
Abstract
The human microbiome is vast and is present in spaces previously thought to be sterile such as the lungs. A healthy microbiome is diverse and functions in an adaptive way to support local as well as organism health and function. Furthermore, a normal microbiome is essential for normal immune system development rendering the array of microbes that live in and on the human body key components of homeostasis. A wide array of clinical conditions and interventions including anesthesia, analgesia, and surgical intervention may derange the human microbiome in a maladaptive fashion with bacterial responses spanning decreased diversity to transformation to a pathogenic phenotype. Herein, we explore the normal microbiome of the skin, gastrointestinal tract, and the lungs as prototype sites to describe the influence of the microbiomes in each of those locations on health, and how care may derange those relations.
The microbiome is defined as the collection of genes harbored by micro-organisms associated with humans, whereas microbiota refers to the collection of particular micro-organisms (Fig. 1). 1 In humans, the microbiota consists mainly of bacteria with smaller proportions of archaea, eukaryotes, and viruses; a total of 10 to 100 trillion microbial cells comprise the human microbiota. The number of genes in the microbiome is staggering. Although the human genome consists of only 22,000 genes, the gut microbiome is composed of 3.3 million unique genes. 1 All sites of the body are colonized by micro-organisms, including the skin, respiratory tract, reproductive tract, and gastrointestinal tract. 2 Each of these sites demonstrate vast differences in micro-organism composition that varies between individuals, suggesting the presence of a personalized microbiome for each person. 3 For a given individual, the specific microbial composition of each site changes over time and can be affected by environmental influences including dietary intake, medications, and age. 4 As such, variations in the microbiome that maintain balance and homeostasis are the norm, rather than an absolute indication of disease.
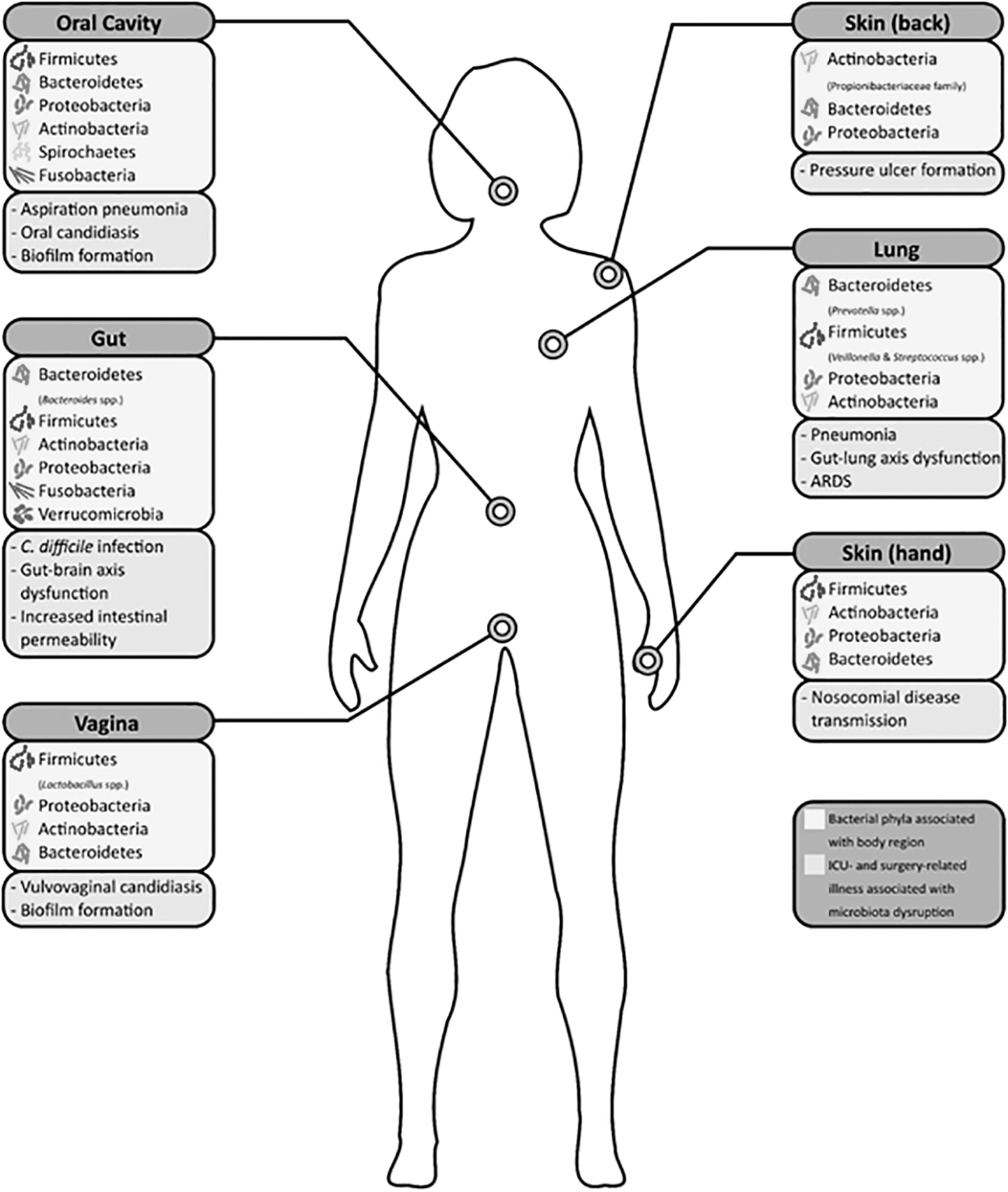
Dominant constituents of the microbiota of various regions of the body are presented in three tiers: location, phylum (with selected genera), and surgery and intensive care unit (ICU)-related conditions associated with microbiota abnormality.
The remarkable variability in microbiota between and within individuals was demonstrated by a study in which the skin flora on both hands of 51 healthy undergraduates was sequenced. Remarkably, the average amount of shared phylotypes between individuals was only 13%. More surprising was the demonstration that the flora of an individual's hands were not remotely similar; the left and right hand only shared an average of 17% of phylotypes. 5 These broad microbiota differences are readily contrasted with the highly preserved 99.9% genomic similarity between unrelated individuals identified by the Human Genome project. 6
Despite the great variability, the notion that there is a conserved core microbiota that characterizes a given body site is supported by a growing body of data. Qin et al., 7 as part of the Metagenomics of the Human Intestinal Tract (MetaHIT) project, assessed fecal samples from 124 individuals and sequenced 3.3 million non-redundant microbial genes (99% bacterial). The entire sample was estimated to contain up to 1,150 bacterial species, and that each individual would house at least 160 species. They identified 75 bacterial species shared in at least 50% of cohort individuals, and 57 species that were shared by at least 90%. However, the relative abundance of species varied quite widely—from 12-fold to 2,200-fold—within the 57 species. 7 Arumugam et al. 8 found a more similar composition of the gut microbiota. In looking at microbiome samples from 39 individuals across different continents, they documented three relatively distinct clusters of bacteria, termed enterotypes, that defined the samples but were not location specific. 8
Further studies demonstrate a more continuous nature to the gut microbiota; whether the gut microbiota exists on a continuum or comprises discrete communities of bacteria remains somewhat controversial.9,10 Even dissimilar microbiota can provide functional similarity within the core microbiome as distinct bacteria may demonstrate similar genetic pathways and therefore serve a similar function within the microbiome. Turnbaugh et al. 11 studied fecal samples of 18 individuals (from a larger sample of 154 adult female pairs of monozygotic and dizygotic twins and their mothers) and noted that although variations in the microbial community were prominent, the functional metabolic pathways demonstrated by the microbiome were relatively consistent. 11 Variations of the microbiome has been implicated in conditions in which genetic variation has failed to explain the observed discrepancies in phenotype, such as obesity, autism spectrum disorder, and various allergic- and immune-mediated diseases. 3 Therefore, we examine the role of the microbiome primarily of the skin, respiratory system, and the gastrointestinal tract as it relates to health, surgical evaluation, peri-operative management, and surgical critical care.
Gastrointestinal Tract
Clostridioides difficile infection
One of the most prominent examples of microbiome aberration leading to disease in the intensive care setting is that of Clostridioides difficile infection (CDI). Individuals who require prolonged antibiotic use may experience alterations in the gut microbiome that predispose to infection with Clostridioides difficile, which spreads through the fecal–oral route via ingestion of spores from asymptomatic carriers, patients, or the environment.3,12–14 Major risks for the development of CDI are antibiotic use, old age, and hospitalization. Although almost all antibiotic agents have been implicated in CDI, exposure to clindamycin, fluoroquinolones, cephalosporins, monobactams, and carbapenems appear to be most associated with CDI.12,13 Alteration of the gut microbiota as a result of even short courses of antibiotic therapy renders the patient susceptible to CDI during the time period for restoration of normal gut microbiome homeostasis. 13 Because antibiotic agents can decrease normal gut flora diversity, it is unsurprising that the absence of Bacteroides and Firmicutes species (more than 90% of the normal gut microbiota) play a major permissive role in establishing CDI.15–17 Relatedly, Deng et al. 18 found that a murine CDI model CDI demonstrated that prophylactic treatment with Bacteroides fragilis led to higher rates of survival, reduced disruption of the gut barrier, and improved diversity and abundance of gut bacteria. However, diversity is not the sole metric of gut microbiome balance and health. Firmicutes production of butyrate (a short-chain fatty acid [SCFA]) helps to maintain colonic epithelium integrity and regulate mucosal immune responses. 15 Although the administration of vancomycin and fidaxomicin (and previously metronidazole) are the initially indicated treatments of CDI, recurrent episodes have been successfully managed with fecal microbiota transplant in a durable fashion.13,15,19–23
Cognitive dysfunction
The gut–brain axis is the signaling pathway through which the gastrointestinal tract and central nervous system (CNS) interact. Gut microbiota appear to play a major role in the bidirectional communication between the gut and CNS. 24 Proposed mechanisms by which the nervous system exerts effects on the gut microbiota include: alteration of the luminal environment through changes in motility and secretion of mucus, gastric acid, bicarbonate, and other substances mediated indirectly by the autonomic nervous system; increasing intestinal epithelial permeability (known to be associated with stress and possibly mediated through catecholamine signaling induced increased mucosal permeability and decreased barrier function); and intraluminal release of catecholamines, serotonin, cytokines, and immune cells that directly alter the behavior and phenotype of gut micro-organisms. 25
Conversely, processes through which the gut microbiome interacts with the CNS include: producing metabolites such as SCFAs, γ-aminobutyric acid (GABA), and tryptophan metabolites such as serotonin that interact with the enteroendocrine and enterochromaffin cells of the gut, the mucosal immune system, and more distant targets via entry into the systemic circulation with potential blood–brain barrier passage; modulating enteric sensory nerves by producing a variety of signaling molecules that influence ligand-dependent ion channels; influencing the vagus nerve via metabolites; and direct activation of neurons as well as spinal afferents via circulating metabolites to trigger cerebral signaling.24,25
Indeed, an elegant murine study assessed the role of the microbiome in post-operative cognitive dysfunction (POCD), a condition characterized by reduced cognition and intelligence occurring after surgery that can persist for months.
26
Cognitive performance was assessed in light of the gut microbiome and metabolic products, serum metabolites, hippocampal immunohistochemistry. Compared with sham mice, post-operative mice demonstrated impaired memory, increased hippocampal levels of interleukin (IL)-6 and IL-1β, decreased gut proportion of Bacteroidales, Mucispirillum, and Clostridiales but an increased proportion of Escherichia, Shigella, Actinomyces, Ruminococcus, and Lachnospiraceae. These derangements correlated with derailed metabolic functional pathways involved in the metabolism of tryptophan, glutathione, histidine, and amino acids. Post-operative cognitive dysfunction was correlated with the proportion of gut microbiome comprised of Fusobacterium, Actinomycetaceae, Escherichia, and Shigella and the concentrations of
Although difficult to prove causation, this study offers compelling evidence to link deranged cognition with a post-operatively maladaptively altered gut microbiome. Moreover, it underscores the importance of maintain gut microbiome balance for neurocognitive homeostasis, and begins to challenge existing dogma related to delirium. Furthermore, it also raises new questions about how peri-injury β-blockade improves cognitive recovery and what overlap that therapy might have with the gut microbiome. 27
Sleep deprivation
Sleep deprivation and stress in intensive care unit (ICU) patients are extremely common and appear related to alterations of the gut microbiome. Sleep deprivation is associated with a greater Firmicutes to Bacteroidetes ratio and increased Coriobacteriaceae and Erysipelotrichaceae. Gut microbial diversity correlates with improved sleep efficiency and reduced sleep deprivation.28,29 In a recent study, gut microbiota diversity decreased after 24 and 40 hours of sleep deprivation in 25 healthy individuals. 30 Moreover, this was associated with impaired cognitive function and increased activation of inflammatory pathways. 30 The same group then performed fecal microbiota transplantation into germ-free mice using fecal samples obtained from the volunteers prior to, and following, sleep deprivation. Mice treated with fecal transplantation samples obtained after sleep deprivation evidenced increased activation of the toll-like receptor-4/nuclear factor (NF)-κβ (TLR4/NF-κB ) inflammatory pathway, increased intestinal barrier permeability, increased neuroinflammation, as well as cognitive impairment compared with mice receiving samples obtained prior to sleep deprivation. 30 This study clearly suggests a role for induced changes in the gut microbiome in neuroinflammation, cognition, as well as intestinal barrier function. There are readily identifiable implications for occupations, including medical professional training, noted for periods of sleep deprivation and its subsequent influence on long-term health and wellness. Similar notions should inform how critically ill patients have sleep enabled as part of an integrated approach to acute physiologic stress and recovery that also helps preserve microbiome balance.
Skin
Geodiversity, mosquitos, and infectious disease transmission
Skin is often considered to be a principally homogeneous organ with notable exceptions for locations that bear apocrine glands. This notion is quite flawed as skin demonstrates substantial diversity from fingertip whorls to different thickness on the plantar surface of each foot. Moreover, the skin microbiome is marked by geodiversity even within the domain of a single body part such as the arm or the back indicating a kind of geofencing. Some of this kind of data is derived from the study of mosquitos, the individuals who they bite, and the location of their initial and repeated bites. Mosquitos are responsible for the spread of a wide variety of viral infections ranging from malaria to dengue fever to yellow fever to a host of encephalitides. 31 Although malaria is not a major driver of morbidity or mortality in the United States, it is a worldwide health issue that engenders vast resource allocation and cost spread across an estimated 241 million cases and 627,000 deaths in 2020. 32 Mitigating mosquito-borne malaria infection is a public health initiative and is aided by understanding who is subject to mosquito attack. Unsurprisingly, mosquitos are drawn to certain scents, and those scents are generated by skin surface flora. 33 Moreover, mosquito preference for a specific individual—compared with an immediately adjacent and equally skin-exposed individual—is uniquely related to skin surface-generated bacterial volatiles or scents. 34 Therefore, it is equally unsurprising that fresh cleansing using an antibacterial soap retards biting, but serves as an only temporary measure. 35 These data suggest that our current approach to surgical site infection prevention, namely topical cleansing and prophylactic antibiotic agents for the vast majority of cases, may benefit from understanding a given patient's local flora. Presently, recommendations are derived from population-based studies and do not account for recent illness or antibiotic exposure and may not be properly aligned with a patient's geofence, a unique microbiome to reduce the incidence of surgical site infection (SSI).
Surgical site infection
Surgical site infections are commonly encountered, representing almost one-quarter of all adverse peri-operative events in the National Surgical Quality Improvement Program registry; they appear tightly linked to anticipated skin microbiota. 36 It is estimated that 70% to 95% of SSIs originate from skin or nasal flora and that failure to adequately control these micro-organisms provides a root cause for SSIs. 37 Microbes that are not pathologic under normal conditions may become directly introduced to new areas during surgery. The change in environmental conditions coupled with surgical stress can induce the elaboration of pathologic characteristics such as virulence factors that promote subsequent infection. 36 Relatedly, murine models note alterations in the gut microbiome after administration of opioid agents as well as with general anesthesia, both common events in surgical patients.38,39
Tissue mobilization and injury during therapeutic operation commonly leads to local inflammatory cell infiltration by a process termed trafficking. This adaptive process guides neutrophils and other elements of cell-mediated immunity to sites of microbe infiltration as well as sites of inflammation. Nonetheless, trafficking can also lead to unanticipated infection as methicillin-resistant Staphylococcus aureus (MRSA) that has colonized distant sites can engulfed by neutrophils that then traffic to surgical sites. Although those neutrophils participate in the expected inflammatory response they also release MRSA into the surgical site; this linked sequence of events is also known as the Trojan horse hypothesis.40,41 As a result, strategies to reduce the negative impact of the microbiome on surgical infection during the pre-operative and intra-operative periods include nasal decolonization (for those with MRSA carriage), antibiotic intestinal preparation, pre-operative nutrition support, home skin decontamination (commonly for joint arthroplasty), temperature regulation, pre-operative antibiotic administration (except for most clean cases), enhanced recovery after surgery (ERAS) pathways that support microbiome homeostasis, and pre-incision skin preparation.29,36,42–49
Strong evidence suggests the superiority of chlorhexidine-based skin preparants over povidone-iodine to reduce SSI. 47 The goal of application of antiseptics such as these is to reduce the number of micro-organisms, including commensals, to prevent their introduction into new environments (surgical wound, systemic circulation), where they can transform into pathobionts. 48 Different commensals demonstrate differential susceptibility to skin preparants in that povidone-iodine and alcohol both reduced flora resident on the volar forearm, but alcohol had no effect on back flora. 49 Moreover, skin flora generally recover from skin preparant-induced colony reduction by 12 hours after application.48,49 Therefore, interventions designed to support recovery of normal skin flora after operation may be unnecessary for clean cases where only skin preparation was used to reduce SSI incidence. It remains unclear whether such interventions would be of benefit for those who underwent skin preparation as well as prophylactic antibiotic administration. A particular target group may be those who undergo extended antibiotic prophylaxis, or to cover indwelling drains in specific patient populations as part of a local practice paradigm.50,51 At present, extended prophylaxis does not conform to major medical professional organization guideline recommendations to reduce SSI incidence.
Pulmonary
Relatively recently the presumed-to-be sterile normal bronchial airways were found to harbor micro-organisms such as Firmicutes, Bacteriodetes, Proteobacteria, Fusobacteria, and Actinobacteria, although in limited density. In its normal and balanced state, the lung microbiome is thought to modulate the immune system and inhibit proliferation of harmful bacteria and protect the pulmonary endoalveolar spaces from invasion. 52 Dickson et al. 53 describe three evidence-supported models of how the pulmonary microbiota might maintain as well as diverge from homeostasis.
First is the concept that changes in the lung microbiota come from microbe migration into or out of the lung, the death of resident microbes, and reproduction of resident microbes. Factors such as blood flow, intubation, mechanical ventilation, pH, temperature, and oxygen tension can influence the health and viability of the pulmonary microbiome and lead to conditions that predispose to proliferation of pathogenic bacteria. 53 Hyperoxia commonly occurs in the ICU and is associated with acute lung injury by a direct effect on type 2 pneumocytes, the generation of reactive oxygen species, and intracellular signaling cascades that rely on mitogen-activated protein kinases.54 55
Second, clinical conditions such as asthma, cystic fibrosis, chronic bronchitis, and acute lung injury as well as acute respiratory distress syndrome (ARDS) flood the alveoli with glycoprotein-rich mucus. This substrate supports bacteria growth as well as dysbiosis. The alteration in nutrient availability drastically perturbs homeostasis and results in a reduction in diversity of the microbiota and emergence of dominant pathologic species that enjoy a selective survival advantage.
Third, that micro-organisms respond to the signaling molecules used by human cells such as catecholamines, glucocorticoids, sex hormones, neurotransmitters, and cytokines is known as microbial endocrinology. For example, Pseudomonas aeruginosa, which demonstrates enhanced growth after catecholamine exposure, was found to be the dominant microbe in catecholamine-rich bronchoalveolar lavage samples obtained from lung transplant recipients.56,57 Therefore, there are a variety of influences that may alter the pulmonary microbiome as a matter for routine care as well as the evolution of certain clinical conditions regardless of concomitant infection.
Through these mechanisms, the disruption of the lung microbiota may be both the cause and the consequence of lung injury noted with critical illness or injury. Furthermore, the existence of the gut–lung axis, characterized by the complex interplay between these two organs through circulatory and lymphatic routes, means that influences within one system may impact the other. 58 It is known, for example, that septic mice and humans experiencing ARDS have enrichment of lung microbiota with gut microbes and instillation of lipopolysaccharide into the lungs of mice leads to changes in lung, blood, and gut microbiota.59,60 Therapeutic approaches based on this interaction have been described. A meta-analysis by Manzanares et al. 61 evaluated 30 trials in which probiotics (live, non-pathologic micro-organisms) were administered to critically ill patients and found that probiotic administration was associated with a reduction in overall infection, ventilator-associated pneumonia, and antibiotic use, without mortality benefit. Additionally, ingestion of dietary fiber, which is used by gut microbes to produce SCFAs, protects against pulmonary allergic inflammation and respiratory disease mortality.
A recent review explored the data underpinning the therapeutic targeting of the pulmonary microbiome in infection-related and inflammation-driven conditions. 62 Rational interventions, only some of which have been explored, range from antibiotic manipulation of pulmonary commensals to manipulation of molecular mechanisms such as quorum sensing inhibition to limit pathogenicity. Several barriers were identified in pursuing therapeutic targeting that may readily inform the articulation of appropriate pulmonary microbiome focused investigations. It is clear that ICU interventions that support a healthy pulmonary microbiome remain in their infancy but are anticipated to impact how clinical elements such as hospital-acquired pneumonia and ventilator-associated pneumonia are assessed especially as we move toward molecular identification of specific microbes (as opposed to culture-driven data). The ability to distinguish a normal pulmonary commensal from an invasive pathogen will also shape clinical metric definitions, antibiotic use, and potentially relieve the selection pressure for multi-drug–resistant organism pathogenesis.
Conclusions
The microbiome has profound effects on the development and normal function of every organ system. The normal microbiome's impact on the gut, cognition, skin, and lungs continues to evolve based on technological advances. This knowledge should prompt a re-evaluation of the interface of the microbiome with diet, behavior, pathologic response, surgical therapies, and critical care interventions. The impact of therapy on normal microbiome dynamics must be considered while treating patients with critical illness or injury to try to avoid triggering imbalance, dysbiosis, and pathobiosis to support desired and adaptive outcomes.
Footnotes
Authors' Contributions
Conceptualization: Smith. Data curation: Reynolds, Smith. Formal analysis: Reynolds, Smith. Writing–original draft: all authors. Writing–review and editing: Smith. All authors approved the final version.
Funding Information
This work was partially funded for Dr. Noorbakhsh's contribution by a T32 training grant (5T32GM095442-12).
Author Disclosure Statement
No competing financial interests exist.