
Abstract
Whole disc organ culture is needed for preclinical testing of biological repair of the degenerate intervertebral disc. Such organ culture is hampered by two major limitations: first obtaining adequate nutrition though the calcified cartilage endplates adjacent to the vertebral bone, and second by loss of tissue integrity if the endplates are removed from the discs. In this work we utilize a recently described technique for whole disc isolation that overcomes these problems, by removal of vertebral bone and the adjacent calcified portion of the endplate, and the construction of a bioreactor that permits long-term viability of these discs in loaded organ culture. The bioreactor consists of a culture chamber in which the disc can be dynamically loaded in a uniform manner. The culture chamber is large enough to accommodate discs up to 60 mm in diameter, and so is amendable to study both bovine and human discs. The discs are loaded in the culture chamber by upper and lower platens, which conform to the shape of the remaining cartilaginous endplate and permit fluid flow across its surface. The bioreactor is able to load the disc under a variety of conditions ranging from static to dynamic and from physiological to pathological, and monitor induced changes in disc height. To date, bovine caudal discs have been maintained viable in the bioreactor for up to 4 weeks without any appreciable loss of disc height under physiological cyclic load and, in principle, could be maintained in such a manner for several months. Such long-term organ culture is essential for studying biological repair of the disc.
Introduction
It is crucial to understand the interplay between mechanobiology, disc composition, and metabolism to understand the underlying cause of disc degeneration and to be able to study ways to regenerate the degenerate disc.10,11 Large animal models, such as ovine and porcine, have been accepted as good models for studying disc structure, geometry, biochemistry, and biomechanics,12–17 whereas small animal models, such as rabbits, rats, and mice, have been used to answer metabolic questions.18–25 While large animals have a disc structure similar to the human, they undergo degeneration slowly and are expensive to use. In contrast, small animals are relatively cheap and undergo degenerative changes rapidly, but their disc structure is different to the human, particularly in the maintenance of notochordal cells within the NP throughout life.26,27 An organ culture model using discs from larger animals is an appealing approach for allowing the entire disc to be studied in its native environment under strictly controlled condition. 28
Organ culture models using larger discs have been established, but often suffer from the inability to achieve long-term NP cell viability.29,30 This difficulty is attributed to insufficient nutrition supply, progressive calcification of the cartilaginous endplates, and the postmortem formation of blood clots blocking the transendplate nutrition path. Some models systemically treat the animals with anticoagulant before slaughter to overcome blood clotting and promote nutrient supply. 29 While this method will enhance disc nutrition, it does not overcome the nutritional barrier imposed by the calcified cartilage endplate (CEP). Other models remove bone and CEP to maintain transport of nutrients to the center of the disc.30,31 In this method disc nutrition is maximized, but at the expense of sacrificing disc integrity, since all naturally confining collagen fibers connecting the disc to its adjacent endplate are severed. The ideal disc organ culture system would eliminate the endplate barrier while maintaining disc integrity.
In the present work, a bioreactor has been developed that facilitates organ culture of intact large discs in a controlled dynamically loaded environment, and is used in combination with a previously reported harvesting method 32 that maintains the integrity of the IVDs by preserving the noncalcified part of the CEP with collagen fibers attached to it.
Materials and Methods
Tissue isolation and culture
Bovine caudal discs of young steers were obtained from a local abattoir. The four largest, most rostral discs were processed within 18 h postmortem, as previously described. 32 Briefly, skin, muscle, and ligaments were removed together with anterior and posterior bone elements. Discs were then isolated by parallel bone cuts close to the cartilage endplates (BEP discs), and the bone and the adjacent calcified part of the cartilaginous endplate was then removed (CEP discs) using a high-speed drill fitted with a surgical fluted ball burr. CEP discs were processed until the surface was soft and flexible without detectable calcified tissue.
The BEP discs were thoroughly rinsed in PBS containing 50 mM citrate to eliminate blood clots, and both CEP and BEP discs were then washed in Hank's solution (Sigma), 100 U/mL penicillin (Invitrogen), and 100 μg/mL streptomycin (Invitrogen) and placed in sterile polypropylene specimen containers (sterile 80 mL specimen containers; STARPLEX Scientific) containing culture media (Dulbecco's modified Eagle's medium with L-glutamine and 15 mM HEPES [Sigma], supplemented with 5% fetal bovine serum, 100 U/mL penicillin, 100 μg/mL streptomycin, and 50 μg/mL l-ascorbate). The specimens were initially cultured in the containers for 7 days without applied external load 32 to exclude discs with any sign of contamination carried over from the isolation procedure, and were then transferred to the culture chambers of the bioreactors. The same culture medium was used, and 60 mL of the total 120 mL medium was exchanged twice per week throughout the entire culturing period. After the nonloaded period, discs were loaded statically for 48 h at 0.1 MPa, allowing the discs to creep, thereby equilibrating its water content to the external load and intrinsic swelling properties. The discs were then dynamically loaded for 2 h at 0.1 Hz between 0.1 and 0.3 MPa peak-to-peak load, interrupted by 6 h 0.1 MPa static load, then again cycled for 2 h at the same scheme, and finally interrupted by 14 h 0.1 MPa static load. The diurnal scheme was repeated for 19 consecutive days. The arbitrarily chosen diurnal loading scheme may best be compared with that a person with a sedentary life style may exhibit. When the effect of load magnitude was evaluated, three different dynamic loads were compared 0.1–0.3 MPa (low), 0.1–0.6 MPa (medium), and 0.1–1.2 MPa (high), following the same diurnal scheme as above for 10 days. Discs were also cultured for 4 weeks or 10 days without load as a comparison.
Stress profilometry
Stress profilometry, 33 to quantify the axial stress across a disc diameter, was used in a preliminary study to identify and optimize axial load transfer across the NP and AF of the discs. A total of 13 bovine caudal discs, isolated using either the BEP or CEP method, were compared. The circumference of the disc was measured and the cross-sectional area calculated assuming it to be round. Flat surface porous interface platens were used to sandwich the isolated discs. In the BEP group (n=6), serving as the native control, the platen covered the entire flat cut bone surface. In the CEP group (n=7), two different porous platen designs were used. For the partial coverage (CEPpart), platens were selected from an array of 1-mm-increment platens ranging from 18 to 23 mm in diameter, to match a 50% normalized disc cross-sectional area. Full coverage platens (CEPfull), similar to the BEP group, covered with overlap the disc's entire endplate.
Controlled axial disc loading was carried out on an 858 Mini Bionix (MTS). All discs were preconditioned for 5 min at 0.35 MPa load, and stress profilometry was then performed at 0.35 MPa. A 1.3-mm-diameter needle with a 1.5-mm-diameter pressure sensor, mounted 3 mm distant from the tip and facing to the surface, was inserted through the annulus in an arbitrary rotation direction, but always through the center of the disc and in its mid plane, until the sensor was visibly perforating the opposite annular region. The needle was then mechanically coupled to a resistor-based linear position sensor. With the sensor facing upward (vertical), the needle was steadily and slowly drawn backward across the entire disc. Continuous recording of positional data and pressure data and plotting against each other allowed drawing the stress profile. The needle was then re-inserted through the same needle track, but this time pulled across with the pressure sensor facing sideways (horizontal). Pressure transducer readings were plotted against needle position readings. Linear model fits with a p-value of 0.05 were used to compare group means for the various loading parameters and sensor orientations.
Bioreactor design
The bioreactor consists of two subsystems: (1) a culture chamber used for culturing and (2) a loading frame mated to the latter to apply controlled axial loading while measuring axial displacement. Bioreactors were used aggregated as a triple unit, with each bioreactor functioning independently.
Culture chamber and organ culturing
All components of the bioreactors are illustrated in Figure 1 and a close-up of the culture chamber is shown in Figure 2. The culture chamber was designed to accommodate disc specimens ranging from 20 to 60 mm in diameter. The disc was sandwiched between a top and bottom porous platen made from a 3.2-mm-thick sheet of hydrophobic polypropylene, having an average pore size of between 90 and 120 μm (Scientific Commodities Inc.). The platens were fitted in a recess of a holder machined from polyetheretherketone (McMaster-Carr). The holders were in a range of 18 to 23 mm diameter, in 1-mm increments, and were selected to match the actual disc's NP area. The latter was assumed at 50% of the total disc's cross-sectional area, necessitating a holder diameter to equal 71% of the disc diameter. An array of 3 vertical short stainless steel pins, forming an equilateral triangle arranged around the lower holder, acted as a fence around the disc periphery to prevent the disc from migrating sideways while being loaded. They could be adjusted in three positions to accommodate discs of varying diameters. The top holder extended to a vertical plunger, with the recess in fluid communication with a central bore and a fluid inlet used to flush media through the top porous platen. The bottom holder was seated firmly in the base plate made from Teflon (McMaster-Carr), again with a central bore and a fluid inlet used to flush media through the lower porous platen. A dome-shaped common fluid outlet, slightly elevated to not be plugged with debris, was located at the bottom of the culture chamber.

The bioreactor system consists of two subsystems: culture chamber (cc) and loading frame (lf). The culture chamber has a base plate (bp) and cover plate (cp), with the disc (d) placed between the two porous platens. The loading frame is a triple unit, each using a pneumatic actuator (pa) controlled with a proportional pressure control valve (pcv). A load cell (lc) and linear variable displacement transformer (LVDT) measure load and displacement. The actuator is connected (con) to the piston (p) of the culture chamber. Color images available online at
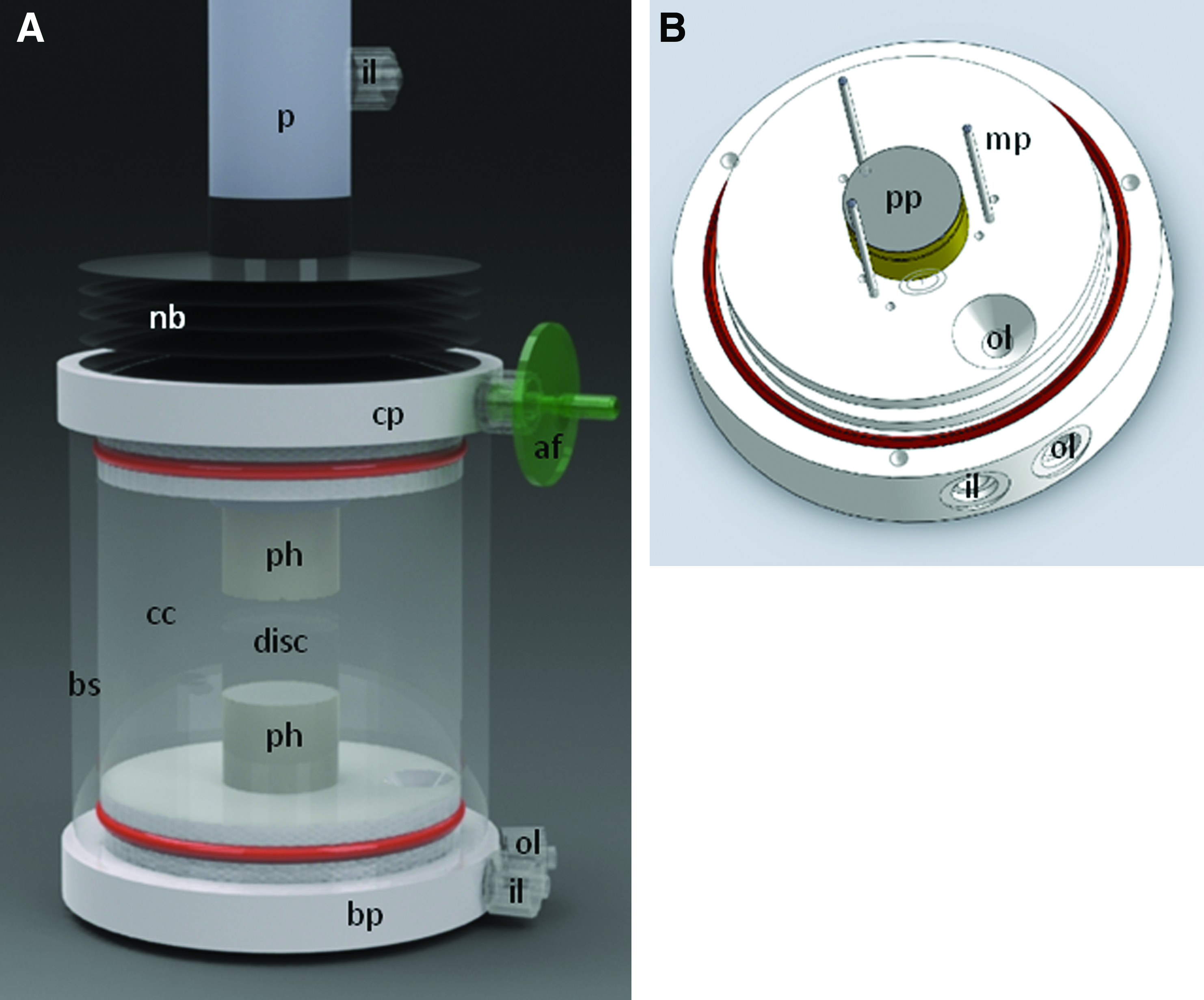
Once the CEP disc (n=6) was placed on the lower platen, the chamber was assembled. A borosilicate glass cylinder (McMaster-Carr) was pulled onto the base plate and topped with a Teflon cover plate. Both the base and cover plates were sealed against the glass cylinder with large O-rings, forming a fluid tight culture chamber. The cover plate, breached in the center with the plunger that was lowered to the disc under direct sight, was sealed off with a Neoprene bellows (
Loading frame
Once assembled and primed with medium, the culture chamber was mated with the loading frame. The plunger was connected to the pneumatic actuator, whereas the base plate was resting on the base of the frame. The main components of the loading frame subsystem (see Fig. 1) for each bioreactor unit were a Load Cell (OmegaDyne Inc), a Linear Variable Displacement Transducer (LVDT; OmegaDyne Inc.), a proportional Pressure Control Valve (VP50; Norgen Pneumatics Ltd), and a pneumatic Rolling Diaphragm linear actuator (1.7DAS; Illinois Pneumatics Inc.).
The proportional valve was used to regulate the input pressure to the pneumatic actuator. A stainless steel axial rod with a 25.4-mm base was passed through an anti-shear bracket connected to the plunger of the culture chamber such that only an axial linear force was transferred to the disc specimen. A input/output controller (U12; LabJack Corp) was used to continuously read load cell data, with the computer using a closed loop feedback to calculate the optimal pressure for the pneumatic actuator controlled with the proportional valve. Data were collected from each bioreactor unit separately. The axial load was recorded from the load cell, the pneumatic actuator displacement from the LVDT. The loading frame subsystem was controlled using software developed in LabView (Version 6.1). A Graphical User Interface was designed to operate the loading subsystem and also to observe and gather data. The software was configured to use a closed loop feedback to continuously monitor the applied load using the load cell, compare this value with the target value, and if required adjust the proportional valve until the measured value was within±5% error of the target value.
The bioreactor triple unit was sized to fit in a standard incubator, kept at a constant 37°C and 5% CO2 concentration. The peristaltic pump was placed outside the incubator. Compressed air supply lines, equipped with an in-line filter and connected to the proportional valves, were fed into the incubator through an access port. Likewise, all electrical connections were fed through an access port and interfaced with the LabJack or amplifier circuit.
Load displacement behavior
Disc height was measured with a calliper equipped with a custom measurement shoe matching the size of the NP area at the time of harvesting and at the end of the experiment when removing the disc from the chamber. Similarly, the disc circumference and weight were recorded. During mechanical loading in the culture chamber axial load and vertical displacement data were sampled continuously at 0.2 Hz, and data were plotted against time. The average disc height (data from calliper and LVDT) was compared for different time intervals: before and after free swelling, after preconditioning, and after cyclic loading.
Cell viability
After culture, a 1-mm section was taken through the center of the disc using a custom-designed cutting tool as described earlier. 32 The entire tissue section was incubated in serum-free medium containing fluorescent dyes (Live/Dead®; Invitrogen), according to the manufacturer's instructions, using 1 mM each of calcein (494/517 nm) and ethidium homodimer-1 (528/617 nm, in the presence of DNA). The status of the cells was observed using an inverted confocal laser scanning microscope (CLSM) (Zeiss LSM 510). Twenty consecutive 6-μm sections were imaged, and then CLSM stacks were split into single images. Five images were selected, merged, and saved as single-color JPEG files (red and green separate), and the labeled cells were quantified separately using the CellC software (Matlab). 34 The live-to-dead cell ratio was then calculated from the proportion of green and red cells. Unpaired t-test was used to calculate significance, and a p-value of <0.05 was used as the criterion for a significant difference.
Extraction and western blot analysis of ECM proteoglycans
NP tissue (n=5 per condition in the 4-week and freshly isolated group, and n=3 per condition in the 10 day group) was sliced thinly and extracted on a wet weight per volume basis using 15 volumes extraction buffer (4 M guanidinium chloride, 50 mM sodium acetate, pH 5.8, 10 mM EDTA, COMPLETE®; Roche). Tissue samples were incubated at 4°C under continuous agitation for 48 h. The extracts were then cleared by centrifugation at 16,000 g for 30 min. Five microliters of aliquots of disc extracts were precipitated by the addition of 9 volumes of ethanol, washed twice in 95% ethanol, and finally lyophilized. Precipitated material was dissolved in 10 mM sodium acetate solution (pH 6.0) and digested with keratanase II (Seikagaku) at 0.1 mU per 5 μL extract for 6 h. The solution was then adjusted to 100 mM Tris and 100 mM sodium acetate (pH 7.3) and digested overnight with chondroitinase ABC (Seikagaku) at 1 mU per 5 μL extract and finally mixed with SDS sample buffer. Treated samples were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (4%–12% NOVEX® gels) under reducing conditions, and separated proteins were transferred to nitrocellulose membranes. 35 After transfer, the membranes were blocked with 3% skim milk powder in 10 mM Tris-HCl, pH 7.4, 0.15 M NaCl, and 0.2% Tween (blocking buffer), and then incubated with antibodies recognizing the G1 domain of aggrecan at a 1:1000 dilution in blocking buffer containing 3% bovine serum albumin, followed by the secondary antibody conjugated with HRP (1:1000 dilution) in blocking buffer containing 1% dried skim milk. The bound antibody was observed by chemiluminescence (GE Healthcare) using a LAS 400 image analyzer (GE Healthcare).
Results
Design optimization of the bioreactor system
Over the course of about 2 years, with at least three marked design iterations, the construction and handling of the bioreactor system was significantly improved. Preliminary experiments indicated that the material of the porous platens is of great importance. Initially, we used hydrophilic polyethylene platens, which resulted in protein precipitation and clogging, leading to impaired medium flow, excessive pump pressures, and ultimately NP cell death in the center of the disc. We then turned to hydrophobic polypropylene platens, which avoid protein precipitation. Occasionally, small debris from the porous platens was observed floating on the media surface, but it seemingly did not affect medium flow or culture results.
Bovine caudal discs are readily available and survive for long periods in unloaded culture when isolated with intact CEP. 32 However, they have concave CEP and flat platens may therefore overload the annular disc region. For initial culturing experiments we used large and flat loading platens, similarly to the CEPfull type platens in combination with CEP discs. Marked cell death was observed in the AF regions (data not presented). Stress profilometry was therefore performed to evaluate load distribution in the discs with different geometries of loading platens attempting to find a shape that mimics native load transfer in the CEP discs. Three scenarios were tested: discs with intact BEP, representing the native form, were loaded with full coverage platens. Discs with removed BEP but preserved cartilaginous endplates (CEP) were loaded with either full coverage (CEPfull) or partial coverage porous (CEPpart) platens (Fig. 3). The CEPfull values were significantly different (p<0.001) from the CEPpart and BEP values. The latter two values were not significantly different (p>0.05). Of particular note, the pressure values for horizontal and vertical sensor orientation were equivalent (Fig. 4), indicating hydrostatic loading. With CEPpart platens, load distribution indicated load transfer as evidenced with pressure peak readings in the AF, comparable with that measured in the BEP discs. The pressure was marginally lower in the CEPpart group than in the native BEP group, but with very predictable amplitudes and low error repeatability. CEPpart platens were therefore used in the subsequent culture experiments.
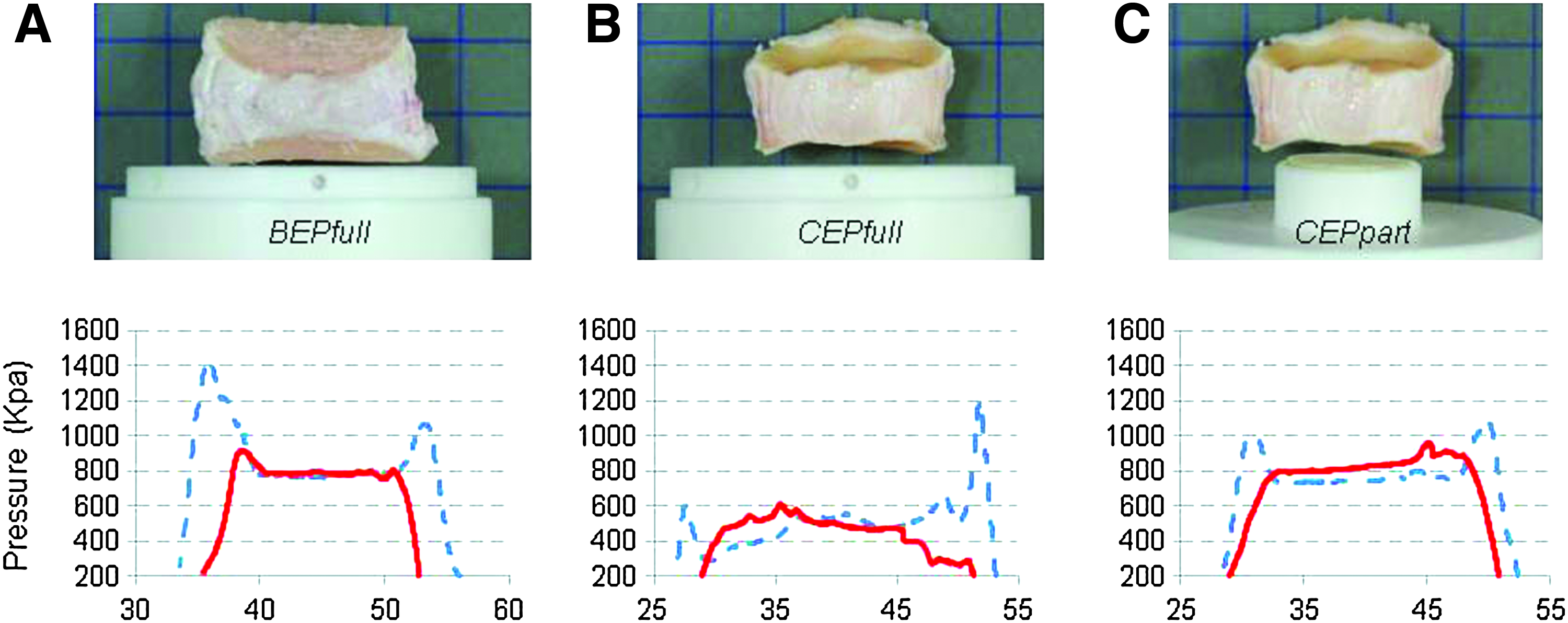
Stress profilometry for BEP, CEPfull, and CEPpart disc groups. Three platen configurations were tested:
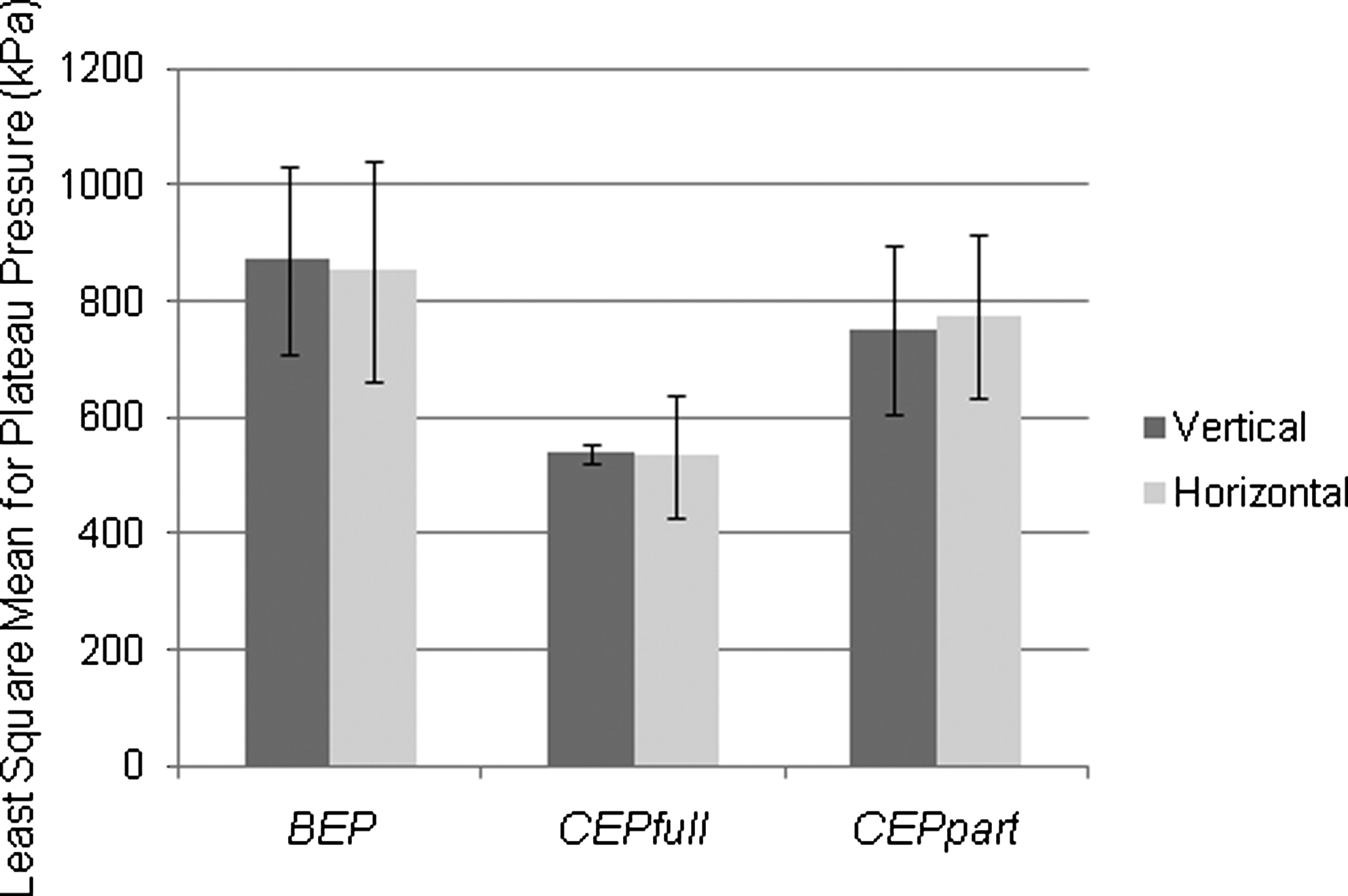
Pressure profilometry readings for NP plateau pressure during horizontal and vertical sensor orientation. Comparison for average plateau pressures (group mean with±1 SD error bars) for each of the three platen configurations. Horizontal (H) and vertical (V) sensor orientations render very similar results. Full coverage platen pressures were significantly lower compared to the other groups.
Biomechanical and biological data
Five CEP discs were maintained in culture for 28 days. Figure 5 displays typical load and displacement curves, both plotted against time for the period from day 8 to 28, during which the disc was loaded in the culture chamber. The discs expand during the first 7 days when cultured without load, and lose height again when being loaded, initially about 1.5 mm during the 48 h preloading period, and later about another 2 mm during low cyclic loading (0.1–0.3 MPa). After about 7 days loading the disc height stabilizes until the end of the experiment (Fig. 5). Table 1 summarizes individual and average values for disc height, disc diameter, and weight at different stages of the experiment. On average the discs decreased 7.7% (range 0.7% to 20.9%) in height, increased 5.9% (range 0.9% to 19.5%) in diameter, and increased 2.5% (range −3.9% to 10.6%) in weight when compared to the time of isolation. These changes are quite minimal.
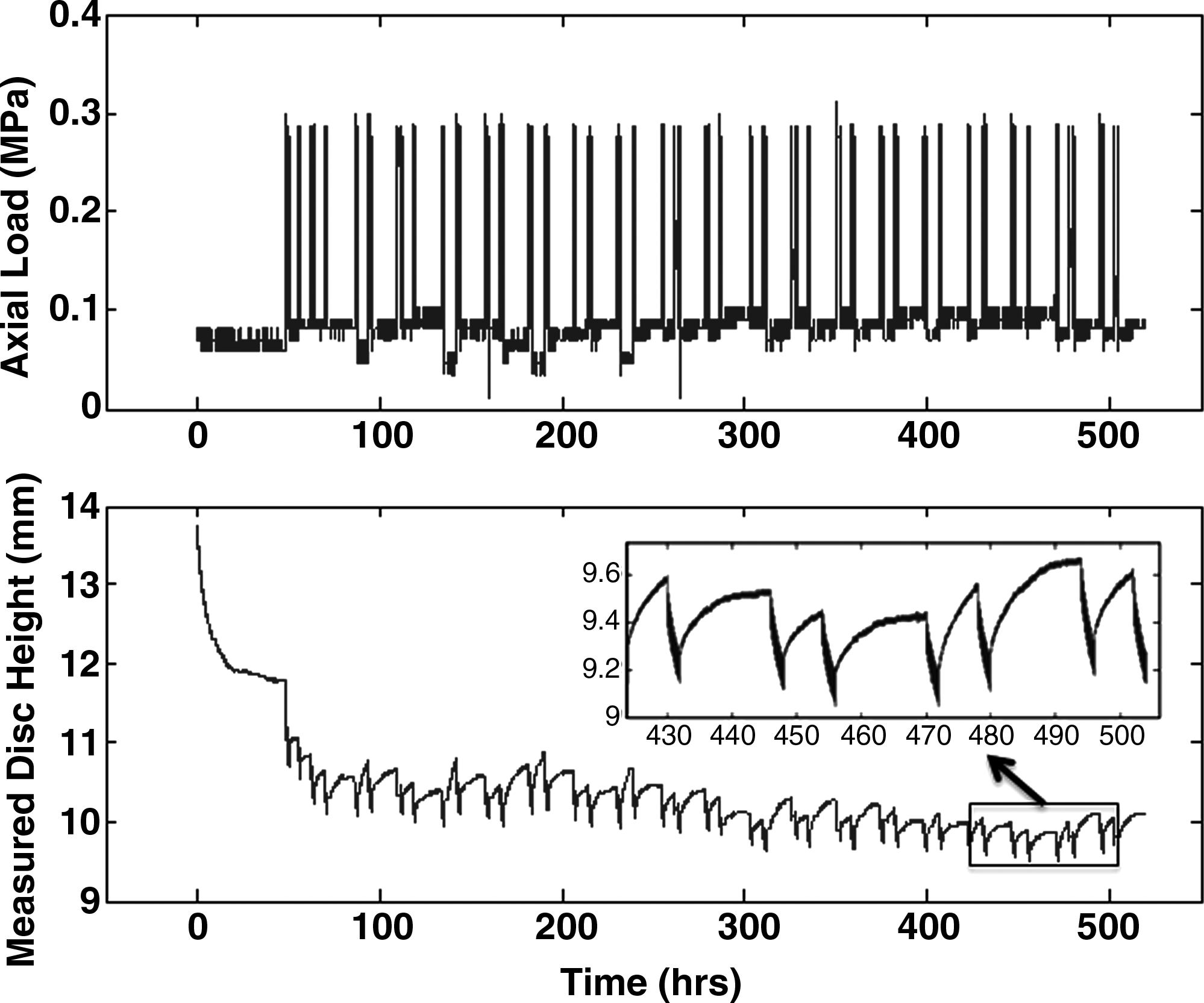
Axial load and disc height while loaded in the culture chamber. Axial load (above) and disc height (below) for disc loaded in the culture chamber (day 8 to 28). After 48 h of 0.1 MPa static loading, the disc was loaded with alternating cyclic and static load. The detailed view in the lower graph shows short cyclic loading periods with height loss, followed by longer recovery periods with the disc swelling back to its original height.
Cell viability and matrix maintenance under physiological loading is a necessity for evaluating the effect of load or the combination of load and potential bioactive candidates on the ability to induce disc regeneration. As both loading and nutrient supply have the potential to adversely affect cell viability, the proportion of live to dead cells was analyzed in the center of the NP and AF as indicated in Figure 6A. A viability of 88.4%±3.9% was observed in the NP and 80.8%±8.9% in the AF after a total of 4 weeks in culture (Fig. 6B, C). This difference was not considered significant (p=0.12). Aggrecan is susceptible to both aggrecanse and matrix metalloproteinase fragmentation, and is most likely the first molecule to show signs of proteolytic activity if induced in the discs by culturing and loading. Aggrecan fragmentation was therefore compared in freshly isolated and in discs cultured for 4 weeks un-loaded or under low cyclic loading. No significant increase in fragmentation was found as a result of this low loading scheme compared to freshly isolated tissue. In contrast, an increase of aggregan fragmentation was observed in discs cultured un-loaded (Fig. 6D).
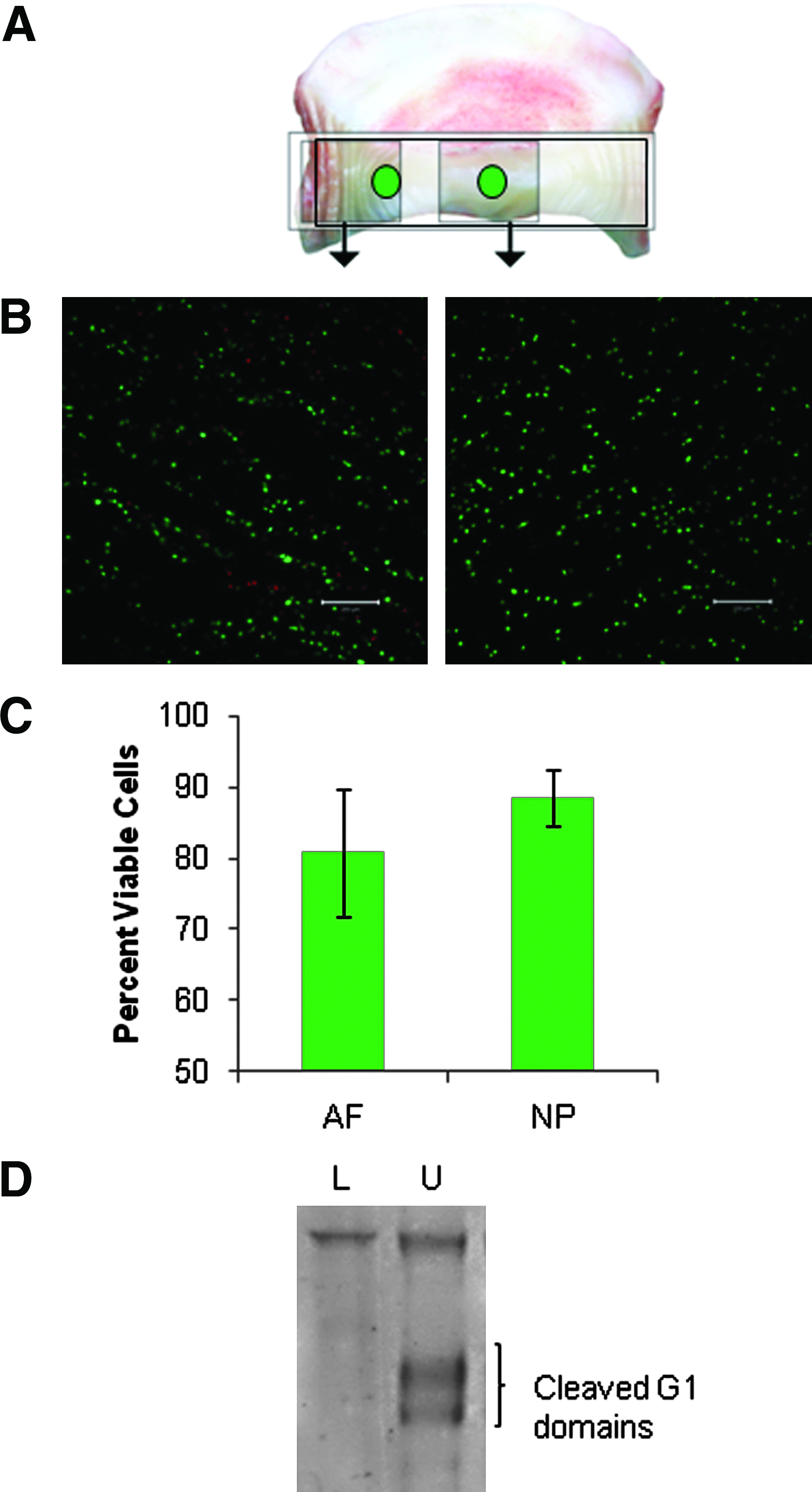
Cell viability in cultured CEP discs.
To validate that the system can be used to study the effect of load, three different loading schemes were followed: low (0.1–0.3 MPa), medium (0.1–0.6 MPa), and high (0.1–1.2 MPa). Cell viability and matrix degradation were studied after 10 days of dynamic loading. Medium and high loads resulted in decreased cell viability; the recorded values were 81.7%±7.7% in unloaded group, 93.2%±3.4% in the low load group, 36.0%±15.1% in the medium load group, and 29.3%±16.2% in the high load group after 10 days (Fig. 7A). In addition, aggrecan fragmentation was increased in the medium and high loaded discs, but fragmentation was unaffected in the low or un-loaded discs after 10 days compared to freshly isolated tissue (Fig. 7B).
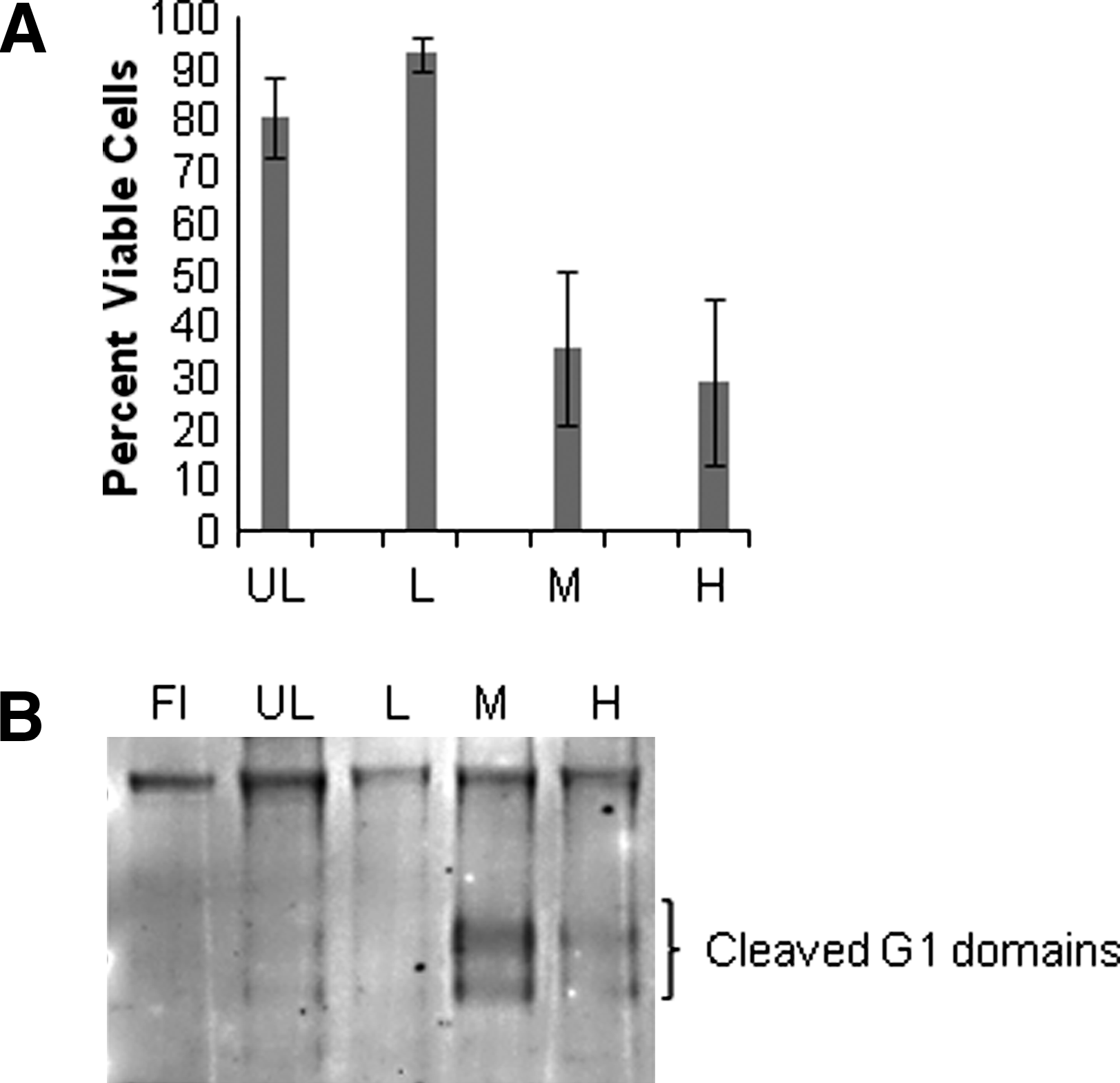
Discussion
Using a novel disc preparation and isolation method along with an axially loading bioreactor, we were able to culture bovine caudal discs for a total of 28 days. After this time the discs exhibited excellent cell viability for the NP and AF. This is, to our knowledge, the longest reported survival period of any loaded IVD organ culture model in the literature.
A large animal disc isolation and culturing method was first described by Lee et al. 30 for bovine discs, who found that cell viability in the NP was compromised after 7 days in culture under static load. They also found that the medium did not fully penetrate the BEP due to blood clots in the endplate. They therefore suggested administrating anticoagulants to the donor before slaughter to prevent the blood clots from forming or to use postmortem treatment of the endplates to remove blood clots. In addition, Lee et al. suggested that dynamic cyclic loading could stimulate nutrient diffusion into the disc, and this could be evaluated using a bioreactor. A bioreactor has been used by Gantenbein et al., 29 who exposed whole ovine caudal IVDs with vertebral endplates to continuous uniaxial diurnal loading for 7 days. Illien-Junger et al. 36 used the same bioreactor system to investigate the effects of limiting nutrition to the IVDs under high-frequency loading for 21 days. However, the culture chamber of this bioreactor cannot house larger bovine or human discs. Also, there is a need to always treat the animals with heparin before slaughter.
The culture of discs with parallel transversal cuts near the endplates has been described by Korecki et al., 31 for the culture of bovine caudal discs with static and diurnal loads. Cell viability was reported for up to 8 days of culture. Even though this method may have few limitations for nutrient flow to the disc, there is disruption of the disc integrity and altered mechanical function as a consequence of the complete removal of the endplates. Therefore, this isolation method may be unsuitable for long-term organ culture to evaluate the effect of mechanical load on the disc's biological response.
Our novel disc isolation and preparation method, whereby the BEP and the adjacent calcified part of the cartilaginous endplates are removed, allows both ample nutrient flow to enter the disc and maintenance of disc integrity. In fact, human and bovine discs have already successfully been cultured using this technique, but without external load application, for up to 4 months.32,37
However, one could argue that the novel preparation method may still disrupt the disc's ability to withstand axial loads. We therefore measured disc height throughout the culture period. The height initially decreased under intermittent cyclic loading, as can be expected after a period of free swelling, but equilibrates after <1 week. Intermittent periods of short cyclic loads, interrupted by longer but lower static loads, allow the disc tissue to keep swelling back to its initial height. Final disc heights, compared to the height at the time of harvest, were on average reduced by only 8%. Under 3 weeks loading the discs exhibited on average a 4% increase in radial diameter, and a 3% increase in weight. In light of disc bulging and diurnal water shifts being largely physiological, the above changes seem negligible. The discs therefore do appear to be mechanically competent in withstanding axial loads over prolonged periods of time.
To evaluate the effect of increasing load, low, medium and high loads were applied for 10 days. Cell viability decreased and aggrecan fragmentation increased in response to medium and high loads, whereas both cell viability and aggrecan fragmentation were unaffected by low loads. Interestingly, aggrecan fragmentation was more pronounced in the medium compared to the high load group. Low aggrecan fragmentation could be because the cells were rapidly killed in response to the high load, and therefore did not have time to produce and activate proteinases to the extent of the medium load group. Alternatively, it is possible that G1 fragments were generated, but that high loads also result in fragmentation of hyaluronic acid so releasing the bound G1 domains into the culture medium. This explanation is, however, less likely, as diffusion from the disc is a slow process. It is also notable that a load range that would be considered as physiological in vivo in a human disc (0.1–0.6 MPa) appears to be nonphysiological in the bovine tail disc. The bovine tail disc is normally not exposed to this level of axial compression and may therefore be more sensitive to load.
We believe that the bovine caudal model has several advantages over all previously presented models. The discs are inexpensive, and readily available in large numbers. Native caudal discs also have a reasonable size (on average 28.0 mm in diameter and 10.0 mm in height), and thereby nutrient transport distances are similar to those of human lumbar discs. Bovine caudal discs are much taller and larger in diameter than ovine lumbar discs.
Finally, the bioreactor used in this work to axially load isolated discs is the result of several years development and incremental improvements. Initial experiments were plagued with early bacterial and yeast infections, and medium flow was not optimally reaching the loaded disc. Rigorous standardization of the tissue preparation techniques, optimal choice of materials, simple assembly, disassembly and cleaning procedures of the culture chamber, and a robust loading frame unit have ultimately yielded a stable experimental platform. The bioreactor culture system is ideally situated for studying IVDs in an in situ environment. This system was designed to study the effect of rather slow loading frequencies and so far with only 0.1 Hz used. The responsiveness of the actuator may, however, not be adequate to test high frequencies or impact loading. The culture system provides an experimental platform where numerous parameters can be evaluated, for example, different loading schemes, O2 tension, glucose levels, and the effect of growth factors. The bioreactor can also be used to evaluate changes in mechanical properties of the disc after biologically induced changes, or use biomechanical stimulus of the disc to generate a biological change. As such, it is equally useful for studying the role of load in inducing disc degeneration or the role of biological stimuli in restoring disc function.
Footnotes
Acknowledgments
Scott McGrail, M.Sc., for his skilful laboratory bench work, including the realization of experimental setup and technical supervision. Julien Bergeron for his laboratory work and skills in conducting the stress profilometry measurements. Donal McNally, Ph.D., for supplying the ready-to-use stress profilometry measurement set and personal advice on how to use it.
This work was funded by AOSpine grant number AOSBR-07-07, the Canadian Arthritis Network, and the Shriners of North America.
Disclosure Statement
No competing financial interests exist.