1β2-integrin); MAC1=macrophage antigen 1; MADCAM1=mucosal vascular addressin cell-adhesion; PSGL1=P-selectin glycoproteinligand 1; PECAM1=platelet/endothelial-cell adhesion molecule 1; P13K=phosphoinositide 3-kinase; VCAM=vascular cell-adhesion molecule 1; VLA4=very late antigen 4 (α4β2-integrin). (Reprinted with permission from reference 30.)
Complications of surgery, including sepsis, systemic inflammatory response syndrome (SIRS), and acute respiratory distress syndrome (ARDS), continue to frustrate medical professionals. The normal volunteer model enabled research that was lacking in other models, as evidenced by studies in dogs that did not show a neutrophil response to epinephrine [33,34], whereas human subjects exhibited signs of increased neutrophil mobilization [28]. It is clear now that catecholamines are crucial to the regulation of the immune system, at least partially because lymphoid organs are innervated predominately by the sympathetic system [35]. This relation is a target of clinical interest in preventing SIRS by blocking the effects of epinephrine [36].
In one of the bolder uses of the healthy volunteer model, nutritional support of patients in the peri-operative period was explored in 1988. Two groups of volunteers were created; for one week, these volunteers were fed either an oral diet to maintain body weight or TPN without a lipid supplement. An important result was that chemotaxis in response to the peptide FMCP and activated serum was decreased but not that in response to LTB4. The underlying mechanism for this finding was not elucidated, but a 2009 study found that parenterally fed mice have depleted pulmonary lymphocyte populations [37]. Because lymphocytes are a source of LTB4, and given that humans mirror the lymphocyte depletion observed in mice, these results may be related. Proper function of the mucosal immune system is of vast importance in preventing ARDS. Improper balance of pro- and anti-inflammatory signals in the form of chemokines and cytokines is connected to neutrophil participation in ARDS.
Reflecting on the central dogma of molecular biology, it can be understood that the protein expression, and therefore the cell activities, of the neutrophil are controlled at the genetic level. Doctor Lowry was part of a team that investigated the genetic changes in circulating leukocytes after severe trauma. Differences were reported in the amount and duration of genetic changes in complicated and uncomplicated recovery [38]. These changes include both the up-regulation of some genes, such as NB1 (cluster of differentiation molecule [CD]177), MMP8 (neutrophil collagenase), LTF (lactotransferrin), and HP (haptoglobin), to increase innate immune system activity; and down-regulation of genetic markers for antigen presentation and T-cell activation. These findings suggest that regulation of the immune system after injury is handled at the level of the individual cell, but the authors concluded that the “the mechanisms responsible for complicated clinical recovery after severe injury remain incompletely elucidated” (Fig. 6).
Genomic storm: Refining the immune, inflammatory paradigm in trauma. (
A target for clinically meaningful treatment is more important than a complete understanding of the mechanisms of recovery after trauma. To this end, the work of Xiao et al. is contested by research with a much different approach. Instead of thinking from the cellular level, perhaps the early findings of Doctor Lowry concerning TNFα and the changes in neutrophil activity can be accounted for better by a central signaling system. In a Journal of Clinical Investigation article, Doctor Kevin J. Tracey proposed that the nervous system is crucial to the regulation of the immune system after injury [39]. These findings, and work extended by Tracey protégé Doctor Jared Huston [40], indicate that the vagus nerve (CN X) is important in the inflammatory process, as demonstrated by vagal nerve stimulation that decreased cytokine production, and vagotomy that resulted in increased TNFα synthesis. A possible link is suggested between ancient techniques, such as acupuncture, and the modern understanding of medicine. Acupuncture creates measurable changes in vagus nerve activity, which may help explain its clinical effects. It may be that not only does Tracey's research suggest a potential target for clinical treatment, but it also may be a mechanism to explain some of the tenets of ancient Chinese medicine.
These two approaches to the investigative process give us glimpses of what are most likely two sides of the same coin. The genes, both up- and down-regulated, are telling in that they show a pattern of dynamic control of the immune system at the level of the individual cell. At the same time, evidence for central control of the immune system after injury reveals another set of potential therapeutic targets. Likely these systems work together with many other factors to create the clinical scenario. These mechanisms evolved well before the time of ICUs and operating rooms; the systems of the human body were meant to address only infection or injury that was survivable without external influences. The treatment of the severely injured patient is something the body's mechanisms were not intended for: Bringing a patient back from beyond a brink from which he or she cannot themselves return. It is this monumental task, involving incredibly complex and dynamic mechanisms, that is explored with these two approaches. The work of Doctor Lowry and his protégés had, and will continue to have, a central position in the understanding of surgical metabolism and sepsis.
Footnotes
Author Disclosure Statement
No competing financial interests exist.