
Abstract
Objective:
This study aims to compare the expression levels of tumor necrosis factor-alpha (TNF-α) and interleukin-1 beta (IL-1β) in osteoblasts infiltrated with Mycobacterium tuberculosis H37Rv (H37Rv) and Brucella ovis to understand the differential bone destruction in spinal tuberculosis (STB) versus Brucella spondylitis (BS).
Methods:
Primary osteoblasts were isolated and cultured from the cranial bones of 2–5 days old mice and characterized by alkaline phosphatase (ALP) staining and alizarin red staining (ARS). H37Rv and B. ovis were cultured to the logarithmic phase, and transfection solutions were prepared. Osteoblasts were infiltrated with these bacteria at various multiplicities of infection (MOI) and time points. Cell survival post-infiltration was assessed using CCK-8 to determine optimal infection conditions. Osteoblasts were divided into three groups: the H37Rv group (infiltrated with optimal MOI H37Rv), the B. ovis group (infiltrated with optimal MOI B. ovis), and a negative control group. TNF-α and IL-1β expression in the cytoplasm was observed using immunohistochemical staining, whereas their levels in cell supernatants were measured using enzyme-linked immunosorbent assay. Protein expression was analyzed by Western blot. Differences between groups were compared with using one-way analysis of variance and t-tests, with p < 0.05 indicating statistical significance.
Results:
Both H37Rv and B. ovis infiltrated osteoblasts, substantially increasing TNF-α and IL-1β expression. The H37Rv group showed substantially higher levels of TNF-α and IL-1β compared with the B. ovis group (p < 0.05).
Conclusion:
Infiltration of osteoblasts with H37Rv and B. ovis substantially increases TNF-α and IL-1β expression, with higher levels observed in H37Rv-infected osteoblasts. This overexpression may contribute to the more severe vertebral bone destruction seen in STB compared with BS.
Spinal tuberculosis (STB), often secondary to pulmonary tuberculosis, results from Mycobacterium tuberculosis proliferation in the spine. Because of its rich blood supply, STB is the most common form of osteoarticular tuberculosis. 1 Conversely, Brucella spondylitis (BS) arises from Brucella infection, where bacteria invade the skin or mucous membranes, proliferate in phagocytes, and disseminate via the blood stream to the spine. 2 Both diseases trigger inflammatory responses leading to vertebral bone destruction and present similar clinical challenges. Rasouli 3 found that BS typically causes infiltrative bone destruction, whereas STB causes multiple worm-like osteolytic lesions, suggesting different degrees of destruction.
Tumor necrosis factor-alpha (TNF-α) is an important member of the TNF family. 4 This potent cytokine promotes bone resorption through various signaling pathways (e.g., NF-KB, JNK, p38, ERK, Akt) and enhances osteoclast differentiation and survival.5–8 Interleukin-1 (IL-1), particularly IL-1α and IL-1β, is crucial for osteoclast proliferation and differentiation. 9 Exogenous IL-1α and IL-1β increase multinucleated osteoclastogenesis in vitro. 10 Thus, TNF-α and IL-1β substantially induce osteoclast activity and bone resorption.
Osteoblasts are essential in bone metabolism, regulating osteoclast function and acting as immunologically critical cells to fight infections. 11 Osteoblasts can activate osteoclast formation via the TNF-α and IL-1 pathways. 12 TNF-α and IL-1 produced by osteoblasts can activate osteoclast formation, with TNF-α acting directly on osteoclast progenitors and IL-1 inducing osteoclast activation independently of RANKL–RANK interactions.13,14
Bone destruction is a severe complication of bacterial infections, with bone metabolism maintained by a balance between osteoclasts (bone resorption) and osteoblasts (bone formation). TNF-α and IL-1β secreted by infected osteoblasts accelerate osteoclast differentiation, increasing activity and exacerbating bone destruction.5–10,13,14 This study hypothesizes that differential inflammatory factor levels contribute to varying degrees of bone destruction in STB and BS. Taking STB as an example, this study maps the hypotheses of possible mechanisms of M. tuberculosis after infestation of the vertebral body (Fig. 1). So, we investigate the reasons for these differences by comparing TNF-α and IL-1β expression following osteoblast infiltration by M. tuberculosis H37Rv (H37Rv) and Brucella ovis.

The hypothesis of mechanisms of bone destruction in spinal tuberculosis.
Materials and Methods
Reagents
The following reagents were used in this study: Colombian blood plates (BeNa Culture Collection, Beijing, China), tryptone soy broth (TSB) medium, and Middlebrook 7H9 broth medium (BD, USA). TNF-α and IL-1β enzyme-linked immunosorbent assay (ELISA) kits, whole protein extraction kits, and BCA protein determination kits were sourced from KeyGen Biotech (Nanjing, China). Additionally, we used Dulbecco’s modified Eagle medium (DMEM) and trypsin (Gibco, USA), fetal calf serum (Hyclone, USA), and alkaline phosphatase (ALP) and alizarin red staining (ARS) kits (Solarbio, Beijing, China). TNF-α and IL-1β antibodies and CCK-8 kits were procured from Abcam (UK).
Instruments
The major instruments utilized included optical and fluorescence microscopes (Olympus Corporation, Japan), a microwave shaker, a cell counter (Thermo Fisher, USA), and a SpectraMax M Series Multi-Mode Microplate Reader (Molecular Devices, USA). Cell cultures were maintained in a CO2 incubator (Fisher Company, USA).
Strains and cells
Brucella ovis (BeNa Culture Collection, Beijing, China) and M. tuberculosis (H37RVPMV261-GFP, kanamycin-resistant green fluorescent bacteria, Beijing Jingnuo Biotech Co., LTD.) were recovered and cultured. The surface of the ampoule tube was sterilized and opened in a biosafety cabinet. The ampoule was sterilized and opened, its top cauterized with an alcohol lamp, followed by the addition of sterile water to rupture it, and then cracked with tweezers. A 0.5 mL aliquot of sterile water was pipetted into the lyophilized tube to dissolve the powder, after which M. tuberculosis was cultured in Middlebrook 7H9 broth medium. Brucella ovis was plated on Colombian blood plates (200 μL each) and spread evenly. Both bacterial cultures were incubated at 37°C with 5% CO2 for 24 h. All procedures were conducted in a biosafety cabinet.
Primary cell extraction and osteoblast culture
Primary osteoblasts were extracted from the crania of 10 newborn SD rats (2–5 d old) from the Animal Experiment Center of Ningxia Medical University [Certificate of Conformity No.: SCXK(Ning)2023-0001] following the method described by Wong and Cohn. 15 Fetal rat cranial slices were separated, washed 2–3 times in phosphate-buffered saline (PBS) buffer with dual antibiotics, and digested with 0.05% trypsin at 37°C for 15 min. Digestion was terminated with 2 mL of DMEM and centrifugation (2,000 rpm, 3 min). The cell suspension was transferred to a DMEM medium containing 10% fetal bovine serum, 100 U/mL penicillin, and 100 µg/mL streptomycin and incubated at 37°C with 5% CO2 for 1–2 days. Cells were passaged when they reached 80%–90% confluency. The Ethics Committee of Ningxia Medical University approved this study.
Osteoblast identification
Alkaline phosphatase staining: Third-generation osteoblasts were seeded in 6-well plates (1 × 106 cells per well) and cultured for 1–2 days until 70%–80% confluent. Cells were fixed with ALP fixative for 3 min, washed with distilled water, and incubated with ALP substrate solution in the dark for 15–20 min. Methyl green staining was performed for 3–5 min, and cells were examined microscopically.
Alizarin red staining: Osteoblasts were seeded in 6-well plates and cultured for 24 h (day 0). The medium was changed on days 7 and 14. Cells were fixed with 4% paraformaldehyde for 30 min, washed, and stained with alizarin red solution for 5–10 min. Calcium nodule formation was observed under a light microscope.
Preparation of H37Rv and B. ovis transfection solutions
Mycobacterium tuberculosis was cultured in Middlebrook 7H9 broth medium for 31 days. Bacterial cultures (15 mL each) were centrifuged (2,500 rpm, 3 min), the supernatant was discarded, and the pellets were resuspended in 3 mL PBS. Brucella ovis colonies were cultured on Colombian blood plates for 48 h and inoculated into a TSB medium for 2 days. Optical density (OD) values at 600 nm were measured after 5 min of shaking. OD values were adjusted to 0.9–1.0, and bacterial counts were performed. Resuspended bacterial cultures were incubated at 37°C with 5% CO2 and stored with 10% glycerol at −80°C.
Detection of osteoblasts infiltrated by H37Rv or B. ovis
Osteoblasts were seeded in 6-well plates (1 × 106 cells per well) and cultured in DMEM with 10% fetal bovine serum for 24 h. Cells were infected with green fluorescent H37Rv at MOIs of 1:5, 1:10, and 1:20 and incubated for 24 h. Cells were washed with PBS (5–8 times), and fluorescence was observed under a microscope. Brucella ovis was used to infect osteoblasts at MOI 1:100 for 24 h, and extracellular bacteria were washed away with PBS. Cells were incubated with DMEM containing 1% gentamicin for 2 h, washed, and collected at 0, 2, 12, 24, and 48 h. Cell lysates were diluted and plated on Colombian blood plates to count colonies.
Selection of comparison transfection MOI replicates and duration
Osteoblasts were seeded in 96-well plates (1 × 105 cells per well) and cultured in DMEM with 10% fetal bovine serum. Cells were infected with B. ovis at MOIs of 50:1, 100:1, 200:1, and 500:1 and with H37Rv at MOIs of 15:1, 20:1, 25:1, and 30:1. A blank control group was included. Cell viability was assessed using CCK-8 assays at 2, 12, 24, and 48 h post-infection. Absorbance at 450 nm was measured, and cell survival rates were calculated. MOIs and time points resulting in 50% cell survival were selected for comparative experiments.
Cellular immunohistochemistry
Osteoblasts were seeded in 6-well plates and cultured for 24 h. Cells were infected with H37Rv (MOI = 1:20) and B. ovis (MOI = 1:100) for 24 h, with a negative control group. Cells were fixed with 4% paraformaldehyde for 30 min, permeabilized with 0.5% Triton X-100, and treated with 3% H2O2. Primary antibodies were added (diluted 1:1000) and incubated overnight at 4°C, followed by secondary antibody incubation (diluted 1:2000).
ELISA
Osteoblasts were seeded in 96-well plates and infected with bacteria. Supernatants from 24-h infections were collected, and TNF-α and IL-1β levels were measured using ELISA kits (six replicates per group).
Western blot
Total cellular proteins were extracted from osteoblasts infected with B. ovis (MOI 1:100) and H37Rv (MOI 1:20) for 24 h. Protein concentrations were determined, and samples were subjected to SDS-PAGE and Western blotting following standard protocols.
Statistics
Data were analyzed using SPSS 28.0 software and presented as mean ± standard deviation (x ± S). One-way analysis of variance was used for group comparisons, with p < 0.05 indicating statistical significance.
Result
ALP and ARS suggested that the primary extracted cells were rat osteoblasts
Alkaline phosphatase and ARS confirmed the identity of primary osteoblasts (Fig. 2). The cytoplasm of osteoblasts changed from colorless (a) to blue (b) after ALP staining, and nuclei appeared green after methyl green staining (b). Alizarin red staining showed calcium nodule formation at 7 days (c), with a substantial increase at 14 days (d).
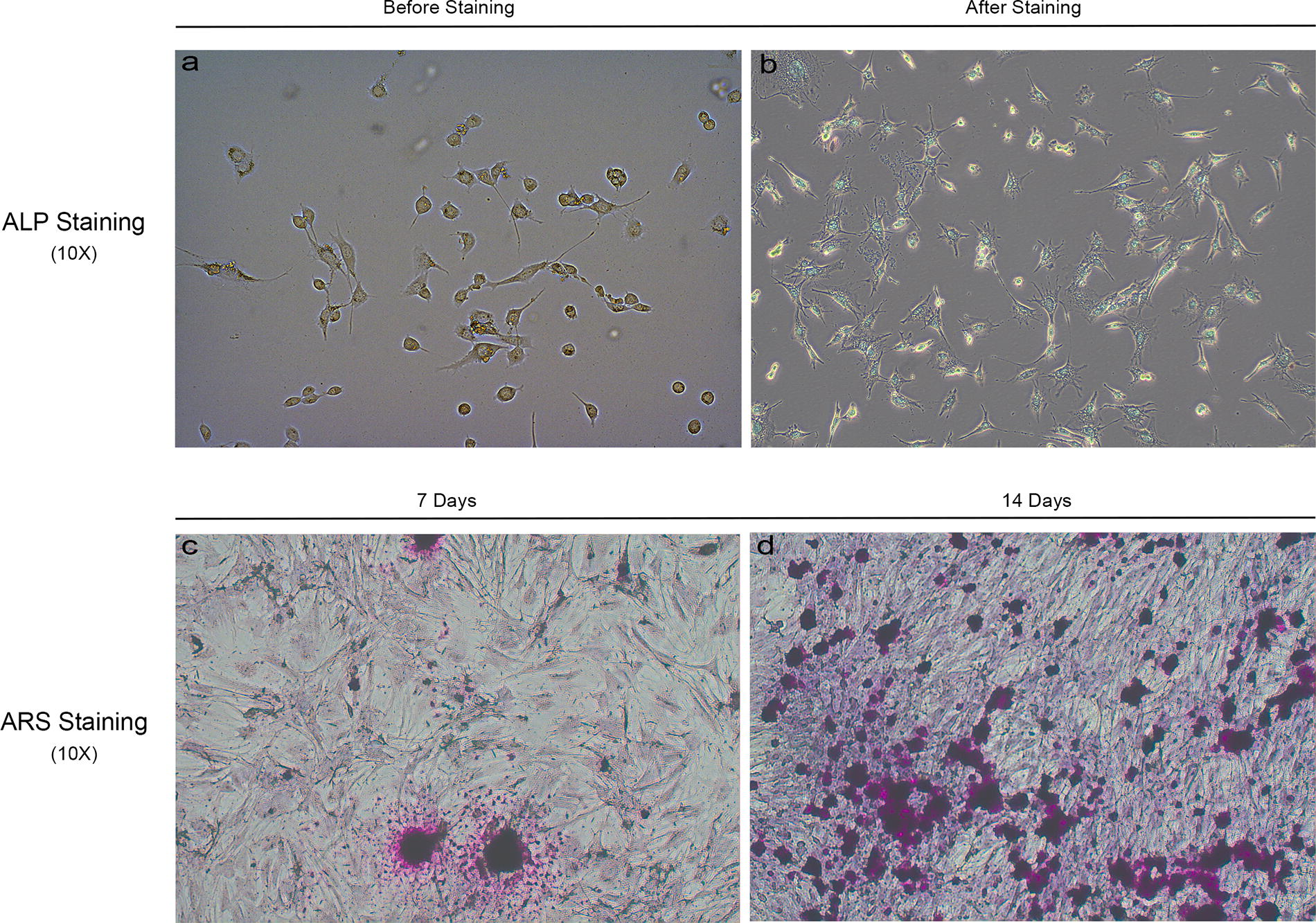
ALP and ARS of osteoblasts. Panel
H37Rv and B. ovis can infect osteoblasts
Detection of fluorescent H37Rv in osteoblasts (Fig. 3A): The intracellular fluorescence intensity of H37Rv increased with higher multiplicities of infection (MOI), confirming that the bacteria had penetrated and multiplied within the cells. Brucella ovis also can infect osteoblasts (Fig. 3B). Using a plate counting method, 100 μL of lysate, diluted 105 times, was inoculated on Colombian blood plates at different time points (0, 2, 12, 24, 48 h). Initially, a decrease in bacterial count was observed at 2 h post-infection. However, subsequent observations showed an exponential increase in B. ovis, indicating successful invasion and intracellular replication within the osteoblasts.

Detection of H37Rv and Brucella ovis infection models in osteoblasts.
These findings demonstrate that H37Rv and B. ovis can effectively infect and multiply within osteoblasts, as evidenced by fluorescence microscopy and colony counting methods.
Selection of infection replicates and experimental time for H37Rv and B. ovis in osteoblasts
The CCK-8 assay results, depicted in Figure 4, show the optimal conditions for achieving ∼50% cell survival following osteoblast infection by H37Rv and B. ovis. For H37Rv, the closest to 50% cell survival was observed at an MOI of 1:20, 24 h post-infection (Fig. 4A). Similarly, for B. ovis, the nearest to 50% cell survival was achieved at an MOI of 1:100 24 h post-infection (Fig. 4B).

Schematic diagram of the multiplicity and time of infection of H37Rv and Brucella ovis in osteoblasts. (
On the basis of these findings, the experimental conditions selected were an MOI of 1:20 with a 24-h incubation for the H37Rv group and an MOI of 1:100 with a 24-h incubation for the B. ovis group. These conditions will be used for subsequent experiments to ensure consistent and reliable results.
H37Rv and B. ovis infiltration elevates TNF-α and IL-1β expression in osteoblasts, with more pronounced effects observed for H37Rv
The immunohistochemistry, ELISA, and Western blot results for the three groups of cells (negative control, H37Rv, B. ovis) are presented in Figure 5. Immunohistochemical staining indicated that the cytoplasm of positively stained cells appeared brownish-yellow, whereas nuclei were blue because of hematoxylin staining (Fig. 5A). The negative control group showed no brownish-yellow cytoplasmic staining for TNF-α (a) and IL-1β (d), with nuclei stained blue-purple. In contrast, the B. ovis group displayed brownish-yellow cytoplasmic staining for TNF-α (b) and IL-1β (e). The H37Rv group exhibited the most pronounced positive pericellular staining for TNF-α (c) and IL-1β (f).
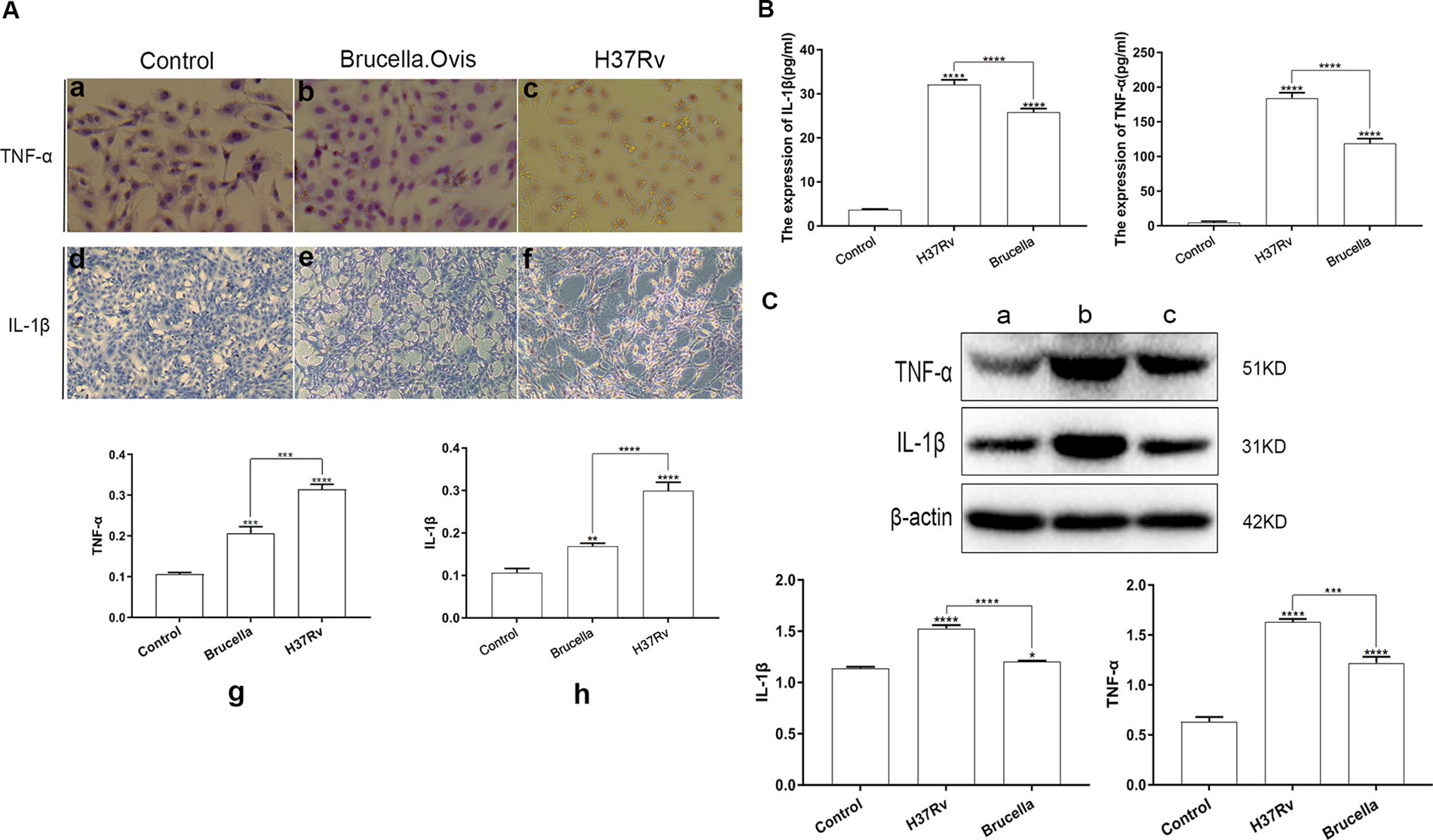
Expression results of IL-1β and TNF-α in three groups of cells (control group, H37Rv group, and Brucella ovis group). (
Quantitative analysis of the immunohistochemistry results using ImagePro Plus 6.0 is shown in Figure 5A (g-h). Both H37Rv and B. ovis infiltration significantly increased the expression of TNF-α and IL-1β in the cytoplasm compared with the control (p < 0.05), with the H37Rv group showing the highest levels (p < 0.05).
ELISA results (Fig. 5B) demonstrated that both H37Rv and B. ovis infection elevated TNF-α and IL-1β levels in the cell supernatants (p < 0.05). The increase was significantly higher in the H37Rv group compared with the B. ovis group (p < 0.05).
Western blot analysis of TNF-α and IL-1β protein bands (Fig. 5C), further quantified using Image J software, corroborated these findings. Both infections elevated TNF-α and IL-1β protein levels (p < 0.05), with the H37Rv-infected osteoblasts showing higher expression levels than those infected by B. ovis (p < 0.05).
Discussion
The mechanism of bone destruction in STB is a complex process influenced by M. tuberculosis infection and the body’s immune response. Inflammatory factors are expressed at various stages of M. tuberculosis infection.16,17 Therefore, studying these factors can help elucidate the mechanisms behind intervertebral disc and vertebral body destruction caused by M. tuberculosis.
BS, first described by Kulowski in 1932, 18 results from the invasion of the spine by Brucella species, leading to an inflammatory response. The precise mechanism of skeletal damage in BS is not fully understood, and it may involve direct bacterial action or immunopathologic processes triggered by innate immunity. 19 Clinically, the degree of vertebral bone destruction in STB differs substantially from BS, but the exact cause remains inconclusive. This study investigates whether bone destruction in these diseases is related to the elevated expression of inflammatory factors following bacterial infiltration of osteoblasts.
Both Brucella and M. tuberculosis can infect osteoblasts.20,21 This experiment confirmed that B. ovis successfully invades osteoblasts, as evidenced by exponential growth in colony numbers after lysis and culturing. Similarly, green fluorescent-labeled H37Rv infiltrated osteoblasts, covering the cell surface and increasing fluorescence with higher MOIs, indicating successful infiltration.
For comparison experiments, we used the MOI and time at which the survival rate of osteoblasts was 50%, as determined by the CCK-8 assay. The results showed that B. ovis achieved ∼50% osteoblast survival at an MOI of 100:1 for 24 h, whereas H37Rv achieved this at an MOI of 20:1 for 24 h. These conditions were selected for experimental comparison.
Osteoblasts can enhance osteoclast activity and cause bone resorption by indirectly secreting TNF-α and IL-1β. No previous reports have compared the expression of these cytokines following H37Rv and B. ovis infiltration of osteoblasts. This study hypothesized that the clinical difference in vertebral bone destruction between STB and BS might be related to the osteoblasts’ secretion of TNF-α and IL-1β. Mtb (H37Rv) promotes TNF-α expression in alveolar macrophages, 22 and infected macrophages and dendritic cells secrete IL-1β. 23 Brucella abortus induces TNF-α but not IL-1β secretion from B cells, 24 whereas B. ovis infection increases TNF-α and IL-1β secretion from macrophages and THP-1 cells.19,25
Our immunohistochemistry results showed that both H37Rv and B. ovis groups had higher TNF-α and IL-1β expression than the negative control group. Unlike Delpino MV and Fossati CA, 20 who found no TNF-α and IL-1β expression in B. abortus-infected osteoblasts, our results suggest species-specific differences. The H37Rv group had higher expressions of both cytokines than the B. ovis group, as confirmed by ELISA and Western blot. ELISA showed substantial differences in TNF-α and IL-1β levels in cell supernatants, with the highest in the H37Rv group. Western blot analysis also indicated statistically significant differences, with the H37Rv group showing the highest expression levels.
In summary, both H37Rv and B. ovis induce TNF-α and IL-1β expression in osteoblasts, with substantially higher levels observed in H37Rv-infected cells. Given that TNF-α and IL-1β play key roles in mediating bone destruction, this increased expression may explain the more severe vertebral bone destruction in STB compared with BS. This study offers valuable insights into the mechanisms underlying tuberculous bone destruction, providing a basis for further exploration of the differing degrees of bone damage between STB and BS, and establishing a foundation for future research on the pathophysiology of STB.
However, there are several limitations to this study that should be noted. First, the absence of in vivo experiment involving the use of P3 laboratory strains limits the interpretation of our findings. Second, mechanistic pathway analyses are needed to better understand why STB exhibits a higher capacity for bone destruction than BS. Finally, a combined approach, incorporating both in vivo and ex vivo models with osteoclasts, would provide more comprehensive insights into the effects of M. tuberculosis and Brucella on bone destruction.
Conclusion
Infiltration of osteoblasts with H37Rv and B. ovis substantially increases the expression of TNF-α and IL-1β, both of which are critical mediators of bone destruction. Notably, H37Rv-infected osteoblasts exhibit higher levels of these cytokines compared with B. ovis-infected cells. This heightened expression of TNF-α and IL-1β may play a pivotal role in the more severe vertebral bone destruction observed in STB compared with BS. These findings suggest that M. tuberculosis has a greater capacity to induce pro-inflammatory responses that lead to enhanced bone degradation, potentially explaining the more aggressive bone pathology in STB.
Footnotes
Acknowledgments
Thanks to each author for their hard work and the P2 laboratory platform provided by the Fourth People’s Hospital of Yinchuan. Thanks to the Ningxia Medical University Science and Technology Building laboratory teachers for their help.
Ethics Approval
The animal experiments involved in this study were approved by the Ethics Committee of the General Hospital of Ningxia Medical University (KYLL-2023-0130).
Author Disclosure Statement
The authors report no conflicts of interest in this work.
Funding Information
This project is financially supported by the