
Abstract
Abstract
We have previously shown that penile corporal structures engineered using autologous cells are able to achieve erection, penetration, and ejaculation. However, fully functional corpora could not be engineered because of the limited cellular content present within the corporal tissue construct. In this study, we investigated whether a dynamic seeding approach would improve cellularity within the corporal tissue construct and thereby restore normal erectile function. Corporal cells were either statically or dynamically seeded on acellular corporal tissue matrices and maintained in a bioreactor system. After 48 h, the cell–matrix complexes were implanted subcutaneously in athymic mice and analyzed for cell attachment, survival, and distribution using histological and molecular techniques. Native tissues and matrices without cells served as controls. The seeded cells attached and proliferated within the sinusoidal walls of the matrices. After completing the seeding, the DNA and cellular content of the dynamically seeded matrices reached 71% of normal corpora, whereas the statically seeded matrices reached 39% of normal corpora. These findings were confirmed histologically, biochemically, and using scanning electron microscopy. This study demonstrates that dynamic cell attachment, using a bioreactor system, leads to the formation of morphologically and biochemically improved corporal tissue, which may be useful for penile reconstruction.
Introduction
Reconstructive surgery has been a challenge because of the limited availability of penile tissue. 3 Various surgical approaches have been attempted to functionally and aesthetically restore the genitalia,1,4 including the radial forearm free-flap procedure 5 with or without autologous bone. 6 However, these procedures are associated with significant donor site morbidity and often require multiple operations for a satisfying aesthetic and functional outcome.
Tissue engineering follows the principles of cell transplantation, materials science, and engineering to create biological substitutes to restore and maintain normal function of various organs. 7 Advances in tissue engineering, for example, have allowed the creation of functional urinary bladders from patients' cells. 8 Development of a similar process for engineering of corporal tissue, which is responsible for erectile function, using patients' own cells would eliminate some of the aforementioned problems with penile reconstruction and allow for restoration of anatomically and functionally normal corpora.
We previously demonstrated that cavernosal cells seeded on collagen matrices lead to the formation of corporal tissues in vivo once the matrix is implanted. 9 Furthermore, functional penile corporal cavernosa tissue was engineered using acellular corporal cavernosa and autologous cells in a rabbit model. 10 The engineered tissue was structurally similar to normal, and erection, penetration, and ejaculation were possible. However, only approximately 50% of normal intracorporal pressures were generated in the engineered penises. Histological analysis of the engineered constructs revealed an overall normal corporal architecture with a lower cell density than in native corpora. 10 Lower density of intracavernous smooth muscle cells is associated with erectile dysfunction, 11 and we hypothesized that this is why the engineered corpora had subnormal intracorporal pressures.
Therefore, to engineer fully functional penile tissues, we investigated the improvement of cellular content of the engineered constructs. This could be achieved by using a cell population with a higher doubling time or by placing more cells on the constructs. Because our established culture protocols have shown adequate corporal tissue formation, we focused on increasing the cell number on the constructs by improving the cell seeding technique. We believe that this approach may eventually increase intracorporal pressures and fully restore penile function. Toward this goal, we investigated whether use of a bioreactor system that permits dynamic cell seeding would result in better morphological and biochemical engineered corporal constructs than with those obtained using conventional, static methodology.
Material and Methods
Cell harvesting
Corporal endothelial cells were isolated using an explant technique previously described.9,10,12 Briefly, cavernosal tissue biopsies from New Zealand rabbits were harvested in a sterile manner. Biopsy specimens were kept in Dulbecco's modified Eagle medium (Gibco Invitrogen Corporation, Grand Island, NY) on ice. Specimens weres rinsed with 1× phosphate buffered saline (PBS) and minced using iris scissors into pieces smaller than 2 mm3. Tissue fragments were placed on gelatin-coated 25-mm dishes containing endothelial basal medium-2 supplemented with bullet kit (EBM-2, Cambrex Bioscience, Walkersville, MD). The explants were maintained in a humidified atmosphere at 37ºC, 5% carbon dioxide. Endothelial outgrowth was monitored daily using phase contrast microscopy. Cells were subcultured every 4 to 5 days (1:3 splitting ratio) using standard techniques. Doubling time of the endothelial cells was approximately 3 days. After establishing the primary culture, endothelial phenotype was confirmed immunocytochemically using three antibodies specific to endothelial cells:13–17 anti-von Willebrand factor (vWF; 1:25; Santa Cruz Biotechnology Inc., Santa Cruz, CA), anti-KDR (1:25; R&D Systems, Minneapolis, MN), and anti-CD34 (1:50, BD Pharmingen, San Diego, CA) (Fig. 1). The cells were grown and expanded (4-5 passages) until sufficient cells were available to seed the constructs.

Immunocytochemical characterization of cultured endothelial cells: After establishing the primary culture, endothelial phenotype was confirmed using antibodies specific to endothelial cells: (
Collagen matrix preparation
Acellular matrices were derived from donor rabbit penile tissue as follows. Corpora cavernosa of New Zealand rabbits were obtained and prepared using a cell lysis technique previously described. 10 Briefly, total penile corporal tissues were dissected and treated with distilled water in a stirring flask at 4ºC for 1 day to lyse the cells in the tissue. Subsequently, the corporal tissues were treated with 1% Triton X100 and 0.1% ammonium hydroxide in a stirring flask at 4ºC for 14 days to remove cellular debris. The resulting collagen matrices were then rinsed with fresh distilled water at 4ºC for 2 days, followed by a treatment with 1× PBS for 24 h. To confirm the acellularity of the matrices, gel electrophoresis, spectrophotometric DNA analysis, and histological examinations were performed (Fig. 2). If cellular remnants were found in the corporal biomaterial, the entire decellularization process was repeated.

Acellular penile bioscaffold: (
In addition, purified DNA extracts were used to demonstrate acellularity. After purifying DNA from the matrices and normal tissue using a DNeasy kit (Qiagen Inc, Valencia CA), 50 μL of the purified samples was loaded on a 1% agarose gel containing ethidium bromide (10 mg/mL) and run at 5 V/cm. The appearance of a large band in or near the well area of the gel indicates the presence of DNA.
Once acellularity was confirmed, corporal matrices were lyophilized, sterilized in ethylene oxide, and stored in a desiccator until use. Eight entire acellular penile biomaterials were used for this study.
Cell seeding on matrices
Primary endothelial cells were detached from monolayer using a solution containing 0.05% trypsin and 0.01% ethylenediaminetetraacetic acid. The pellets were resuspended in EBM-2 containing 10% fetal bovine serum, and the cell number was counted using a hemacytometer. The cell-containing solution was diluted in EBM-2 to achieve the exact cell concentration per volume. Trypan blue staining was performed to confirm cell viability. Endothelial cells were then seeded on whole acellular corporal tissue matrices. In the statically seeded group, 6 × 107 cells per cubic centimeter biomaterial were suspended in EBM-2 and applied to the matrix with a syringe. The seeded cells were allowed to acclimate on the matrix for 120 min before the addition of EBM-2. The volume of medium with cells was equal to the fluid volume removed from the matrix. This procedure prevented cell loss due to excessive volume. Subsequently, 30 mL of medium was added to each of the 10-cm dishes, and the cells were allowed to attach for 4 days. The medium was changed every 24 h to ensure sufficient supply of nutrients. For the dynamically seeded group, the cells were applied in a stepwise fashion. In the first step, 3 × 107 primary endothelial cells per cubic centimeter were suspended in EBM-2 and the biomaterials seeded statically, as described above. The seeded constructs were placed in EBM-2 and allowed to attach for 48 h. To ensure consistency in cell preparation and seeding, the cell number was calculated thoroughly, and the same concentration of cells per volume was seeded on each scaffold. In an effort to minimize the discrepancy between each sample, we used the same passage cells and pooled together.
The second step included the use of a bioreactor system consisting of a closed glass beaker and a magnetic stirrer. The glass beaker had three individual openings on the top, allowing for placement of the constructs through the central opening and sterile medium changes through the two lateral openings. During incubation, all openings were loosely capped for gas exchange. The biomaterials were placed in the center of the stirring bioreactor using a long wire. The bioreactor was filled with EBM-2 containing primary endothelial cells at a concentration of 3 × 107 cells per cubic centimeter of biomaterial. The penile grafts were maintained in the bioreactor for 48 h. A stirring speed of 40 revolutions per min (rpm) was used to keep the cells in suspension. The EBM-2 was changed every 24 h to ensure sufficient supply of nutrients. The cells seeded in both groups were equal, totaling 6 × 107 cells per cubic centimeter of biomaterial. At the end of the seeding process, the seeded penile constructs were cut into 2-mm sections and used for immediate analysis or implanted in vivo. Unseeded controls were placed in EBM-2 for 48 h before use. Six seeded sections were used to analyze the DNA concentration in each group, and six sections were used to perform the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. In addition, six sections of native tissues and six sections of acellular biomaterial were used as controls.
MTT assay
The MTT assay was used to evaluate the influence of the bioreactor on survival and cellular growth of attached cells. After completing the 4-day seeding, six samples from each group were used and analyzed following well-established protocols. 18 Six constructs without cells were used as controls. Briefly, MTT solution containing 5 mg/mL of MTT in PBS was prepared. Seeded tissue samples were trimmed to 0.5 g and placed in a 24-well culture dish; 1 mL of MTT solution was added to each well, and the plate was incubated for 3 h at 37ºC. Afterwards, every well was washed twice with PBS, and the cells were lysed by adding 2 mL of 99% isopropyl alcohol. The plate was shaken at room temperature for 1 h, and the absorbance was measured at 540 nm using a plate spectrophotometer (ELX 800, BioTek Inc., Winooski, VT). The intensity of the blue color is directly proportional to the metabolic activity of the cells.
DNA analysis
DNA analysis was performed to precisely evaluate the cell densities of the seeded penile constructs. DNA was purified using a DNeasy kit and used according to the manufacturer's guidelines for DNA extraction from animal tissues. The resulting DNA concentration was then measured using a spectrophotometer (BioMate 3 series, Thermo Spectronic, Rochester, NY) to measure the ratio of absorbances at 260 nm and 280 nm wavelength (A260/280). Decellularized matrices served as controls (n = 6). To normalize the metabolic activity to cell numbers, the average ratio of MTT activity to DNA content was calculated. The cell distribution throughout the entire penile scaffold from proximal to distal was evaluated by analyzing five different sections of the same penile sample.
Scanning electron microscopy
Two seeded samples from each group were analyzed using scanning electron microscopy (SEM) after the completion of the 4-day seeding process to evaluate cell attachment, cell distribution, and cell density. Environmental SEM (Hitachi S-2600 N/H, Hitachi High Technologies America, Schaumburg, IL) requires no sample preparation and therefore minimizes the possibility of losing partially attached cells during alcohol washes or critical point drying. Representative areas in the upper right quadrant of the penile corpora were compared in all samples.
Animal study
The animal study was performed in accordance with the guidelines set by the Animal Care and Use Committee at Wake Forest University. Experiments were performed using athymic mice (Charles River, Wilmington, MA) as construct recipients. Animals were housed together, allowed free access to food and water, and maintained on a light–dark cycle of 12 h each. For implantation of constructs, all animals were anesthetized using ketamine (75 mg/kg) and xylazine (15 mg/kg). In each animal, two samples were implanted beneath the skin of the back. After 14 and 28 days, animals were sacrificed, and samples were retrieved. The retrieved structures were analyzed grossly, histologically, and with molecular studies.
Histology and immunohistochemistry
Tissue specimens were formalin fixed, paraffin embedded, cut in 5-μm-thick sections, and stained with hematoxylin and eosin. Immunocytochemical stains were performed on cultures cells grown on Lab-Teck chamber slides (Nunc, Rochester, NY) and on the retrieved specimens. Endothelial cells were identified using polyclonal anti-vWF (VWF C-20, goat polyclonal, sc-8068, F2905, 1:25, Santa Cruz Biotechnology Inc.), monoclonal anti-CD34 (RAM34 Rat, 553731, 22597, 1:50, BD Pharmingen), and polyclonal anti-KDR (biotinylated vascular endothelial growth factor receptor 2 (KDR), BAF357, CVN03, 1:25, R&D Systems, Minneapolis, MN). As secondary antibodies for anti-vWF, we used biotinylated anti-goat made in horse (BA-9500, Q0422, 1:250, Vector Laboratories, Burlingame, CA), and for anti-CD34 we used biotinylated anti-rat made in goat (BA-9400, 1:400, Vector Laboratories). Immunolabeling was performed using the VectaStain ABC avidin-biotin detection kit (Vector Laboratories) and visualized using 3, 3-diaminobenzidinetetrahydrochloride chromogen. All sections were counterstained with Gill's hematoxylin. Native penile tissue sections served as positive controls. Tissue sections without primary antibody were used as negative controls.
Western blot analysis
Western blot analysis for vWF was performed on protein isolated from some retrieved tissue specimens (n = 4). Protein samples were prepared using routine extraction methods. Briefly, the specimens were rapidly homogenized in standard lysis buffer (Tris-20 1 M, sodium chloride 3 M, Triton 10% with protease inhibitor) and incubated in the buffer on ice. After 30 min on ice, the lysates were centrifuged at 12,500 × g for 15 min, and the supernatants were kept. Protein concentration in each supernatant was determined using the Bio-Rad DC Protein Assay Kit (Bio-Rad, Hercules, CA). Aliquots of 20 μg of total protein were then separated using sodium dodecyl sulfate polyacrylamide gel electrophoresis (12% gel; 120 V and 200 mA). The proteins were transferred to nitrocellulose membranes. After transfer, the membranes were blocked with 5% bovine serum albumin (BSA). The blots were probed over night at 4ºC with the polyclonal vWF antibody mentioned above, washed, and subsequently treated with secondary antibody conjugates for 1 h at room temperature. Immunoblots were treated with an enzyme-linked chemiluminescence reagent (Western Lightning Plus, Perkin Elmer, Boston, MA) and exposed to X-ray film for 30 s to 5 min.
Statistics
All presented data are expressed as means and the corresponding standard errors of the means. For statistical analysis, we used SPSS v11 (SPSS Inc., Chicago, IL). Outliers were defined as having a z-score larger than + 3.0 or smaller than −3.0 using modified z-score (median of the absolute deviation). All results were analyzed using one-way analysis of variance. The groups were further analyzed using Bonferroni post hoc testing. An alpha of p > 0.05 was considered significant.
Results
Cell harvest, cell culture, and preparation of the scaffold
Primary rabbit endothelial cells were successfully cultured, expanded, and characterized using immunocytochemistry. The acellular penile grafts were produced successfully, and acellularity was confirmed using histology, gel electrophoresis, and DNA quantification (Fig. 2). Staining with hematoxylin and eosin showed intact micro-architecture of the corpora cavernosa without cell remnants. Gel electrophoresis showed no detectable DNA remnants, and the quantification of DNA according to spectrometery showed minimal DNA remnants in the scaffolds (19.0 ± 0.8 ng/mg, n = 6).
Seeding
The endothelial cells were used for the seeding experiments at passage 4 to 5. Static and dynamic seeding strategies were performed successfully, with the cells attaching to the bioscaffold in both cases.
Four days after statically or dynamically seeding the constructs, six samples from each group were analyzed using MTT assay. This assay, which measures mitochondrial activity, showed actively metabolizing, surviving cells in both groups (Fig. 3). However, dynamically seeded constructs demonstrated a statistically significantly more metabolic activity than statically seeded constructs (all comparisons between groups, p < 0.001). The absorbance for control, statically seeded, and dynamically seeded constructs were 0.000 ± 0.001, 0.029 ± 0.007, and 0.049 ± 0.009, respectively.
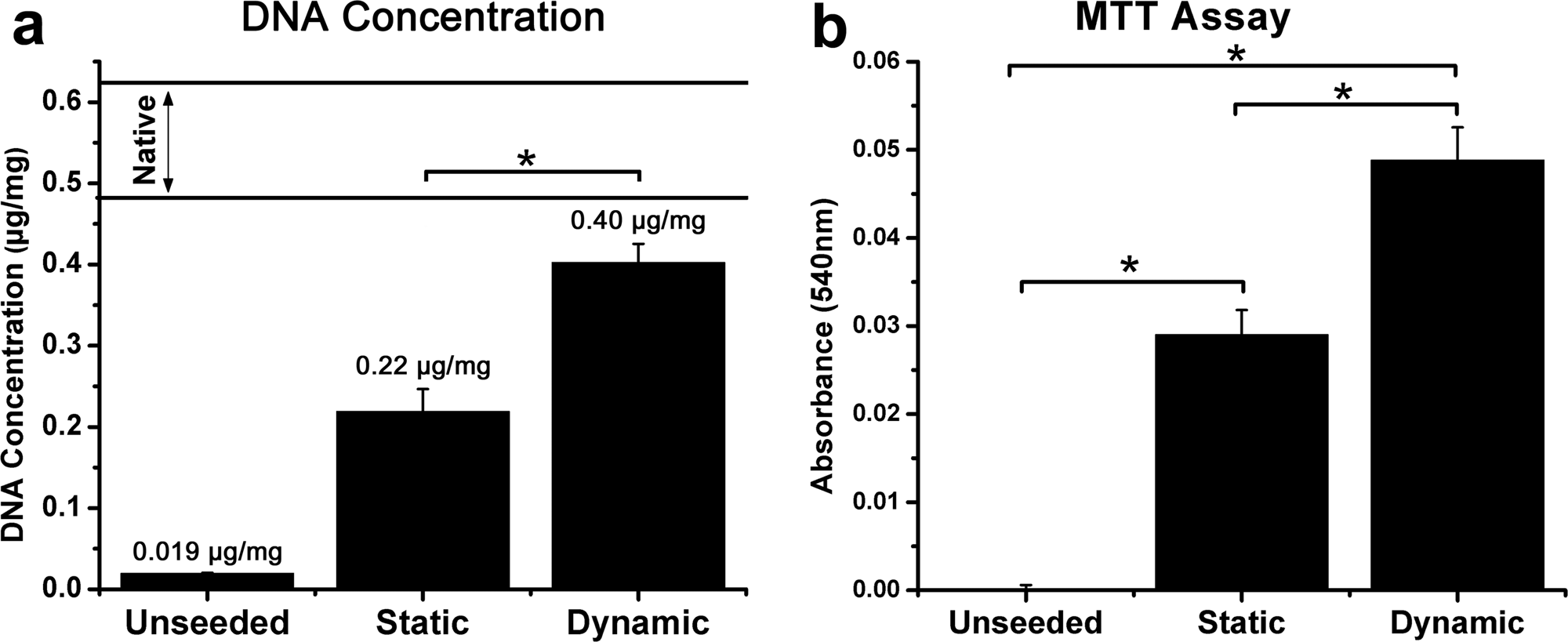
In vitro data: (
In addition, DNA content in statically and dynamically seeded samples was measured after completion of the seeding process (n = 6 in all groups). The amount of DNA was 0.019 ± 0.0008 μg/mg for unseeded biomaterial, 0.22 ± 0.028 μg/mg for statically seeded constructs, 0.40 ±0.023 μg/mg for dynamically seeded constructs, and 0.56 ±0.065 μg/mg for native penile tissue (Fig. 3). Static and dynamic methods resulted in statistically significant scaffold seeding (static vs acellular, p = 0.001; dynamic versus acellular, p < 0.001), and the dynamic method led to greater cellularity on the scaffold than with the statically seeded samples (static vs dynamic p < 0.001). The dynamically seeded samples were not shown to be statistically different from normal tissue (dynamic vs normal p = 0.03). According to these observations, the dynamically seeded matrices approach normal tissue in terms of metabolic activity.
The concentration of seeded cells along the length of the penile scaffolds was also measured by counting cells in five serial sections of each penile scaffold. When dynamic seeding was used, a more equal distribution of cells was observed. This observation was quantified by expressing the standard deviation as a percentage of the group mean (mean number of cells counted in the five sections). This value was 43.5% tor the static group and 21.5% for the dynamically seeded constructs, indicating a more even distribution of cells across the scaffold.
However, the overall ratios of MTT activity/DNA content, a measure of metabolic activity in regard to cell numbers, were similar, at 1.32 and 1.23 for statically and dynamically seeded constructs, respectively. This indicates that, although the seeding process used might affect cell distribution on the scaffold, it does not affect cell viability.
Scanning electron microscopy
SEM was performed after completion of the seeding process. All seeded samples showed endothelial cells within the sinusoidal walls of the matrices. Specimens seeded statically showed a lower cell count and more loosely attached cells than the dynamically seeded group (Fig. 4).
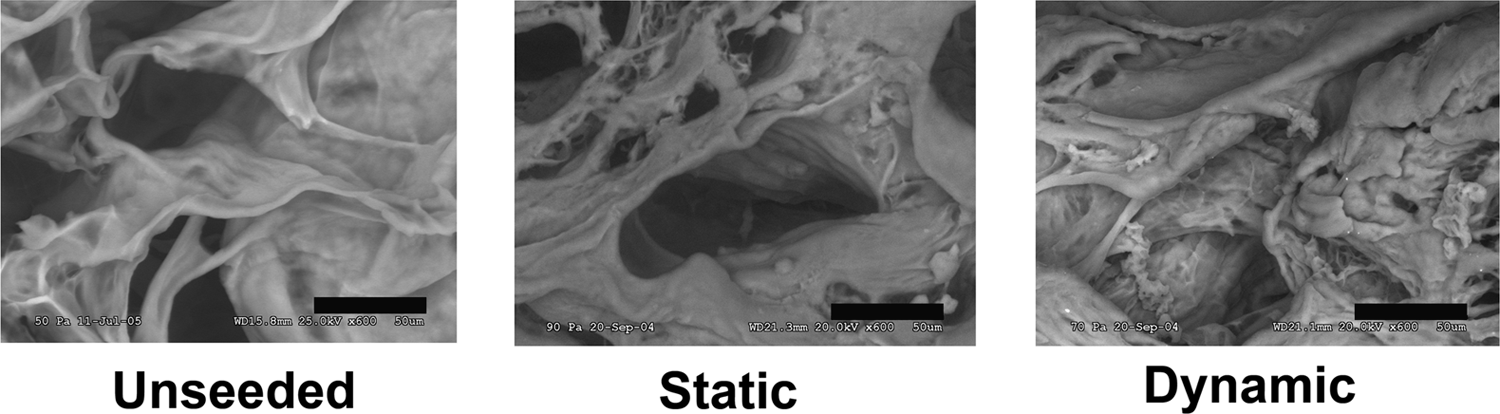
Environmental scanning electron microscopy of statically and dynamically seeded penile acellular grafts. The pictures show the top right quadrant of the corpora cavernosa. Static seeded constructs showed lower cell count and loosely attached cells. Dynamic seeded constructs showed higher cell density and good cell attachment along the sinusoids. Unseeded construct served as control. Scale represents 50 μm.
In vivo studies
Acellular, statically seeded, and dynamically-seeded constructs were implanted into the backs of athymic mice to study differences in in vivo responses. At 14 and 28 days post-implantation, the constructs were examined grossly and then removed for histological analysis. At harvest, there were no gross differences between the samples; all samples were well integrated into the subcutaneous tissue, and there were no signs of inflammation, infection, or fibrosis.
Histologically, hematoxylin and eosin–stained tissue sections showed minimal host tissue response. Neovascularization and some mixed cell infiltration were seen in both groups (statically and dynamically seeded constructs). There were no signs of inflammation or infection. All the specimens had the characteristic organization of corporal tissue, with thick collagen bundles lining the sinusoids. However, the dynamically seeded samples showed a higher cell density, more uniform cell distribution, and a tissue organization comparable with that of native tissue. The statically seeded samples showed a lower cell density, with cells organized into clusters that did not resemble native corpora (Fig. 5). The histological findings for both groups at 14 and 28 days were comparable.

Histology: Engineered corpora sections stained with hematoxylin and eosin. The top row shows low power images with approximately one-quarter of the corpora cavernosa (40×). The middle row shows representative high-power images (200×). The statically seeded group showed a lower cell density, with cells organized in clusters. The dynamically seeded group demonstrated morphological characteristics of native tissue, with significantly higher cellular content, better alignment along the sinusoids, more-uniform cell distribution, and denser corporal tissue formation. All sections stained positive for von Willebrand factor, demonstrating the endothelial origin of the cells in the sinusoids (bottom row, 200×). Normal cavernosal tissue serves as the positive control for the antibodies used. The endothelial lining of the central cavernosal artery stained positively. Color images available online at www.liebertonline.com/ten.
To confirm the origin of the cellular content within the engineered corpora, immunohistochemistry was performed using anti-vWF. Within the sinusoids of the corpora cavernosa, the majority of cells were vWF positive. The density of positively stained cells was higher than in native tissue, in which smooth muscle cells did not stain positive (Fig. 5). Theses findings were further confirmed using Western blot for vWF. All constructs seeded with endothelial cells were positive for vWF at all time points (data not shown).
Discussion
Organ damage or loss of function can occur from congenital disorders, cancer, trauma, infection, inflammation, iatrogenic injuries, or other conditions and often necessitates reconstruction or replacement. Depending on the severity of damage, autologous tissues can be used for reconstruction. However, for penile reconstruction, there is often not sufficient tissue, and there is a degree of morbidity associated with surgical procedures. Medical treatment options rarely replace the entire function of the penis. However, the replacement of deficient tissues with functionally equivalent tissues may improve the outcome for these patients. Engineered biological substitutes that can restore and maintain normal tissue function would be useful in tissue and organ replacement applications.
The engineering of fully functional organs requires normal tissue architecture, the use of tissue-specific cell types, and a cellular content comparable with that of native tissue. The key to regenerating homogeneous tissue includes stable cell attachment, cell growth, and a uniform distribution of the cells throughout the biomaterial.
To successfully engineer corporal tissues, extensive cell expansion in culture and an efficient seeding technique are required. Improving the seeding technique and loading more cells into the sinusoids of the corpora cavernosa allow for the formation of corporal tissue that resembles native tissue.
The use of acellular biomaterial derived from cavernosal tissue has the advantage of improving tissue organization. However, corpora matrix is a low-porosity biomatrix with sinusoids of varying size and shape, making it challenging to evenly load high cell contents. Cells in solution tend to follow the path of least resistance and build clusters in the larger sinusoids, leaving the small sinusoids cell free.
We have developed a cell-seeding system that allows for uniform distribution and higher loading density of cells into the acellular corporal biomatrix. To the best of our knowledge, this is the first report specifically investigating dynamic cell-seeding techniques for the engineering of penile tissues. Static seeding has been a routine approach for many tissue-engineering studies.19–20 Over the past few years, researchers began to use hydrodynamic forces to seed cells on scaffolds before implantation, but only the combination of dynamic seeding and the use of a bioreactor system resulted in a significant improvement in cell attachment and distribution.21,22 These early studies were performed using synthetic high-porosity biomaterials that are easier to seed.
Dynamic seeding, in combination with the use of a bioreactor, allows for higher cell yield, better attachment, and better distribution than static seeding. However, the use of a new device introduces new variables that have to be addressed. For example, cell survival is highly dependent on stirring speeds. In bioreactors using rotation speeds of more than 150 rpm, cell damage is known to occur. 22 We saw that speeds lower than 30 rpm lead to the formation of cell collections on the bottom of the bioreactor. In the present study, 40 rpm was used. Therefore, we did not expect to compromise cell survival during the seeding process or during the time in the bioreactor.
DNA assays were used to precisely evaluate the cell density achieved in our constructs. We demonstrated, through the use of the bioreactor system, an improvement in cell density of 32 percentage points (from 39% to 71% of the density of normal corporal tissue). To confirm the survival of the seeded cells, a mitochondrial activity assay was performed. The MTT assay, first described by Mosmann in 1983, 23 is based on the ability of a mitochondrial dehydrogenase enzyme in viable cells to cleave the tetrazolium rings of the pale yellow MTT reagent. This results in the formation of dark blue formazan crystals, which are largely impermeable to cell membranes, thus resulting in their accumulation within healthy cells. It is important to understand that the MTT results of three-dimensional (3D) constructs do not allow for quantification of cell number but show a relative comparison of overall metabolic activity in the seeded constructs. Therefore, a higher measured metabolic rate (formation of more-intense blue color) may be the result of a small grouping of very active cells rather than a representation of an actual increase in cell number. Because of this, normalization methods must be used, and those used for 2D cultures may or may not be accurate. However, the MTT assay is a powerful tool to compare different experimental conditions in 3D scaffolds. Our findings are consistent with results of others using dynamic cell seeding and synthetic biomaterials. 21
A second measure of well-being of the seeded cells is the ratio of MTT absorbance to DNA absorbance. The overall ratio of both groups did not show significant differences. Therefore, we can conclude that the use of the bioreactor increases cell number and does not impair the well-being of the seeded cells. The agitation of medium, improving the gas transport and distribution of nutrients throughout the construct, can explain this. These findings were further confirmed using SEM, which showed better attachment and higher cell density for the dynamically seeded constructs than the statically seeded tissues.
The histology and immunohistochemistry showed enhanced tissue organization when dynamic seeding was used. Not only were there higher cell densities and intact cell lining of the sinusoids, but there was also complete coverage of the sinusoidal space if dynamic seeding was performed. Immunohistochemistry and Western blot analysis further confirmed the aforementioned findings.
One limitation of the in vivo evaluation is the possibility that host endothelial cells could have participated in the regeneration of the sinusoids. However, it is well known that ingrowing host endothelial cells in wound healing build fine networks for future blood vessel formation24–26 and do not form patches of endothelial cells as demonstrated in this study. Therefore, we strongly believe that the patches of vWF-positive endothelial cells lining the sinusoids are likely to have derived mainly from the seeded cells and that the participation of host endothelial cells was not significant. This finding has been consistently demonstrated in our previous studies.
This study was designed to investigate whether it was possible to substantially improve cell density and distribution within the sinusoids of the corpora. Therefore, only one cell type was used, namely primary penile endothelial cells. However, this technique allows the loading of different cell types by mixing, suggesting that future studies using multiple cell types will be possible. We are currently conducting a study in which an entire anterior corporal tissue is replaced using this technique to determine whether fully functional erectile tissue could be engineered.
In conclusion, this study demonstrates that dynamic cell seeding, using a bioreactor system, leads to the formation of morphologically and biochemically better corporal tissue. The seeded cells attached and proliferated within the sinusoidal walls of the matrices. DNA and cellular content of the dynamically seeded matrices reached 71% of normal corpora, whereas the statically seeded matrices reached 39%. Greater cellular content within the corporal tissue matrix may improve corporal tissue function when implanted in vivo.
Footnotes
Acknowledgments
We thank Kuo-Liang Chen, M.D., for his technical assistance. We also thank Karl-Erik Anderson, M.D. Ph.D., and Jennifer L. Olson, Ph.D., for their editorial assistance. This research was supported in part by a grant for career advancement of the University Basel, Switzerland, and by the FAG Foundation, Basel, Switzerland.