
Abstract
Background:
Sepsis is defined as a dysregulated host response to infection, resulting in life-threatening organ dysfunction. It is now understood that this dysregulation not only constitutes excessive inflammation, but also sustained immune suppression. Immune-modulatory therapies thus have great potential for novel sepsis therapies. Here, we provide a review of biomarkers and functional assays designed to immunologically stage patients with sepsis as well as therapies designed to alter the innate and adaptive immune systems of patients with sepsis beneficially.
Methods:
A search of PubMed/MEDLINE and clinicaltrials.gov was performed between October 1, 2019 and December 22, 2019 using search terms such as “sepsis immunotherapy,” “sepsis biomarkers,” “sepsis clinical trials,” and variations thereof.
Results:
Despite more than 30 years of research, there is still no Food and Drug Administration (FDA)-cleared biomarker that has proven to be effective in either identifying patients with sepsis who are at an increased risk of adverse outcomes or responsive to specific interventions. Similarly, past clinical trials investigating new treatment strategies have rarely stratified patients with sepsis. Overall, the results of these trials have been disappointing. Novel efforts to properly gauge an individual patient's immune response and choose an appropriate immunomodulatory agent based on the results are underway.
Conclusion:
Our evolving understanding of the different mechanisms perturbing immune homeostasis during sepsis strongly suggests that future successes will depend on finding the right therapy for the right patient and administering it at the right time. For such a personalized medicine approach, novel biomarkers and functional assays to properly stage the patient with sepsis will be crucial. The growing repertoire of immunomodulatory agents at our disposal, as well as re-appraisal of agents that have already been tested in unstratified cohorts of patients with sepsis, may finally translate into successful treatment strategies for sepsis.
The definition of sepsis has evolved along with our improved understanding of the disease. Initially thought of only as severe infection and systemic inflammation, sepsis has been re-defined as a life-threatening organ dysfunction caused by a dysregulated host response to infection [1]. This definition accounts for the complex changes to the immune system during sepsis, because it is now understood that sepsis induces a fundamental re-organization of immune and metabolic processes, in which both pro- and anti-inflammatory mechanisms contribute to the clearance of infection and tissue recovery, as well as organ injury and secondary infections. The mechanisms known to be involved so far have been reviewed extensively elsewhere [2,3] (Fig. 1). Thus, it is crucial to develop therapies for sepsis that aim not only at eradication of pathogens and depression of any overwhelming inflammatory responses, but also the restoration of homeostasis and proper immune function (Fig. 2).

Hallmarks of sepsis. Sepsis is defined as life-threatening organ dysfunction caused by a dysregulated host response to infection. This involves a fundamental reorganization of immune and metabolic processes, in which both pro- and anti-inflammatory mechanisms contribute to the clearance of infection and tissue recovery, as well as abnormal coagulation, organ injury and secondary infection.
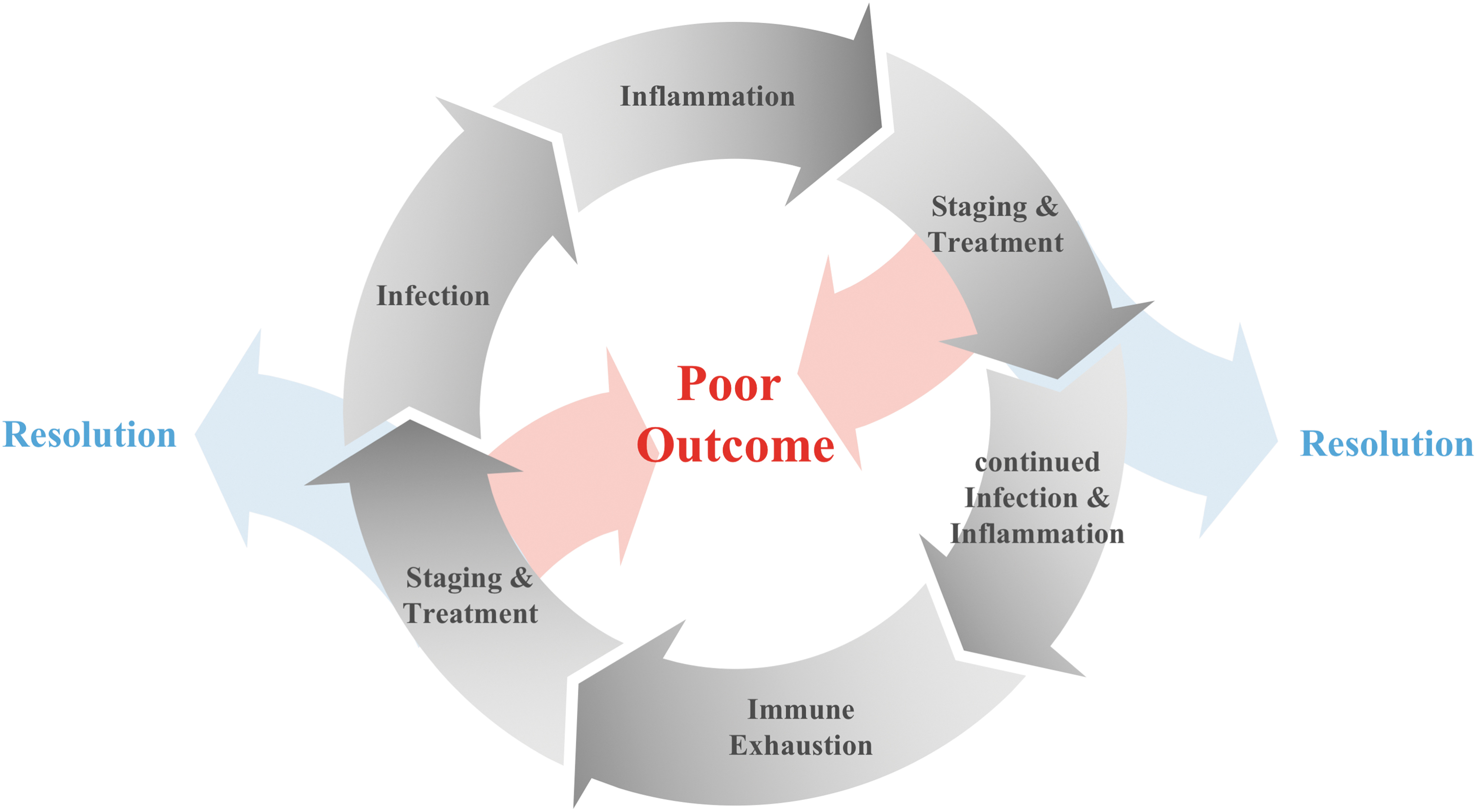
The sepsis cycle. The initial infection results in an inflammatory response, which, in sepsis, can be excessive. Failure to clear the infection and continued inflammation can result in immune exhaustion, making patients more susceptible to secondary infections. These, in turn, will further drive the vicious cycle of inflammation and immune exhaustion leading to poor outcomes, i.e., death or chronic critical illness. Immune-phenotyping patients and choosing therapies specifically for the respective immune status of an individual patients offers the chance to break the wheel: tempering the hyper-inflammatory response prevents inflammation-related organ dysfunction and enables successful resolution of infection and inflammation before the immune system can become exhausted. Immune-augmentation strategies, on the other hand, will re-activate the suppressed immune response and thus enable clearance of infection and resolution of inflammation in patients with immune-exhaustion.
Currently, we have access to a growing repertoire of immune modulating drugs. Many have been developed as anti-neoplastic agents, because tumor growth and progression depend on successful immune evasion as well as cancer-mediated inflammation. These similarities in pro- and anti-inflammatory changes happening in parallel indicate that immune-modulating therapies developed originally for patients with cancer may also be promising new therapies for patients with sepsis. One example of an available immune-modulator that has already been tested for its suitability for patients with sepsis is a monoclonal antibody (mAb) against tumor necrosis factor-α (TNF-α). Unfortunately, the first trial failed to demonstrate a survival benefit and was terminated early [4]. A later analysis, however, showed that in a subgroup of patients with shock, TNF-α mAb treatment actually reversed septic shock and delayed the onset of organ failure [5]. Similarly, a study using anakinra (recombinant human interleukin-1 receptor antagonist) initially failed to demonstrate a reduction in mortality and was also terminated early [6]. However, a post hoc analysis later demonstrated that a subgroup of patients with macrophage activation syndrome had had improved 28-day survival and had actually benefited from anakinra [7]. However, these promising findings were lost in the overall analysis of patients, because of their varying immune status at the time of agent administration. This highlights the importance of properly staging the immune status of the specific patient with sepsis for therapeutic success.
As these two examples show, failure of past clinical trials in sepsis is not necessarily because of an ineffectiveness of the tested drugs per se, but rather caused by the lack of restricting treatment to selected groups of patients who may actually benefit from a specific type of therapy [2,8–10]. Therefore, it is generally agreed that accurate and reliable immune-monitoring is imperative for the success of future trials [2,8,10,11].
The first part of this review addresses potential new biomarkers and immunologic methods that may be incorporated into our clinical routine to make precision medicine possible in sepsis. The second part addresses novel interventions under consideration for sepsis therapy.
Sepsis Biomarkers and Patient Stratification
Current sepsis biomarkers
Prior to the current Sepsis-3 definition, sepsis was defined as a confirmed or suspected infection in addition to two or more systemic inflammatory response syndrome (SIRS) criteria, which include leukocytosis or leukopenia and fever or hypothermia. Leukocyte counts could thus be considered one of the earliest biomarkers for sepsis. However, none of the SIRS criteria are sensitive or specific enough to diagnose sepsis per se. On the other hand, persistent lymphopenia has been associated with increased risk of developing secondary infections and predicts early and late mortality [16].
To date, biomarkers in sepsis are static measurements, i.e., they provide a snapshot of the concentration or level of the respective analyte at the time of analysis. For a while, C-reactive protein was used commonly as a biomarker for infection in sepsis. C-reactive protein (CRP) is an acute-phase reactant secreted by the liver, which increases markedly in response to infection. Measurements are routine and rapidly available, which can aid in the early diagnosis of sepsis, particularly because laboratory confirmation of infection by microbiologic means can be inconsistent. Additionally, monitoring CRP levels after intervention can predict an individual's clinical course [17]. However, CRP has since fallen out of favor, because its specificity as a sepsis biomarker has been reported to be as low as 40% despite the use of higher than recommended cutoff values, causing concern that many patients with non-infectious inflammatory conditions would be misidentified as having sepsis based on CRP values alone [18,19]. As an alternative, procalcitonin (PCT), the prohormone of calcitonin, has been suggested to gauge accurately the onset and magnitude of the hyperinflammatory response. High PCT levels have also been shown to predict progression to septic shock, and PCT clearance has been correlated with survival in some studies [20–22]. However, among the 17 trials assessing the prognostic value of PCT concentrations, five also yielded negative or equivocal results [20–22]. Another soluble molecule under consideration is a monocyte activation marker, soluble receptor for advance glycation end-products (sRAGE), which has been associated with poor survival in sepsis [23]. Further work is needed on these markers to establish their usefulness beyond the diagnosis of sepsis, and in guiding immune-therapy targeting of excessive inflammation.
Immunologic staging of sepsis
Requirements for an immune status panel
Previous studies have focused only on inflammatory mediators as biomarkers in an effort to direct early therapeutic interventions toward suppression of the cytokine storm [13]. In that regard, the previously mentioned example of anakinra demonstrates how macrophage activation syndrome—a fulminant cytokine storm with simultaneous hepatosplenomegaly, liver dysfunction, hyperferritemia, pancytopenia, and disseminated intravascular coagulation [14]—may serve as a stratification criterion to identify patients with a severe hyperinflammatory state. However, to administer the appropriate immunomodulatory regimen for each patient at the right time, it is not just the patient's inflammatory response that needs to be considered, but also the degree of immunosuppression. Based on the complexity of the septic response, it is clear that a single biomarker will not have the necessary sensitivity to achieve both. The combination of biomarkers coupled with functional assays, however, will likely improve the accuracy of sepsis staging [12]. In addition to accurately differentiating the varying levels of hyperinflammation and immunodeficiency, such an immune status panel would also need to be available in a timely manner, enabling clinicians to make an informed decision at the time of care. Ideally, the panel would also be reproducible over time to allow for monitoring of disease progression and therapy response, as well as to guide the duration and intensity of drug application as well as the decision to switch to a different regimen [15]. Biomarkers could also help to identify patients with a high risk of adverse outcomes, i.e., secondary infection, which could prompt closer monitoring and implementation of more stringent protective measures for these patients. In the next section, we will review biomarkers and functional assays that may come to comprise such an “immune status” panel for sepsis.
Potential biomarkers and functional assays for immunologic staging of sepsis
Cytokines are promising biomarkers for the immune staging of sepsis, because results can be obtained within hours by enzyme-linked immunosorbent assay (ELISA), which can be easily standardized and even automated for widespread clinical use [24]. TNF-α, interleukin (IL)-6, and IL-1β are the predominant cytokine mediators of the hyperinflammatory response in sepsis, whereas IL-10 is the major opposing player for the compensatory anti-inflammatory response. Correlations between the levels of these cytokines, disease severity, and outcomes in sepsis have been described [25–27]. Mechanistically, cytokine measurements can be repeated easily to monitor disease progression. However, the success of this strategy may be limited, given that the cytokines currently being considered are released non-specifically in other inflammatory conditions as well and may not remain elevated long enough to be useful [15,19].
Transcriptomic or epigenetic changes may prove to be useful biomarkers. Although the relatively long turnaround time for obtaining the data limits clinical usefulness so far, promising results linking whole-blood mRNA levels of CD74, human leukocyte antigen-DR (HLA-DR), and IL-10 were able to predict and risk-stratify patients at risk for intensive care unit (ICU)-acquired infections [28]. Moreover, a study using global genomic profiling of circulating leukocytes in patients with pneumonia identified a subset of patients with relative immune-suppression, endotoxin tolerance, and T-cell exhaustion [29]. Similar results were identified in a group of patients with peritonitis [30]. Another study in patients with sepsis also reports the identification of a transcriptome profile associated with improvement of organ function after early supportive therapy. Unfortunately, however, the study could not yet distinguish a profile based on gene expression immediately upon ICU admission [31].
As mentioned previously, current biomarkers for sepsis are mostly static measurements. This is a limitation, because the functional status of a patient's immune cells is not being assessed routinely. Although still a static measurement, flow cytometric measurements can be used to gain at least some preliminary insights into a patient's functional status. Flow cytometry enables the characterization and enumeration of leukocyte phenotypes and activation status in sepsis. Lymphopenia, including reduction of CD4+ and CD8+ T cells and B cells, can be seen at the onset of sepsis and can persist for up to 38 days [32]. It has been shown to predict mortality and is associated with increased infection susceptibility [16], suggesting that lymphopenia may serve as a marker and a target for immune-augmentation therapy. Specifically, regarding T cells, the absence of activation marker expression (i.e., CD25, CD44, CD69 and CD71) but presence of CD62L indicates T cell inactivation and potentially a shift toward a helper T cell 2 (TH2) profile and apoptosis [33,34]. This resulting apoptosis drives a contraction of the T-cell population and T-cell receptor repertoire diversity, which has been observed using flow cytometry [35,36]. Decreased T-cell receptor diversity and expansion of regulatory T cells, which can mediate T-cell anergy in patients with sepsis, are associated with an increased risk for infection and death in sepsis [37,38]. Expression of immune checkpoint receptors on T cells (i.e., PD-1, CTLA-4) and expression of their ligands on myeloid cells (i.e., PD-L1, PD-L2, CD80, CD86) may also be useful biomarkers for sepsis-induced immunosuppression [39–42], in addition to being investigated as therapeutic targets for sepsis. In neutrophils, low expression of CD88 (C5a receptor) indicates suppression of myeloid cells [43,44]. In addition, reduced levels of HLA-DR have been implicated during immune-paralysis and have been associated with the development of nosocomial secondary infections [47–49]. On the other hand, expression of CD64 increases in parallel to the degree of the inflammatory response [48]. Despite concerns that CD64 sensitivity may be low, CD64 has been demonstrated to be a more sensitive predictor of sepsis than HLA-DR in the pediatric population [49]. Human leukocyte antigen-DR expression is already being used for patient stratification in ongoing sepsis clinical trials (i.e., NCT0261528).
Although such flow cytometric measurements certainly offer important insights, they still do not assess adequately the functionality of patients with sepsis immune cells. Munoz and Cavaillon [50] first demonstrated that, despite elevated plasma concentrations of TNF-α, IL-1β, and IL-6 in patients with sepsis, the ex vivo production of TNF-α and IL-1β by blood monocytes in response to endotoxin was reduced dramatically. Similar findings have since been reported multiple times, not just for blood monocytes but also for studies involving whole blood from adults with sepsis, as well as experimental models of sepsis [51,52]. Importantly, the ex vivo response of whole-blood stimulation in pediatric patients with sepsis has been associated with both multiorgan failure and death, with receiver-operator curves predicting survival (0.71–0.75) [53,54]. Thus, while ex vivo approaches may take longer than measuring serum cytokine levels or other static parameters directly, they provide important functional insight. It is promising that clinical studies aiming to reverse sepsis-induced immunosuppression with granulocyte-macrophage colony stimulating factor (GM-CSF) and interferon-γ (IFN-γ) have started to assess monocyte responsiveness upon ex vivo lipopolysaccharide (LPS) challenge to guide and monitor therapy [55,56]. Additionally, such ex vivo analyses offer a unique opportunity to test the responsiveness of cells from individual patients to the agent being considered for their personalized treatment.
Another promising option to functionally determine the individual's immune status is enzyme-linked immune absorbent spot assays (ELISPOT). This method can gauge how many cells respond to the stimulation and/or intervention. This assay also requires only a low number of cells, so that multiple analyses can typically be conducted with one sample for both pediatric and adult critically ill patients.
Although further validation, standardization, and ultimately automation will be necessary before the aforementioned assays will find widespread clinical application [57], this is likely only a short-term limitation. For example, ELISPOTs specifically are technically easy to perform and standardize and the ELISPOT technology is already cleared by the Food and Drug Administration (FDA). The ELISPOT assays have excellent dynamic range (among 100,000 cells, as little as a single cytokine producing cell can be identified and as many as several thousand), and can gauge both the number of cytokine-secreting cells and the relative amount produced [58]. Because ELISPOTs quantitate the total amount of a cell's cytokine production over an extended period (≤24 h), they are more sensitive at revealing group differences in cell function compared with other methods using shorter incubation times. Additionally, ELISPOTs can assess independently the functional status of both adaptive and innate immunity by assessing CD3/CD28-stimulated IFN-γ production and LPS-stimulated TNF-α production.
A potential limitation of all functional assays is the compartmentalization of the immune response. Circulating leukocytes, although easy to obtain, may not exhibit the same functionality and responsiveness as leukocytes obtained from the site of infection or other non-blood compartments [19]. The latter, however, may be more difficult or even impossible to isolate without invasive methods, and thus may not be available for testing, much less repeated testing during the clinical course.
Speculative model of immune phenotypes in sepsis
Despite the growing list of sepsis biomarkers and developing efforts to assess immunologic function in patients with sepsis, these advances have not been adapted readily to biomarker discovery and clinical development as yet. For the fastest clinical implementation, the best options are measurements obtained with instruments that are already FDA-approved. For the markers and measurements discussed here, this means blood sampling for functional ELISPOT assays along with complete blood counts with differentials. We speculate that immune phenotyping based on a combination of cell counts and functional ELISPOT assessments will distinguish four innate response categories and four separate adaptive response categories (Fig. 3a and 3b).
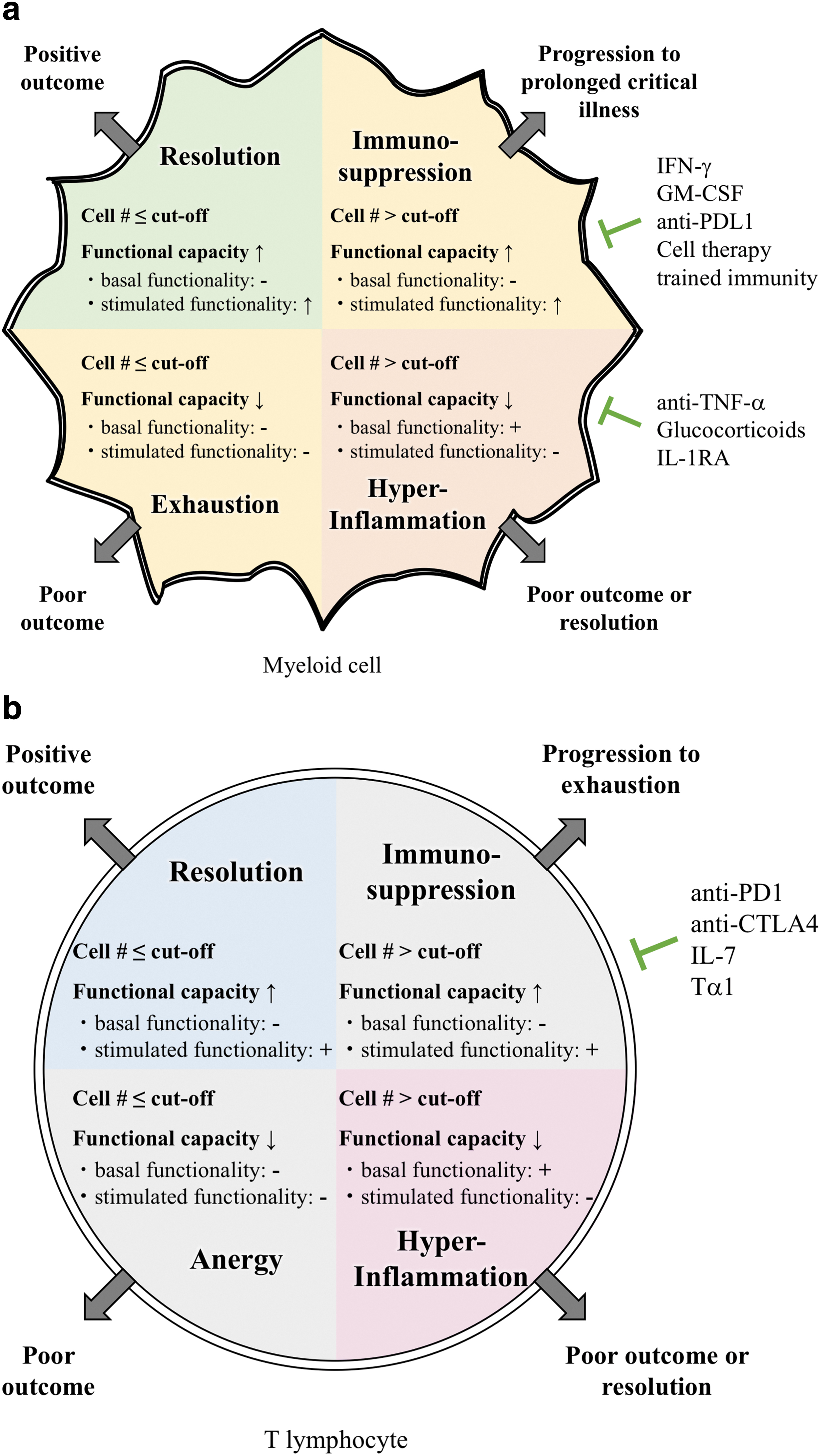
Predicted immune phenotypes in sepsis and their treatment needs. Immune-phenotyping in sepsis based on cell counts and ex vivo functional assessments of a patient's cells is predicted to assign a patient into one of four groups based on their innate response and functionality
The patients most likely to have a positive outcome are patients whose cell numbers are at or below the cutoff (or average) for the respective cell population and whose cells retain functional capacity (left upper quadrant of Fig. 3a and 3b). In other words, whereas there are little or no basal functionality observed, ex vivo stimulation results in an increased response, so the fold change comparing baseline to stimulation is high. The fact that there is little basal activity in combination with normal/low cell numbers in these patients reflects that they have resolved the infection successfully (or were not infected).
In the second group of patients, the cell numbers will currently be at or rise above the cutoff level (right upper quadrant of Fig. 3a and 3b). The lack of basal activity in these patients' cells would indicate that they are not responding adequately to the infection in vivo. However, the functional capacity of these cells may still seem intact because they respond to stimulation. Thus, these patients are immunosuppressed in vivo and would potentially benefit from immune-augmenting therapies.
A subset of immunosuppressed patients may progress to a third stage, exhaustion or anergy, in which their cell numbers drop below the cutoff and the remaining cells lose their ability to respond to stimulation (left lower quadrant of Fig. 3a and 3b). It is unclear if these patients would still benefit from immune-augmenting therapies and their outcome without intervention is likely poor.
Finally, a fourth group of patients can progress to or present as being in a hyperinflammatory state, indicated by high cell numbers and high basal functionality, which cannot be enhanced further with stimulation (right lower quadrant of Fig. 3a and 3b). These patients may be ideal candidates for agents that temper the inflammatory response.
Distinguishing the innate and adaptive responses into each of these categories is thus predicted to differentiate 16 distinct patient populations. This approach likely only begins to model the true complexity of the septic response. It is important to note again that each patient population in this prediction has specific treatment needs. The next part of this review focuses on novel treatment strategies for sepsis and which subset of patients may benefit from them.
Novel Treatment Strategies for Sepsis
Treatments targeting hyperinflammation
Previous therapeutic interventions have almost exclusively been immunosuppressive in nature. Despite the failure of clinical trials thus far, the previously discussed retrospective analyses of TNF-α, mAb, and anakinra indicate that these interventions may still prove useful for sepsis patients that are indeed in a hyperinflammatory state. In the anakinra trial, at least 6% of patients suffered from macrophage activation syndrome. The actual percentage may be as high as 25%, the proportion of patients with sepsis who die within the first four days of admission, likely the result of an excessive inflammatory response [11]. Other clinical consequences of hyperinflammation may help to identify patients who may benefit from such therapies, including high fever, septic shock, disseminated intravascular coagulation (DIC), acute respiratory distress syndrome (ARDS), and fulminant liver failure [19].
Glucocorticoids
Glucocorticoids are natural steroid hormones with pleotropic effects, including the upregulation of anti-inflammatory protein expression and downregulation of proinflammatory protein expression through binding of the transcription factor glucocorticoid receptor (GR). Since the first trial conducted in the 1960s, 24 randomized placebo-controlled trials have been published with conflicting results [59–63]. Factors affecting the response of a patient with sepsis to glucocorticoid treatment may have accounted for the conflicting results of the trials, including the time to treatment, dose and dosing strategy, type of glucocorticoid, other drugs administered, as well as patient comorbidities and genetic polymorphisms [64,65]. The timing of treatment initiation is likely a particularly important point. In animal models of SIRS, glucocorticoids were only effective when given at the time of, or immediately after, a TNF challenge. Administration six to eight hours later showed no protective effects. Thus, the anti-inflammatory qualities of glucocorticoids are likely most beneficial in the acute pro-inflammatory state of sepsis. Once the compensatory anti-inflammatory pathways of the host gain traction, the immunosuppressive effects of glucocorticoids are redundant or may even contribute to immune paralysis.
Anti-toxin therapies
Endotoxin is a lipopolysaccharide component of the outer membrane of gram-negative bacteria. It is a potent catalyst of the acute inflammatory response and has thus long been considered a potential therapeutic target in sepsis [66]. Past clinical trials involved antiserum raised against a heat-killed Escherichia coli mutant, humanized antibodies against the active lipid-A component of endotoxin (HA-1A), an antibody directed against a common enterobacterial antigen (T88), and the neutralizing agents reconstituted high-density lipoprotein (HDL), and bactericidal-permeability increasing protein (BPI) [67]. Two studies reported a reduction in mortality [68,69], but no other trials have shown a similar beneficial effect in patients with sepsis [61,67,70–74]. Extracorporeal removal of circulating endotoxin from blood is another strategy that has been discussed since the 1970s [75]. One example is Toraymyxin (Toray Medical Co., Ltd., Tokyo, Japan), a hemoperfusion column composed of polymyxin B fibers, an antibiotic that binds and inactivates endotoxin. Polymyxin B is not suitable for direct in vivo use because of nephrotoxicity and neurotoxicity. Toraymyxin therapy has been associated with reduced mortality and improvements in hemodynamic condition and pulmonary function in some but not across all studies [76].
An overall limitation of these anti-endotoxin therapies is the administration timeframe required for utility. They must be used early in the course of sepsis to derive full potential, because the response to LPS is triggered so rapidly. Additionally, use is limited to patients with gram-negative infections. However, sepsis can also develop from infection with gram-positive bacteria, i.e., Staphylococcus aureus and Streptococcus pneumoniae. These gram-positive bacteria release cytotoxic, pore-forming exotoxins, causing substantial tissue damage. AB103, a peptide antagonist that inhibits the interaction of bacterial super-antigens with T-cell receptor CD28, reduced mortality in mouse models and is currently being tested in a phase 3 clinical trial [77–79]. An extracorporeal blood-cleansing approach that can remove gram-positive as well as gram-negative bacteria, fungi, and endotoxins is also currently being evaluated in preclinical studies [80]. In a new approach, artificially engineered liposomes that compete with the host cells for toxin binding were shown to rescue mice from Staphylococcus aureus- and Staphylococcus pneumoniae-induced septicemia [81]. Potentially, liposomes could also be engineered to sequester toxins secreted by gram-negative pathogens. In the future, early administration of engineered liposomes may be a promising adjunct therapy for patients with sepsis to prevent tissue injury from bacterial toxins.
Therapies targeting specific pro-inflammatory mediators
Tumor necrosis factor-α is released by monocytes and lymphocytes and is a crucial mediator of the innate response. It is one of the first cytokines to appear in circulation in experimental models of sepsis and plays a key role in the development of multiple organ failure and death [82]. Elevated levels of TNF are detected in blood samples of patients with sepsis, with even higher levels in patients who died compared with sepsis survivors [83]. The use of recombinant, soluble TNF receptors to compete for TNF-α binding with the endogenous host receptors has not been successful [84]. Although meta-analyses of trials using anti-TNF-α antibodies do suggest a survival benefit [85], particularly in the patient population suffering from septic shock [5].
Interluekin-1 is another proinflammatory cytokine released early in the course of sepsis. Similar to soluble TNF-α receptors, IL-1 receptor antagonists compete with the cytokine for binding to its endogenous receptor, thus competitively inhibiting the signal. As mentioned, initial studies failed to demonstrate a survival benefit in sepsis [6], but a post hoc analysis later demonstrated improved 28-day survival in a subgroup of patients with macrophage activation syndrome [7].
Although they are not classic immune cells, vascular endothelial cells are also potential targets for new sepsis interventions. Engagement of innate immune receptors including Toll-like receptors, expressed on endothelial cells leads to activation of inflammatory pathways, modulation of endothelial cell permeability and coagulation. Dysregulation of these pathways during sepsis leads to coagulopathy and increased vascular permeability, which contributes substantially to organ failure [86]. Several new treatment strategies aiming to restore endothelial barrier function are in development, including angiopoietin 1 and/or TIE2 modulators, S1P1 agonists, fibrinopeptide Bb15-42 and RAR1 pepducin [87]. Our own preclinical work indicates that amitriptyline, which has already been clinically used as an antidepressant for decades, may be useful in mitigating the initial hypercoagulable state in sepsis and protect from delayed hypocoagulability [88].
Therapies commonly used in the ICU, such as vasopressors for hemodynamic support in septic shock, may have important immune-modulating effects as well. In vitro studies indicate that norepinephrine inhibits macrophage migration and alters cytokine release [89–-92], and in vitro studies and animal models show that vasopressin suppresses pro-inflammatory cytokine release [93,94]. Use of these drugs early in the course of sepsis, when excess inflammation still dominates, may thus have beneficial effects on immune function during sepsis [15].
Immune-augmentation therapies
Immune checkpoint inhibitors
Given the recognition that immune deficits in cancer and sepsis have some overlap has led to the appraisal of existing cancer immunotherapies in sepsis trials. A prominent example is nivolumab, an mAb targeting the PD1/PDL1 immune checkpoint. This major T-cell inhibitory pathway leads to T-cell anergy and apoptosis, IL-10 generation, and myeloid cell dysfunction [95]. Elevated PD1 expression has been reported in patients with sepsis, with a direct correlation to serum IL-10 levels, T-cell apoptosis, nosocomial infection rates, and overall mortality [41,96,97]. In vitro blockade of PD1 reversed T cell apoptosis, decreased serum IL-10 levels and increased pro-inflammatory cytokine levels [96,97].
Other potential immune checkpoint inhibitors for the treatment of sepsis are anti-cytotoxic T-lymphocyte-associated protein 4 antibodies. Anti-cytotoxic T-lymphocyte-associated protein 4 (CTLA4) is expressed on activated T cells and normally binds the costimulatory receptor CD28. However, it has greater affinity for CD80 and CD86, which are expressed on antigen-presenting cells and outcompete CD28 for CTLA4 binding, resulting in the transmission of an inhibitory, rather than a stimulatory signal. When CTLA4 is inhibited, T-cell function is recovered [97]. It has been documented that some patients with sepsis have elevated CTLA4 expression [42,98]. In animal models of sepsis, anti-CTLA4 antibody treatment decreased T cell apoptosis rates and increased survival [99] and even better success was achieved upon combination with anti-PD1 and anti-PDL1 checkpoint inhibitors [52].
It will be important to limit the use of these agents to patients with sepsis who are actually in a T-cell suppressed state, but even then, their use in critical illness may be limited. Cancer studies indicate that the rate of serious adverse effects can be as high as 24% [15]. However, a phase 1 clinical trial recently concluded and indicates that nivolumab had no unexpected safety issues in the treatment of sepsis and septic shock [100].
Recombinant cytokines
Analogous to anti-cytokine therapies targeting excessive inflammation in sepsis, recombinant cytokine therapies can be used to augment the immune response during sepsis-induced immune paralysis. Interferon-γ and GM-CSF are both potent stimulators of innate immunity that have been used in sepsis trials. IFN-γ has been shown to reverse monocyte deactivation and endotoxin tolerance [56,101,102] and has been used safely and effectively in other immune-compromised patient groups (i.e., in human immunodeficiency virus-positive patients and after renal transplant) [103–105]. A phase 3 study on IFN-γ as an adjunctive therapy in patients with septic shock has completed but results are not yet available (NCT01649921).
Granulocyte-macrophage colony stimulating factor, and the related cytokine G-CSF, have been studied in 12 randomized control trials. A meta-analysis concluded that these trials showed improved infection clearance, but the survival benefit is still unclear, because the studies were underpowered for this outcome measure [106,107]. It is important to note that only one of these studies was biomarker-guided, using HLA-DR expression to determine the intensity of GM-CSF administration. Proper stratification of patients is particularly important for GM-CSF, because it also exerts significant anti-inflammatory properties, including expansion of regulatory T cells [108,109]. Administration at the improper stage of sepsis progression therefore carries the risk of exacerbating sepsis-induced immunosuppression. A phase 3 clinical trial to assess the efficacy of GM-CSF in preventing ICU-acquired infections in immunocompromised patients with sepsis has completed recruitment (NCT0261528).
Interleukin-7 has diverse effects on both the innate and adaptive immune system. Recombinant interleukin-7 (rIL-7) successfully restored immune function in patients with human immunodeficiency virus and cancer [110–112], while also being well tolerated [113–115]. Interleukin-7 levels in patients with sepsis have been reported to be lower than in controls [116], but there are some discrepant results regarding possible differences in IL-7 levels during different stages of sepsis [116,117]. Further studies suggest that the IL-7 pathway is inactivated in sepsis, although intrinsically intact, as ex vivo treatment of patients' cells with rIL-7 restored intracellular signaling [118]. The mechanisms through which IL-7 may combat sepsis-induced immune suppression include the reversal of lymphocyte depletion and the maintenance and expansion of T-cell receptor diversity. This is achieved through antiapoptotic and lymphopoietic effects on naïve and memory T cells [112,119], improved T-cell trafficking to sites of infection through enhanced expression of LFA-1 and VLA-4 [120] and reversal of monocyte deactivation through enhanced IFN-γ secretion [113,120]. In animal models of sepsis, rIL-7 improved survival [51,113,120,121] and the results of a phase 2 clinical study assessing its efficacy in patients with severe sepsis are pending (NCT02640807).
Interleukin-15 administration has been reported to reduce lethality in animal models of sepsis. The protective effect is thought to be because of the antiapoptotic properties of IL-15 and its stimulation of IFN-γ production [122]. However, unlike IL-7, severe toxic reactions were reported in patients with cancer treated with IL-15, which may limit utility in sepsis and make IL-15 a less attractive agent than IL-7 [3,123].
Cell-based therapies
Regenerative medicine and stem-cell–based therapies have experienced considerable public interest and have seen substantial growth in recent years [124]. Whereas application to the field of sepsis has not yet been explored fully, studies in animal models indicate that treatment with allogeneic mesenchymal stem cells (MSCs) reduces organ dysfunction and mortality [125,126]. The underlying mechanisms are thought to involve antimicrobial, antiapoptotic, and immunomodulatory effects [125–128]. A phase 1 clinical trial on the safety of MSCs for patients with sepsis has completed with results pending (NCT02421484), and a phase 2 study has been initiated (NCT03369275).
Another phase 1 study currently recruiting patients involves the administration of apoptotic allogenic mononuclear donor cells (NCT03925857). Although this study relies on the anti-inflammatory mechanisms triggered by apoptotic cells, it is an interesting new strategy to restore peripheral immune homeostasis in sepsis. Widespread clinical application may be difficult, however, because this requires locating matching donors, as well as preparation and administration of apoptotic cell suspensions, all while the patient is still in a hyperinflammatory state.
Therapies targeting epigenetic modifications
Classically, only adaptive immunity has been attributed memory functions and innate immunity was not thought of as having memory. However, the term trained immunity has been coined based on studies demonstrating that innate immunity can indeed be influenced by prior encounters with pathogens as well as their products [129]. Trained immunity is thought to be mediated by epigenetic reprogramming. For instance, β-glucan induces trained immunity through genome-wide changes in H3Kme3, H3K4me1, and H3K27 acetylation [130, 31]. These and other studies have suggested that trained immunity is the epigenetic and functional opposite of sepsis-induced immune tolerance [3,131]. Thus, therapeutically inducing trained immunity and reversing immune-paralysis through either β-glucan administration or direct manipulation of epigenetic enzymes (i.e., SIRT1 and SIRT2) [132,133] may be new strategies for immune-augmentation [134].
Other immune-augmenting therapies
Thymosin α-1 is a naturally occurring peptide currently being tested for its efficacy in sepsis. Thymosin α-1 (Tα1) is produced by thymocytes and interacts with Toll-like receptors, resulting in increased activation of dendritic cells, increased natural killer cell activity, increased levels of cytotoxic T cells, and a shift toward a TH1 response both in terms of cell subsets and cytokine levels [135]. Recombinant Tα1 is already in clinical use for the treatment of hepatitis B and C and chemotherapy-induced myelosuppression in different cancers [136–138], and is well tolerated with few side effects [139]. A meta-analysis of sepsis trials with Tα1 to date indicates a substantial reduction in mortality and disease severity [140]. A phase 3 clinical trial is currently recruiting patients (NCT02867267). Some of the Tα1 studies used a combination of Tα1 with ulinastatin. This naturally occurring protease inhibitor that inhibits several proinflammatory proteases and decreases proinflammatory cytokine levels. Meta-analysis of its use in combination with Tα1 has indicated a synergistic effect [140]. Ulinastatin is already approved for the treatment of severe sepsis in India and a phase 4 clinical study for patients with sepsis and septic shock is underway in China (NCT02647554).
Certain vaccines may also provide a boost to overcome sepsis-induced immune paralysis. The results of an initial clinical trial assessing the efficacy of BCG-vaccination enhancement of the immune response during endotoxemia are pending (NCT02085590). Another phase 2 trial assessing the efficacy of heat-killed Mycobacterium w administration is currently recruiting patients (NCT02330432).
Conclusions
A personalized medicine approach will most likely be the key to success for future clinical trials in sepsis. New biomarkers and functional assays (summarized in Table 1) will allow us to stage patients with sepsis and find a customized therapy regimen for each individual's immune phenotype (Fig. 3a and 3b). By limiting immune-suppressive therapies to patients in a hyperinflammatory state and immune-augmenting regiments to patients with immune exhaustion, treatment success will likely improve and adverse effects may be limited. It is encouraging that ongoing clinical trials have already begun enrolling patients based on their immune status. Parallel efforts in the search for biomarkers and immunomodulatory drugs for the treatment of sepsis will likely yield positive results and even the re-appraisal of therapies previously deemed unsuccessful may still show benefit when limited to the proper patient population.
Markers of Immune Activation or Suppression
TNF-α = tumor necrosis factor-α; LPS = lipopolysaccharide; sRAGE = soluble receptor for advance glycation end-products; IL = interleukin; HLA-DR = human leukocyte antigen-DR; IFN-γ = interferon-γ.
Footnotes
Acknowledgments
Literature review, drafting the article, preparation of figures: N.B. and C.C.C. Critical revision of the article, and final approval of the version to be published: N.B., C.T.S., P.A.C., V.N., and C.C.C..
The authors would like to acknowledge B. Beckmann for excellent support in designing the figures.
Funding Information
The project described was supported by Shriners Hospitals for Children award number 85000-CIN-17 (CCC) and T32 GM08478 (CTS) from the National Institute of General Medical Sciences. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institute of General Medical Sciences or the National Institutes of Health.
Author Disclosure Statement
The authors declare no conflict of interest.