
Abstract
Abstract
Background:
The early diagnosis of viral reactivation after kidney transplantation (KTX) is an unsolved problem. Survey of virus-specific T-cell responses may identify patients at risk for viral reactivation. We therefore quantified virus-specific CD8+ T-cells to evaluate their potential predictive value for viral reactivation and infection in KTX patients.
Methods:
We quantified the virus-specific responses of CD8+ T-cells for CMV, EBV, HPV and HHV in 23 patients undergoing KTX for 6 mo after transplantation. We enumerated T-cells for 36 virus-specific binding peptides and five different human leukocyte antigen (HLA) alleles through the binding of Class I iTAg major histocompatibility complex (MHC) tetramers. The patients' pre-operative serologic status for CMV and CMV-specific CD8+ T-cell numbers were correlated with one another (p=0.0046).
Results:
Three patients had clinical CMV disease and all three remained or became CMV-tetramer-positive for at least one HLA allele during follow-up. Three of the four patients with viral infections caused by or reactivations of viruses other than CMV were initially negative for CMV-specific CD8+ T-cells but became CMV-positive. Most of the patients who were initially CMV-tetramer positive also had tetramer-positive T-cells specific for Epstein–Barr virus (EBV); human papillomavirus (HPV)-6b, -11, -16, or -18; or human herpesvirus (HHV)-8. All of the patients who developed viral disease other than that caused by CMV remained or became positive for at least one binding peptide that was specific for a virus not directly related to the clinical features of a viral disease.
Conclusion:
Patients who were positive for any virus had a significantly greater risk of developing complications of viral disease during the 6-mo follow-up period in the study (p=0.026), suggesting a general susceptibility to viral reactivation. The evaluation of virus-specific CD8+ T-cells may prospectively help to identify patients at risk for viral reactivation after KTX.
V
Without antiviral prophylaxis, the incidence of CMV disease after KTX ranges from 20%–60%, with the disease typically occurring within 1–4 mo after transplantation [1]. Routine prophylactic application of ganciclovir or valganciclovir can significantly reduce the incidence of CMV disease, but is not a universally appropriate approach. Experience with high-risk patients (seronegative recipients of organs from seropositive donors) has show that antiviral prophylaxis reduces only partly the prevalence of CMV disease after transplantation. Although antiviral treatment can be effective in controlling viremia, from 5%–15% of infected patients still experience CMV disease when the antiviral drug is withdrawn [2]. Additionally, long-term anti-viral treatment can have side effects, some of them severe, such as bone marrow ablation and hepatotoxicity [8]. Beyond this, the cost-benefit ratio for the universal application of antiviral prophylaxis is currently insufficient to merit its use [3]. Nevertheless, extended prophylaxis with valganciclovir in high-risk patients has been shown to be beneficial within the first two years after transplantation (Improved Protection Against Cytomegalovirus in Transplant [IMPACT] study) [4].
The absence of complications of CMV infection in long-term transplant recipients is reflected by stable numbers of virus-specific T-cells, which do not differ from those in healthy CMV-positive controls. However, reactivations of CMV can contribute to age-related changes in the CD8+ T-cell compartment [5], which are of special importance in the immunocompromised subpopulation of transplant patients. By contrast, clinical symptoms during the first months after KTX may be correlated with a decrease in CMV-reactive CD4+ T-cell numbers, with a possible subsequent increase in viral load [6]. Because individual immune response and CMV replication are balanced in asymptomatic carriers of the virus, the availability of antiviral T-cells might serve to characterize a patient's CMV status.
Furthermore, in patients who have undergone stem cell transplantation, the quantification of virus-specific T-cells can be useful for differentiating those with sufficient, insufficient, and absent T-cell activity, suggesting such T-cell activity as a diagnostic tool [7–8]. Preliminary data on the quantification of virus-specific T-cells are also available for patients having heart and lung transplantation [9] because the T-cell response in solid-organ transplant recipients differs from the T-cell recovery in patients having bone-marrow stem cell transplantation, the currently available results for bone-marrow transplant patients cannot be applied to patients having solid-organ transplants without critical reflection. Furthermore, T-cell activity seems to play an important role in different viral diseases after transplantation [10,11], with virus-related complications not being limited to CMV and possible also occurring with other types of viruses. For example, the correlation between CMV and HHV infection after transplantation is well known, and the activation of HHV-6 often coexists with CMV infection in organ-transplant patients [20,21]. Epstein-Barr virus infection is known to be a major risk factor for post-transplantation lymphoproliferative disorder (PTLD), representing a severe complication of organ transplantation [22]. Several studies have investigated the risk of cervical carcinoma in females positive for HPV after kidney transplantation. Nevertheless, the effect of viral infection or reactivation on acute rejection or decline or loss of function after KTX has not been investigated thoroughly.
Therefore, the development of a diagnostic tool to identify transplant-patient subgroups that benefit from antiviral prophylaxis is essential, along with the investigation of novel tools for the early detection of CMV replication. The extent of CMV replication usually considered as constituting reactivation can be followed with a number of diagnostic methods. However, the detection of de novo CMV infection or reactivation with standard procedures, such as duplex quantitative real-time polymerase chain reaction (PCR) or the COBAS Amplicor CMV Monitor test system (Roche Molecular Diagnostics, Pleasanton, CA), can yield differing results and have a risk of giving false-negative results [23]. In addition, these tests have limited predictive value for identifying patients at risk, and are only partly suitable for early diagnosis. Furthermore, simple diagnostic tools to assess the quality of the cellular antiviral immune response of an individual patient are not routinely established.
Virus-specific T lymphocytes can be enumerated by flow cytometry, allowing rapid and sensitive quantitative measurement of an individual patient's cellular immune response. In clinical settings, major histocompatibility complex (MHC) tetramers need to exhibit robust analytical characteristics and excellent reproducibility. The enumeration of virus-specific CD8+ T-cells using iTAg MHC virus tetramers was shown to have good reproducibility, sensitivity, linearity, and specificity [24–26]. A recent prospective trial of tetramer-based immune monitoring for CMV in stem cell transplant recipients, in conjunction with virologic monitoring, demonstrated this technique as an important new tool that permits clinicians to assess the risk of CMV-related complications and to guide choices for pre-emptive treatment to prevent infection [27].
In the present study we quantified virus-specific CD8+ T-cells to evaluate their potential value for predicting the risk of viral infection in KTX patients. This analysis was not limited to CMV, but also included EBV, HHV, and HPV.
Patients and Methods
Patient population
Twenty-seven patients undergoing KTX were prospectively included in the study between November 2009 and February 2010. Complete HLA typing for all of the patients was available pre-operatively. In case of the risk constellation of donor positivity and recipient negativity for CMV, antiviral prophylaxis was administered. Informed consent was obtained from all patients before their final inclusion, in accord with the requirements of the local aethics committee. Four patients were initially excluded for technical reasons (incomplete pre-operative information, patient's refusal). Because three patients did not have a complete 6-mo follow-up, only their initial data were analyzed and they were excluded from follow-up evaluation. The patients (eight female, 15 male, with an age range of 24–77 y) were investigated pre-operatively (day 0, n=23) and at 2-wk intervals for 2 mo, and subsequently every 4 wk for the first 6 mo after transplantation (n=20). Between follow-up visits, patients were seen by their home-care physicians at least once a week. Patients with symptoms suspicious for disease were immediately referred to the study center. Human leukocyte antigen typing and pre-operative serologic status for CMV were available for all patients and kidney donors. According to a prospective design, blood samples were taken for tetramer analysis, assay or renal function parameters, and blood cell counts at each time point in the study, and patients were examined clinically at these points with special emphasis on detecting signs of viral diseases/exacerbations. Diagnostic tests for viral infection, including polymerase chain reaction (PCR) for CMV, were done only if patients demonstrated apparent viral disease as indicated by clinical signs.
Pre-operatively, tetramer analysis was done with the entire panel of tetramers to enable the measurement of background signals. During follow-up, only those tetramers were used that corresponded with alleles expressed in the typing of kidney recipients' HLA.
Polymerase chain reaction analysis for viral deoxyribonucleic acid
Quantification of viral load was done by PCR with ethylenediamine tetra-acetic acid (EDTA)-treated blood for patients who had any clinical sign of viral disease. For this assay, deoxyribonucleic acid (DNA) was isolated and virus-specific PCR was done through TaqMan analysis (Life Technologies, Grand Island, NY) according to standard procedures.
Class I major histocompatibility complex tetramer analysis
We evaluated the performance of Class I iTAg MHC CMV Tetramers (Beckman Coulter, Krefeld, Germany) for the enumeration of CMV-specific CD8+ T-cells with a three-color, single-platform, two-panel, whole-blood technique. Absolute numbers of CD3+/CD8+ T-cells were enumerated through the use of anti-CD3 and anti-CD8 antibodies and Flow-Count Fluorospheres (Beckman Coulter) in a lyse/no-wash method (Panel 1). Tetramers were labeled with either phycoerythrin (PE) or fluorescein isothiocyanate (FITC). The frequency of tetramer-positive cells was determined with anti-CD3 and anti-CD8 antibodies and tetramers in a lyse-and-wash method (Panel 2 ). Antibodies and tetramers were added to EDTA-blood and incubated for 30 min at 4°C. Lysing buffer was added, and the cells were repeatedly washed with phosphate-buffered saline (PBS) containing 1% fetal calf serum (FCS) and 0.1% sodium azide. For the quantification of tetramer-binding CD8+ T-cells, at least 50,000 living T-cells were counted with a flow cytometer (FC500, Beckman Coulter). The absolute numbers and percentages of circulating tetramer-binding CD8+ T-cells were calculated from the proportion of CD8+ T-cells that bound tetramer and the simultaneously obtained absolute CD8+ T-cell count.
Thirty-six virus specific binding peptides for five different HLA alleles were used for tetramer analysis (Table 1). All antibodies and buffers were obtained from Beckman Coulter Immunomics Operations.
For some virus species only binding sequences for HLA A0201 were sufficiently described.
CMV=cytomegalovirus; EBV=Epstein–Barr virus; HPV=human papillomavirus.
Statistical analysis
Continuous variables are expressed as median and range. The χ2 and Fisher exact tests were used for categorical parameters when appropriate. Kendall tau rank correlation was used to compare the predictive values of the parameters, and Mantel–Haenszel tests and Cochran tests were used for multivariable analysis. Values of p<0.05 were considered significant. All Statistical analyses were done with the SPSS 16.0 software package (SPSS, Chicago, IL).
Results
Virologic status of patients
Pre-operatively, none of the patients showed clinical or serologic signs of acute CMV reactivation (CMV-IgM negative). Eleven recipients and 14 donors were seropositive (CMV-IgG+) prior to KTX (Table 2). All of the patients were negative for acute viral hepatitis. Pre-operatively, none of the patients had a known viral reactivation and all of them were free of clinical signs suggestive of virus-related disease.
CMV=cytomegalovirus; IgG=immunoglobulin G.
Clinical signs of viral reactivation or disease were observed in seven patients during follow-up, including reactivation of CMV or disease caused by CMV (n=3), polyoma virus (BK1, n=3), varicella zoster virus (VZV) (n=2), herpes simplex virus (HSV) (n=2) and EBV (n=1). In two of these patients multiple viral reactivations were found and confirmed by serologic testing. Five of the patients who developed viral disease during the first 6 mo after KTX were CMV-seronegative (CMV-IgG-) pre-operatively, but two of them received organs from seropositive donors (CMV-IgG+). None of the patients died during follow-up, and all of the transplanted organs were functionally active during the observation period.
Virus-specific CD8+ T-cells
In a preliminary study of 87 patients, a cut-off value of 0.3% for tetramer positivity was defined (unpublished results). With this cut-off valuem tetramer binding to virus specific T-cells was detected at least once in our specimens for 32 of the 36 binding sequences. Because monocytes can phagocytose tetramers, cells exhibiting fluorescence can be generated by specific binding to T-cell receptors, but also by the intracellular uptake of tetramers. T-cell specificity was therefore ensured by the exclusion of fluorescence-positive monocytes (Fig. 1).

Cytomegalovirus-specific CD8+ T-cells
Pre-operatively, CMV-specific CD8+ T-cells were found in seven patients. These CMV-specific cells were not found in three CMV-seropositive (CMV-IgG+) kidney recipients prior to KTX, but were detected in three CMV-seronegative (CMV-IgG-) patients (Table 3). This resulted in negative and positive predictive values of 0.81 and 0.57, respectively. Pre-operative serologic CMV status and the percentages of CMV-specific CD8+ T-cells were significantly correlated (p=0.0046). All but one of the patients initially positive for CMV-specific CD8+ T-cells remained tetramer-positive, and in six initially tetramer-negative patients, CMV-specific CD8+ T-cells were found during follow-up. All three patients with clinical CMV reactivation remained or became CMV-tetramer-positive for at least one allele. In addition, four patients in whom CMV-specific CD8+ T-cells were not initially detected became positive for these cells during follow-up. Surprisingly, three of the four patients with non-CMV viral reactivations were initially negative for CMV-specific CD8+ T-cells but became positive for these cells during follow-up. Because of the limited number of patients in this pilot study, the effect of different immunosuppressive agents was not investigated.
CMV=cytomegalovirus; IgG=immunoglobulin G.
The viral load of CMV was quantified with PCR in patients with CMV diseases. The DNA of CMV was detected in all diseased patients, but the numbers of copies did not correlate with thr percentages of CMV-specific CD8+ T-cells.
In contrast, CMV-specific CD8+ T-cells were detected before the clinical appearance of CMV disease in two of the three patients with post-operative CMV disease and in one patient at the time of clinical diagnosis of CMV disease. This T-cell response was specific for certain alleles and peptide sequences (Fig. 2a). After antiviral treatment was begun, the percentage of CMV-specific CD8+ T-cells in these patients declined (Fig. 2b).
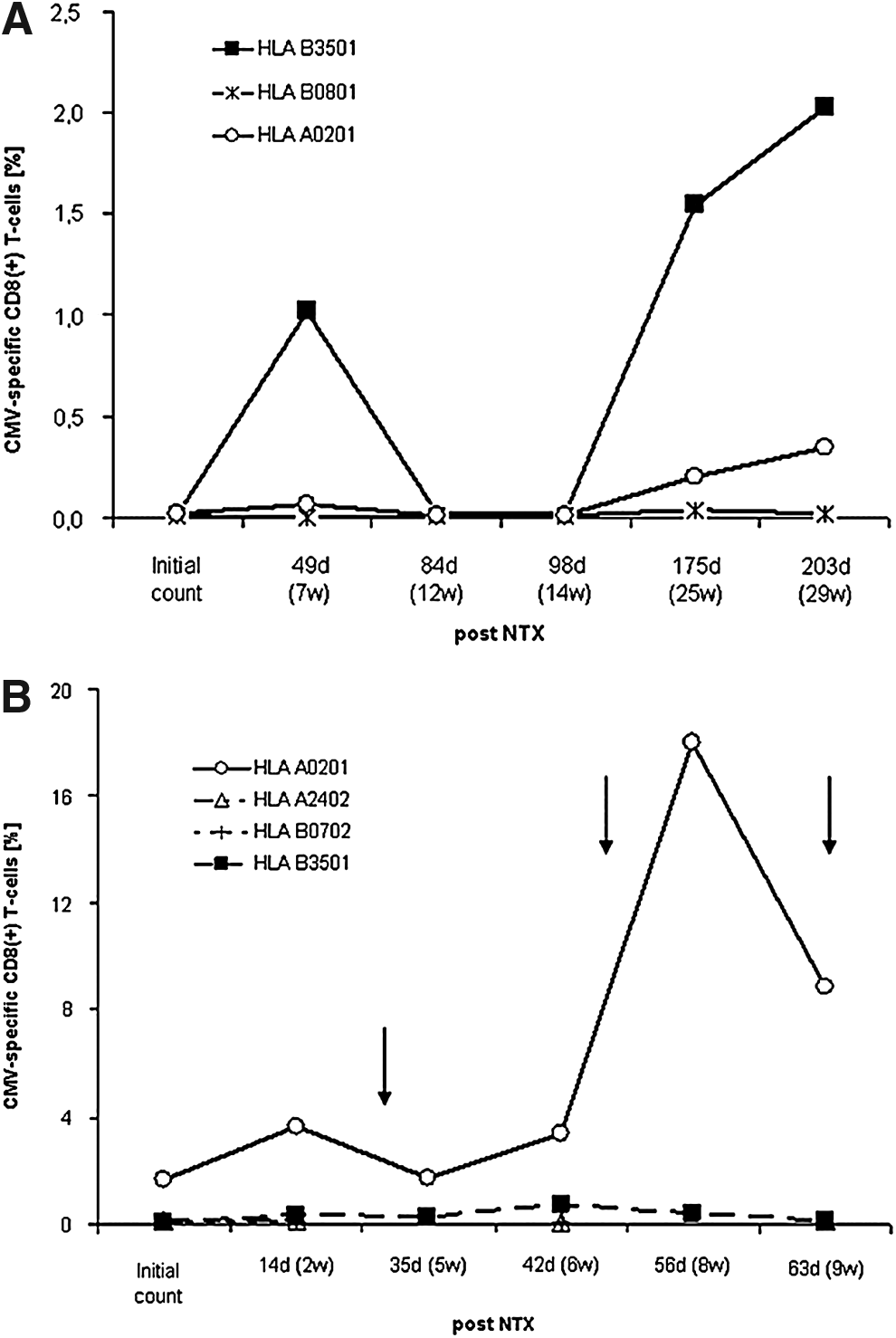
Other virus-specific CD8+ T-cells
Besides CMV-specific tetramer binding, CD8+ T-cell responses specific for other types of viruses were also observed (Fig. 3A). With the exception of one patient, all of the patients who were positive initially for CMV-specific CD8+ T-cells also initially had tetramer-positive cells that bound peptides specific for EBV; HPV-6b, -11, -16, -18; or HHV-8. All four patients who developed non-CMV-induced viral disease remained or became positive for at least one binding peptide. This was even more noticeable in that only a limited number of allele-specific tetramers was available for most of these viruses.
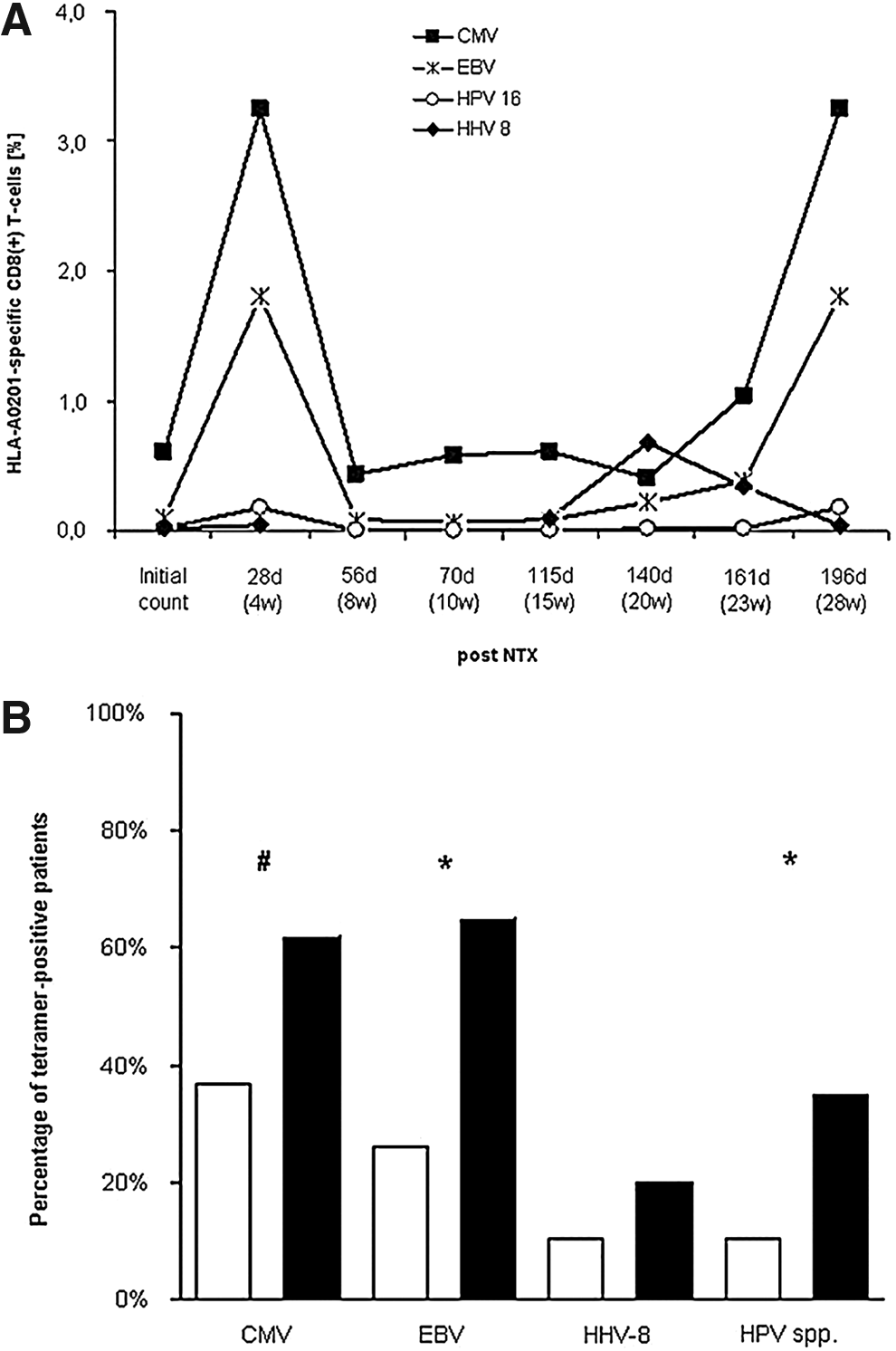
Nine patients who were initially negative for EBV-specific CD8+ T-cells became EBV-tetramer positive. A similar finding was made for four patients for subspecies of HPV and for two patients with HHV. Only one patient who was initially positive for EBV- and HHV-specific CD8+ T-cells turned from became negative for these cells during follow-up, without any clinical signs of viral disease. For all of the types of viruses investigated in the study, a statistically significant increase in the percentage of patients with tetramer-positivity for CMV (p<0.05), EBV (p<0.005), and types of HPV (p<0.005) was observed in the findings of pre- and post-operative analyses (Fig. 3B).
Predictive potential of virus-specific CD8+ T-cell determination
Although the small number of patients in this study limited the application of statistical tests, we proceeded with statistical analyses on the basis that the study data were preliminary. Kendall tau tests showed nearly significant bivariate correlation (p=0.077) in the relationship between pre- and post-operative numbers of CMV-specific CD8+ T-cells. In testing the dichotomous dependence of initial tetramer positivity and post-operative follow-up (Mantel–Haenszel and Cochran tests), we found that the pre-operative absence of CD8+ T-cells specific for EBV (p=0.111) and HPV (p=0.028) appeared to potentially predict an absence of virus-related disease during the first 6 mo after transplantation.
Discussion
Viral diseases occurring after organ transplantation are among the foremost determinants of transplant function, morbidity, and mortality in organ transplant recipients. Immunosuppression and the underlying disease are major risk factors for the reactivation of latent CMV and de novo viral infection. Thus, for example, the inhibition of lymphocyte proliferation by immunosuppressive drugs correlates with the development of active CMV disease in transplant patients, and can be observed in up to 75% of patients depending on the immunosuppressive regimen that is being used [28–33]. These observations have been confirmed with the clinical use of anti-lymphocyte therapy such as anti-thymocyte globulin (ATG) or muromonab-CT3 (OKT3), as well as of pulsed steroids, for the prevention and treatment of graft rejection [34].
Primarily, control of the replication of CMV and other viruses depends on anti-viral T-lymphocyte activity. However, the functional T-cell responses to CMV and other viruses in immunosuppressed solid organ transplant recipients are only partly known. Recent advances in the characterization of T-cell functions and identification of distinct functional signatures of T-cell responses to viruses have opened new perspectives for monitoring transplant recipients at risk for CMV disease [12–17]. In kidney, heart, and lung transplant recipients, CMV disease was found to develop exclusively in patients with a dominant pp65-specific CD8+ T-cell response, and an increased number of IE-1-specific CD8(+) T-cells was correlated inversely with the risk for CMV disease [18,35]. In both healthy controls and KTX recipients, numbers of CMV-specific T-cells were found to correlate strongly with the serostatus of anti-CMV immunoglobulins. This was also found in our study with high specificity but only moderate sensitivity. In addition, Radha et al. [36] reported that seropositive KTX recipients had significantly greater CMV-specific CD8+ T-cell populations than did healthy controls (without KTX and without clinical signs and symptoms of viral diseases).
However, because serologic status and T-cell response reflect different physiologic levels of antiviral activity the determination of sensitivity and specificity for tetramer-positivity remains difficult. The endpoint in further investigation of the usefulness of tetramer diagnostics in KTX patients should be viral reactivation rather than serostatus.
Recently, Lee et al. [37] reported that their patient cohort did not show significant differences in the percentages of pentamer-positive CD8+ T-cells in patients who and those who did not experience episodes of CMV infection, and concluded that the MHC-pentamer-based technique does not seem to provide a clear distinction between patients who are and those who are not at risk for CMV infection.
However, studies have shown that patients with acute CMV disease displayed high initial but rapidly decreasing numbers of CMV-specific T-cells [38], and that the numbers of CMV-specific T-cells decreased rapidly after transplantation [39]. Our data, for a limited number of patients with CMV disease, support these findings and suggest that newly acquired tetramer-positivity or an increase in the percentage of CMV-specific CD8+ T-cells may permit the early diagnosis of CMV reactivations and/or de novo CMV infection. Because all of the patients in our study who developed viral disease in their early follow-up were or became positive for virus specific CD8+ T-cells, prospective determination of their responses may contribute to the early identification of patients at risk for transplantation-associated CMV disease. Because we also found this increase in tetramer conversion/tetramer positivity for all of the other viruses investigated in our study, tetramer-based diagnosis can probably identify a number of other viral diseases, if not all viral diseases, that can follow solid-organ transplantation.
This novel diagnostic strategy might be combined with other new developments related to the tetramer-based identification of virus-specific T-cell responses. For the first time we observed in our study that anti-viral T-cell responses appear to not be limited to a single type of virus, but can encompass an enhanced response to various types of viruses, such as EBV, HPV species, and HHV. Although tetramers were available for only one allele, respectively, in the case of HPV and HHV-8, and not at all for BK1 polyoma virus, we identified a high frequency of multiple anti-viral T-cell responses that probably reflect physiologic efforts to control enhanced viral replication in immunosuppressed KTX recipients. The delay between virus reactivation and the expansion of a virus-specific T-cell population to the point of detectability is probably influenced by the different expression kinetics of various viral antigens according to the intermediate early, early, and late responses to them [35].
Viral genomes contain large numbers of potentially immunogeneic epitopes with different involvement in the development of viral disease and allograft rejection. Screening for larger numbers of these epitopes in patients with rejection and CMV reactivation would shed more light on the T-cell responses induced by viruses.
As shown in a recent pilot study, CMV-specific tetramer-positive CD8+ and IFN-γ-positive CD8+ T-cells regenerated within 3 mo after transplantation, whereas IFN-γ-positive CD4+ cell recovery was impaired for up to one year after transplantation. The proportion of IFN-γ-positive CD4+ cells at 2 mo after transplantation as compared with the baseline proportion of such cells correlated strongly with the concentration of CMV DNA in the blood [39].
In a recent clinical trial of antiviral prophylaxis, PCR-based quantification of the copy number of CMV DNA was proposed as a marker for determining the duration of prophylaxis in high-risk patients after KTX. However, a high individual variability in the predictive value of this putative marker for subsequent CMV disease was applicable only after logarithmic adaptation. Therefore, in the developed Markov model, this parameter achieved only minor importance. Instead, because the number of virus-specific CD8+ T-cells reflects functional antiviral host responses, the enumeration of these cells at certain post-operative time points could add predictive power [40]. Furthermore, monitoring of virus-specific CD8+ T-cells and the reduction in number of CD8+ T-cells that are producing interferon (IFN)-γ and their CD4+ counterparts could be considered in predicting the risk for a high concentration of CMV DNA in the blood [39]. Although it has so far been investigated only in small preliminary studies, such a combined approach might assist in strategic decisions about pre-emptive and/or prophylactic therapy against CMV infection [40].
A lack of tetramer staining has been observed despite properly used methodology and wikth T-cells carrying the appropriate epitopes. Therefore, in class I or II MHC tetramer staining assays in which mixed populations of T-cells are analyzed, it is possible that not all of the relevant antigen-specific T-cells will be detected. The fraction of the total T-cell population represented by these non-staining cells, and how they differ from the antigen-specific cells that do stain, is not currently known [41].
The clinical importance of early virus detection in transplant patients exceeds the management of acute infectious problems. Reactivations of HPV are considered as causative or at least as a co-factor in the development of various types of cancers [42], many of which can be found at increased incidences in solid-organ transplant recipients [43–45]. Similarly, reactivations of EBV appear to be a relevant factor in the high incidence of lymphoproliferative disorders in transplant patients [46,47], making continuous screening of EBV DNA necessary in these patients. Our new observation that KTX patients frequently have multiple anti-viral CD8+ T-cell responses should be investigated for its relationship to immunocompetence, immunosuppression, and long-term outcome, to shed light on the underlying pathomechanisms of the diseases caused by the respective viruses.
The major limitations of our pilot studies are the small number of patients they involve and the consequently limited possibility of evaluating the specificity and sensitivity of the virus-specific CD8+ T-cell counts found in these patients, along with the predictive values of these counts. In addition, HLA A0201 is not expressed in the usual Caucasian population in central Europe, and a number of virus-specific T-cell responses to types of HPV and HHV may therefore not be detected in MHC-teramer-based assays. Furthermore, virus families other than those of the species investigated in the present study may be important in terms of long-term morbidity in KTX patients.
The monitoring of cellular immunity through the assay of anti-viral CD8+ T-cell responses, and the combination of this information with the determination of viral load by PCR and the counting of interferon-responsive T-cells, merits further exploration [41]. Ultimately, this approach may allow the identification of patients at risk for CMV or other viral diseases, the tailoring of prophylactic and therapeutic decisions to meet specific needs, and the prevention of viral complications of organ transplantation.
Footnotes
Acknowledgments
This study was supported by an unrestricted grant from Roche Pharma AG. The authors thank Mrs. H. Lehmann and Mrs. F. Spiecker for their strong support of the study. Soeren Torge Mees, Linus Kebschull, and Wolf Arif Mardin collected the data for this paper. Soeren Torge Mees and Wolf Arif Mardin analyzed the data for the paper, and Soeren Torge Mees drafted the manuscript of the paper. Norbert Senninger, Barbara Suwelack, and Heiner Wolters revised the manuscript, and Joerg Haier provided the concept and design for the paper and was responsible for the statistical analyses.
Author Disclosure Statement
No competing financial interests exist.