
Abstract
We verified the proendocrine effects of Matrigel overlay in an adult mouse pancreatic ductal epithelial cells (PDEC) model and then decomposed the environment to delineate the specific factors responsible for this effect. Following overlay with Matrigel, supplementation of Matrigel to the culture medium, or suspension within Matrigel, neurogenin3-infected mouse PDEC underwent dramatic morphogenesis, transitioning from a two-dimensional monolayer to three-dimensional (3D) cysts. Along with these morphogenic changes, the cells displayed up to approximately sixfold increase in mRNA for the islet hormones somatostatin and ghrelin. Following overlay with collagen or suspension within collagen, PDEC also displayed similar morphogenic changes, but a much smaller increase in expression was observed (1.5- to 3-fold), suggesting that while 3D morphogenesis is capable of independently enhancing islet differentiation, biochemical factors present within Matrigel also have proendocrine effects. Following suspension within laminin gels, PDEC formed 3D cysts and also displayed an increase in islet hormone expression, similar to those cultured within Matrigel. However, medium supplemented with laminin failed to promote 3D morphogenesis of PDEC or enhance islet hormone expression, suggesting that while laminin is capable of enhancing islet hormone expression, 3D morphogenesis is required for this effect. Cell clustering appeared to maximize differentiation, as PDEC cultured on Matrigel formed aggregates and stimulated the highest expression of somatostatin and ghrelin (up to ∼200-fold).
Introduction
In an effort to address the anticipated demand for additional islets, several cell types have been investigated in vitro as potential progenitors for the generation of islet cells in vitro, including embryonic stem cells,4–9 liver stem cells,10,11 adult pancreatic nestin-positive cells,12–14 pancreatic ductal epithelial cells (PDEC),3,15–17 and clonal cultures derived from single cells isolated from the pancreas.18,19 While each of these cell types has shown promise, the efficiency of differentiation toward islet cells has been very low, and a better understanding of the factors and mechanisms that regulate differentiation from islet precursors is necessary. Broadly speaking, two classes of extracellular signals likely cooperate to regulate differentiation of precursors to islet cells: (1) biochemical signals transmitted through soluble factors (e.g., growth factors and hormones) and (2) biochemical and physical signals transmitted through extracellular matrix (ECM) proteins. While soluble factors may be delivered from various locations throughout the body, the basement membrane proteins in direct contact with the pancreatic epithelium are fairly well defined. 20 These factors can influence cell shape and polarity, and transmit signals that affect gene expression. 21
As islet precursors appear to bud from duct-like structures during both fetal development22,23 and adult pancreatic regeneration,24–26 the focus of research3,15–17,27,28 has been the transdifferentiation of PDEC to islet cells. In one of the earliest demonstrations of the feasibility of this approach, cultures enriched for human PDEC were coated with Matrigel, a commercially available basement membrane–rich gel, which stimulated the formation of three-dimensional (3D) cysts with protruding, multicellular islet-like buds. 3 Using this protocol, it was estimated that up to 32,000 islet equivalents could be generated from a single-donor pancreas, a significant accomplishment, but only about 4% of the islets required for a single human islet transplant.
Interestingly, Matrigel has also been used to stimulate or maintain differentiation of several other cell types.29–35 For example, when hepatocytes were plated on or overlaid with Matrigel, they assumed a rounded cell shape and maintained differentiated function over extended periods (e.g., elevated gene expression and secretion of albumin, transferrin, and fibrinogen).34,36,37 This effect was mainly attributed to cell rounding that was promoted by Matrigel rather than a particular biochemical signal generated by an ECM ligand.38,39 As with hepatocytes, Matrigel-mediated differentiation of mammary epithelial cells (MEC) toward the lactogenic phenotype also was dependent on cell rounding and clustering. However, biochemical signals, specifically those transmitted by laminin, were also necessary for differentiation. 40
As Matrigel is a complex tumor-derived gel consisting of a variety of growth factors and ECM factors that could potentially stimulate differentiation through a variety of mechanisms, it is unclear what the primary stimulus for islet differentiation following overlay of PDEC was. We found it particularly interesting that islet differentiation was associated with cyst formation and/or cell clustering in the Matrigel overlay islet differentiation model3,41 and others.3,7,8,10,15,42 Our own experiments with human PDEC showed that induced cell clustering enhances islet differentiation independent of exogenously added ECM, 27 but the magnitude of this effect appeared minor when compared to the effects of Matrigel overlay previously reported, 3 suggesting to us that biochemical signals (i.e., growth and/or ECM factors) presented by Matrigel also play a role in the stimulation of islet differentiation.
Here we utilized a mouse (mPDEC) model developed in our laboratory 43 to investigate which specific biochemical components of Matrigel might promote endocrine differentiation, whether the biochemical signals have an additive or synergistic effect with morphogenesis, and whether the nature of morphogenesis (i.e., cell clustering vs. cyst formation) is important to islet differentiation. When infected with retroviral vectors coding for neurogenin-3 (Ngn3), a transcriptional factor implicated in islet cell specification in vivo, cultured mPDEC express a number of markers of islet cells, including the islet hormones somatostatin and ghrelin, 28 which are characteristic of pancreatic delta and epsilon cells, respectively. Ectopic expression of Ngn3 also stimulates cultured human PDEC to express endocrine markers, including those for β cell phenotype, 16 suggesting that mPDEC ectopically expressing Ngn3 represent a convenient model for studying factors regulating the efficiency of islet cell differentiation despite the fact that they differentiate to islet cell types other than β cells.
Materials and Methods
Isolation, growth, and retroviral infection of mPDEC
Pancreatic duct fragments were isolated from 3- to 4-month-old wild-type B6SJLF1 mice or 15-month Ngn3-EGFP ± knock-in mice 44 and cultured on thick collagen gels in six-well plates as previously described to generate monolayer outgrowths of mPDEC. 43 To initiate islet differentiation, passage 6–7 mPDEC were infected with Ngn3 retrovirus, with LacZ retrovirus used for controls. Infection was conducted by exposing subconfluent cultures of mPDEC retrovirus containing 8 μg/mL polybrene (Sigma, St. Louis, MO) for 8–14 h at 32°C. Details of the virus production and infection were previously described. 28
Exposure of mPDEC to various ECM environments
The experimental design is illustrated in Figure 1. For ECM gel overlay experiments, subconfluent mPDEC on thick collagen were coated with a thick layer (1.2 mL/well) of collagen (2.3 mg/mL) or Matrigel (8 mg/mL; BD Biosciences, Bedford, MA) at 2 days after infection. For experiments involving exposure to soluble ECM, mPDEC were fed with normal medium supplemented with 62.5 μg/mL soluble laminin (BD Biosciences) or 112 μg/mL Matrigel (containing 56% laminin, or ∼62.5 μg/mL) starting at 2 days after infection. For the remaining experiments, mPDEC were passaged from thick collagen gels onto nontreated plastic (NTP; BD Biosciences), fresh collagen gels, or Matrigel, or encapsulated within fresh collagen gels, Matrigel, or high-concentration gel-formining laminin (4.5 mg/mL, containing 16% entactin; BD Biosciences). Passage of mPDEC has been previously described. 28 Both laminin and Matrigel were derived from the Engelbreth-Holm-Swarm (EHS) tumor; the major laminin family member produced by the EHS tumor is laminin-1, although trace amounts of laminin-5 can be detected. Soluble laminin is 90% pure but contains 3% entactin, a basement membrane cross-linker, while Matrigel contains 8% entactin. For all experiments, gene expression was analyzed at 10 days after infection of wild-type mPDEC or mPDEC isolated from Ngn3-EGFP +/− mouse pancreata.
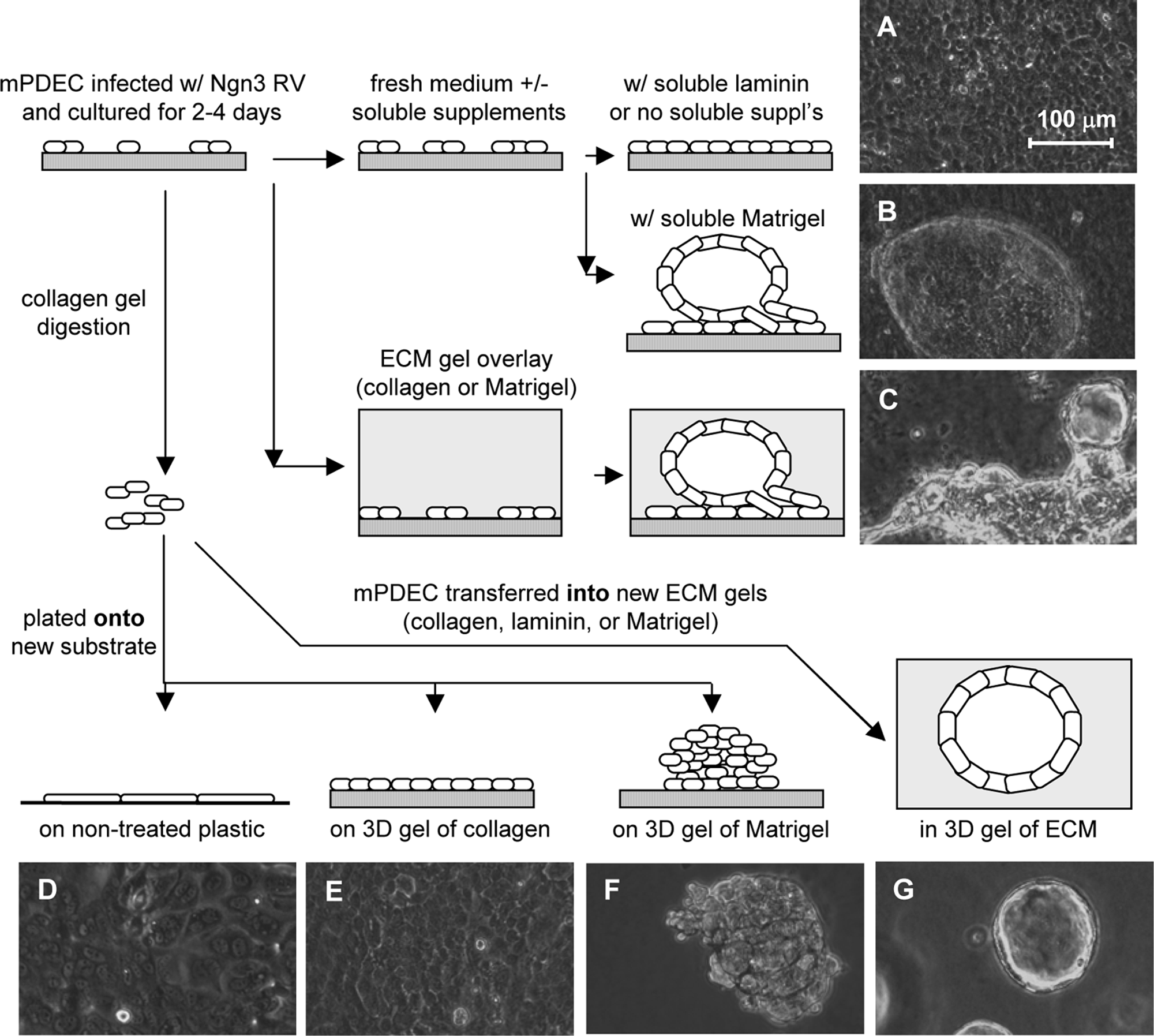
Illustration of experimental design and photographs of mPDEC morphology in various ECM environments (
RNA isolation and quantitative RT-PCR
mPDEC were recovered from collagen by digesting the gels with collagenase type V (1 mg/mL; Sigma) and from Matrigel and laminin with dispase (2 U/mL; Invitrogen, Carlsbad, CA) at 37°C on an orbital shaker for ∼20 min. Total RNA was isolated as described previously. 28 As it was impossible to obtain single-cell suspensions without compromising the integrity of mPDEC, cell yields under the various culture conditions were estimated using total RNA yields.
Quantitative RT-PCR was performed using the FastStart DNA Master SYBR Green I kit and LightCycler system (Roche Applied Science, Indianapolis, IN) using primers and conditions previously described. 28
Statistical analysis
For mouse experiments, RNA was isolated separately from replicate wells. From three to nine replicates were analyzed per condition (replicate number for each condition is detailed in the figure legends), and quantitative RT-PCR analysis was run separately on each sample. Gene expression levels were averaged for each particular treatment and reported as mean ± standard deviation (SD). As residuals appeared proportional to the means, data were log transformed to assure equal variance prior to statistical analysis using the Tukey–Kramer honestly significant difference (HSD) test (α < 0.05).
Results
Ngn3 stimulates somatostatin and ghrelin expression in a dose-dependent manner
When primary mPDEC were cultured on thick collagen gels, they expressed typical ductal markers (e.g., CA-II, CFTR, and CK19), 43 while islet markers were barely detectable. 28 Following infection with LacZ retrovirus, a negative control, no change in gene expression was observed, but following infection with Ngn3 retrovirus, a small percentage of mPDEC transdifferentiated toward islet cells, with somatostatin- and ghrelin-expressing cells primarily produced. 28 Somatostatin and ghrelin transcript levels were directly dependent on Ngn3 expression levels (Fig. 2).
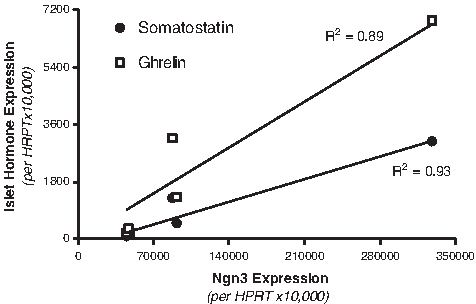
Somatostatin and ghrelin expression levels correlate highly with Ngn3 expression levels. Infection of mPDEC with ultracentrifuged retrovirus (50×- to 1×-volume concentration) resulted in elevated levels of Ngn3 expression.
ECM overlay stimulates 3D morphogenesis of mPDEC
When cultured on a thick collagen gel, mPDEC formed colonies of proliferating monolayers with typical epithelial cobblestone morphology (Fig. 3A–D). When subconfluent LacZ- or Ngn3-infected mPDEC were overlaid with a thick layer of collagen or Matrigel, colonies started to contract (Fig. 3E, I) and invade the ECM gels at the colony edges (Fig. 3F, J) within the first couple of days following ECM overlay. Over the next week, morphogenesis continued, as mPDEC formed multilayered structures (Fig. 3F, G, J, and K) and hollow cysts (Fig. 3H, L) that were somewhat similar to the in vivo tubular architecture of ducts. 45 While there appeared to be evidence of cell death (i.e., cell debris) during the first few days following ECM overlay that was most prominent in the cultures overlaid with collagen (Fig. 3E, F), the nature and extent of morphogenesis appeared similar following overlay of mPDEC with collagen and Matrigel.

Morphogenesis of mPDEC cultured on a thick collagen gel with (
Biochemical signals presented by Matrigel enhance islet differentiation from Ngn3-infected mPDEC
Matrigel overlay did not stimulate the expression of islet hormones in LacZ-infected mPDEC. However, Ngn3-infected mPDEC overlaid with Matrigel (8 mg/mL) demonstrated a significant increase in somatostatin and ghrelin expression relative to Ngn3-infected mPDEC overlaid with collagen or no ECM (Fig. 4). In particular, somatostatin and ghrelin expressions were upregulated approximately three- to fourfold following normalization to the housekeeping gene HPRT. In contrast, overlay with collagen did not increase somatostatin/HPRT or ghrelin/HPRT levels, but instead appeared to decrease the expression of these islet hormones (Fig. 4A). Surprisingly, Ngn3/HPRT levels were significantly reduced following overlay with collagen (Fig. 4B), which may account for this change. Since islet hormone expression is directly dependent on Ngn3 levels (Fig. 2), somatostatin and ghrelin expressions were normalized to Ngn3. In contrast to normalization to HPRT, a slight upregulation (1.5- to 3-fold) was apparent in the cultures overlaid with collagen following normalization to Ngn3 (Fig. 4C). The effect of Matrigel overlay was more prominent, with somatostatin/Ngn3 and ghrelin/Ngn3 levels upregulated approximately sixfold relative to mPDEC with no ECM overlay. Thus, while collagen overlay was capable of supporting morphogenesis in a similar manner as Matrigel, it did not have as potent an effect on islet hormone expression.

Levels of islet hormone and Ngn3 expression in Ngn3-infected mPDEC at 10 days after infection and 8 days following overlay with no ECM, collagen, or Matrigel. Islet hormone expression values are relative to mPDEC grown on collagen that were not treated with additional ECM supplements. (
Solubilized Matrigel, but not solubilized laminin, is capable of directing morphogenesis and enhancing islet hormone expression
As biochemical factor(s) within Matrigel appeared to present proendocrine signals to Ngn3-infected mPDEC, we explored the individual effect of laminin, the primary component of Matrigel (∼56% of total protein). At 2 days after infection with Ngn3, mPDEC were fed with normal medium (control), or medium supplemented with 62.5 μg/mL laminin or 112 μg/mL Matrigel (containing ∼62.5 μg/mL laminin). Following treatment with Matrigel-supplemented medium, mPDEC underwent morphogenesis, formed 3D cysts (Fig. 1B), and displayed a four- to fivefold increase in somatostatin and ghrelin expressions as compared to mPDEC fed with normal medium when normalized to either HPRT or Ngn3 (Fig. 5A, C). Thus, despite the absence of a solid ECM support, the observations were similar to those made following Matrigel overlay. In contrast, no changes in morphology (Fig. 1A) or islet hormone expression (Fig. 5A, C) were observed following culture in laminin-supplemented medium.
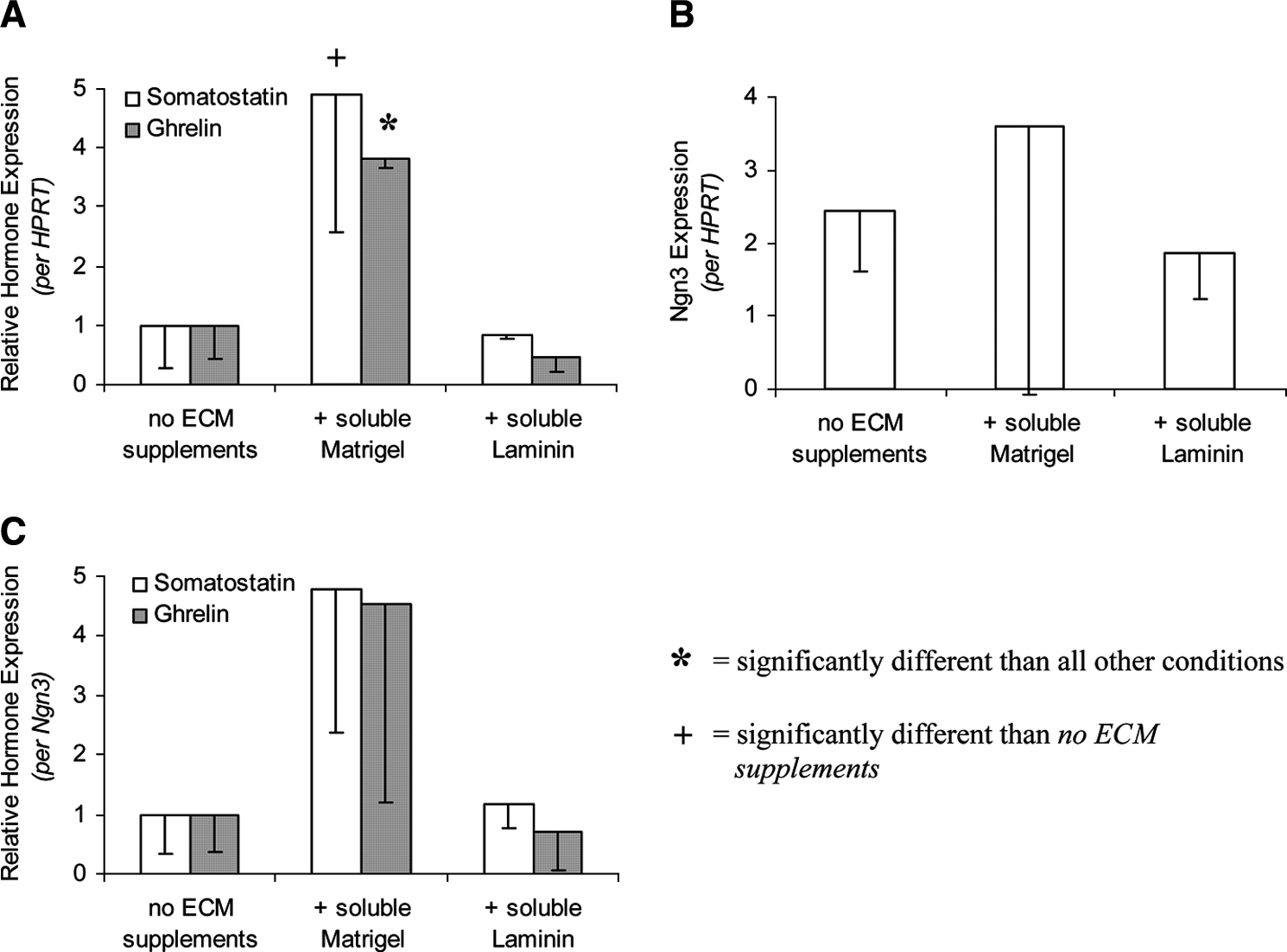
Levels of islet hormone and Ngn3 expression in Ngn3-infected mPDEC at 10 days after infection and 8 days following culture in Matrigel- or laminin-supplemented medium. Islet hormone expression levels are relative to mPDEC grown on collagen that were not treated with additional ECM supplements. (
Laminin gels support morphogenesis and enhance islet hormone expression
At 4 days after infection with Ngn3, mPDEC were passaged onto fresh collagen gels or into collagen, Matrigel, or laminin gels. When mPDEC were passaged onto collagen, as they are normally grown, mPDEC colonies attached, spread, and formed a rapidly proliferating monolayer (Fig. 1A). When suspended in collagen or Matrigel, mPDEC formed rapidly proliferating 3D cysts (Fig. 1G). Although culture of Ngn3-infected mPDEC with laminin-supplemented medium did not stimulate 3D morphogenesis, mPDEC formed rapidly proliferating cysts when cultured within laminin gels, analogous to the observations made following suspension of mPDEC in collagen and Matrigel.
Relative to mPDEC passaged onto collagen, mPDEC passaged into collagen appeared to have slightly increased islet hormone expression levels (∼1.5- and 3-fold when normalized to HPRT and Ngn3, respectively; Fig. 6A, C). However, passage of mPDEC into Matrigel had a much more potent effect, causing ∼13-fold increase in somatostatin and ∼15-fold increase in ghrelin expression when normalized to either HPRT or Ngn3 (Fig. 6A–C). Importantly, passage into laminin gels also had a strong effect on somatostatin (∼4- and 14-fold upregulation following normalization to HPRT and Ngn3, respectively) and ghrelin (∼11- and 44-fold, respectively) expression. Thus, comparing the effects of solubilized laminin with gel-forming laminin, the data suggest that laminin is capable of enhancing islet differentiation, but only when presented in an environment that promotes 3D morphogenesis.
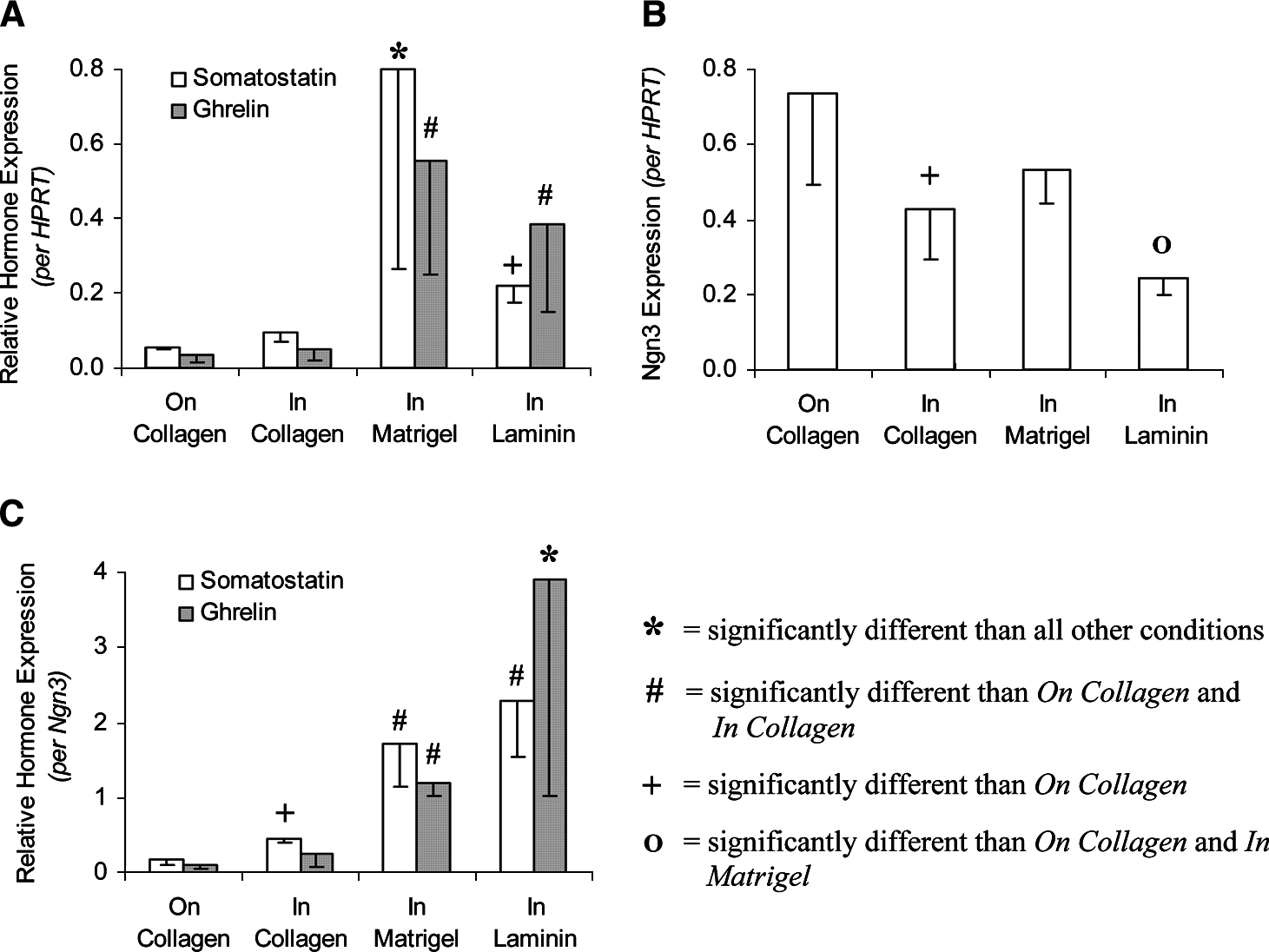
Levels of islet hormone and Ngn3 expression in Ngn3-infected mPDEC at 10 days after infection and 6 days after passage onto or into various ECM gels. Islet hormone expression levels are relative to mPDEC grown on collagen that were not passaged and not treated with additional ECM supplements. (
Matrigel-mediated increase in islet hormone expression is maximized by cell aggregation
While passage of mPDEC onto collagen led to monolayer formation and passage into ECM gels (collagen, Matrigel, and laminin) led to the formation of fluid-filled cysts, passage onto Matrigel resulted in the aggregation of cells into solid islet-like cell clusters (Fig. 1F). In particular, mPDEC colonies attached to the gel, but failed to spread and instead maintained a clustered morphology throughout the course of the experiment. Interestingly, expression of islet hormones was significantly higher following passage onto Matrigel than all other conditions (including passage into Matrigel), with >170-fold upregulation of somatostatin and ghrelin compared to mPDEC passaged onto collagen when normalized to HPRT (Fig. 7A) and >80-fold upregulation when normalized to Ngn3 (Fig. 7C). As culture of human PDEC on NTP stimulated clustering under low-serum conditions, 27 mPDEC were passaged onto NTP in an attempt to evaluate the effects of mPDEC aggregation independent of biochemical signals provided by Matrigel. However, unlike human PDEC, mPDEC formed highly spread monolayers on NTP (Fig. 1D). Further attempts to control cell aggregation through the use of poly-2-hydroxyethyl methacrylate (polyHEMA), which has been used to cluster MEC, 40 were unsuccessful, thus it is not clear from these data whether cell aggregation is adequate for the robust differentiation observed when mPDEC are cultured on Matrigel or whether additional biochemical signals are required from Matrigel.

Levels of islet hormone and Ngn3 expression in Ngn3-infected mPDEC at 10 days after infection and 6 days after passage onto or into various ECM gels. Islet hormone expression levels are relative to mPDEC grown on collagen that were not passaged and not treated with additional ECM supplements. (
Matrigel stimulates an increase in total somatostatin and ghrelin expression
While the elevated levels of somatostatin and ghrelin expression following passage onto Matrigel and into Matrigel and into laminin were dramatic in comparison to mPDEC passaged onto or into collagen (Figs. 6A, C and 7A, C), it should be noted that cell passage itself (regardless of the particular culture environment) resulted in decreased Ngn3/HPRT levels (Figs. 6B vs. 5B and Figs. 7B vs. 4B). Since there is a linear dependence between endocrine hormone expression and Ngn3 levels (Fig. 2), it was not surprising that somatostatin/HPRT and ghrelin/HPRT levels were generally decreased when comparing passaged mPDEC to nonpassaged mPDEC (Figs. 6A vs. 5A and Figs. 7A vs. 4A).
While Matrigel and laminin, when presented in an environment that supported 3D morphogenesis, clearly stimulated an increase in somatostatin and ghrelin expression when normalized to HPRT or Ngn3, we felt that it would also be important to assess whether total islet hormone expression was increased under these conditions after taking into consideration potential differences in rates of cell proliferation and death. Since cell counts could not be carried out due to difficulty in achieving single-cell suspensions without compromising the integrity of mPDEC, total RNA yields were used as a measure of total HPRT levels under the various culture conditions, which enabled us to estimate the relative levels of total Ngn3, somatostatin, and ghrelin expression (Tables 1 and 2; e.g., Total Ngn3 = Total RNA × Ngn3/HPRT). Of note, RNA yields appeared to correlate extremely well with presumed cell number, based on observations that were made during the culture period. For example, the highest RNA yields occurred following passage of mPDEC into Matrigel, which clearly stimulated the highest degree of cell expansion, while passage onto Matrigel and NTP resulted in the lowest rates of cell proliferation, and consequently, the lowest RNA yields (Tables 1 and 2). Despite a slight decrease in RNA yields and total Ngn3 expression following overlay with Matrigel (Table 1) or culture in medium containing soluble Matrigel (Table 2), total somatostatin and ghrelin expression levels still appeared to be elevated over mPDEC with no treatment. Further, while a significant decrease in somatostatin and ghrelin expression levels was generally observed following cell passage, culture on Matrigel still resulted in total islet hormone expression levels that exceeded unpassaged, control mPDEC (Table 2).
Significantly different than “on collagen” condition.
Significantly different than all other conditions.
Significantly different than “on collagen” condition.
Total Ngn3 levels = RNA yield × Ngn3/HPRT.
Total somatostatin levels = RNA yield × Som/HPRT.
Total ghrelin levels = RNA yield × Ghr/HPRT.
Discussion
Laminin-mediated increase in islet hormone expression by Ngn3-infected mouse PDEC requires 3D morphogenesis
Previously, it was demonstrated by others that Matrigel overlay stimulates the differentiation of islet cells from cultures enriched for adult human PDEC. 3 In the present study, we verified the proendocrine effects of Matrigel overlay in an adult mouse PDEC model, and then decomposed the environment to determine the responsible factors. Laminin (more specifically, laminin-1, as it is by far the most abundant laminin family member produced by the EHS tumor) was found to increase somatostatin and ghrelin expression in Ngn3-infected mPDEC in a similar manner as Matrigel, but only when presented in an environment that supported morphogenesis and formation of 3D cysts. Further, while 3D morphogenesis in the form of cyst formation was associated with enhanced islet hormone expression, cell clustering maximized the proendocrine effect provided by Matrigel.
The results of the current study build upon a previous report from our laboratory suggesting that clustering of human PDEC, independent of exogenous ECM, stimulates approximately three- to fourfold increase in islet cell differentiation. 27 Although we were unable to consistently induce mPDEC clustering in an environment devoid of laminin, our data revealed a similarly small, independent effect of 3D cyst formation on islet hormone expression. In particular, overlay of mPDEC with collagen or culture of mPDEC within collagen, devoid of exogenous laminin, led to 3D cyst formation and a slight (1.5- to 3-fold) increase in islet hormone expression when normalized to Ngn3. While it remains possible that endogenous expression of laminin by PDEC is responsible for this effect, it seems unlikely given that similar PDEC culture models show no evidence of laminin expression in vitro46,47 and that laminin-1 is not detected in adult pancreatic ducts in vivo. 20
Taken together, the data suggest that 3D morphogenesis, independent of laminin, promotes a slight increase in islet differentiation, but that the combination of 3D morphogenesis and laminin greatly enhances this effect and that 3D morphogenesis is specifically required for a laminin-mediated increase in islet hormone expression. While this is the first report that both spatial organization and biochemical signaling events contribute to the ECM-dependent regulation of islet differentiation in vitro, a similar dependence has also been reported in MEC, where cell rounding and laminin signaling were each necessary but not sufficient for differentiation of MEC toward the lactogenic phenotype, while the combination of the two resulted in the stimulation of β-casein expression. 40 Although the mechanisms by which specific 3D morphogenic changes of PDEC and MEC permit the proper translation of laminin signals are not known, several aspects of morphogenesis are potentially important. In particular, as mPDEC form 3D cysts or clusters, changes in the cytoskeleton, cell shape, and cell–cell interactions can occur, each of which could alter molecular signaling. 21
While 3D cyst formation permitted laminin-mediated upregulation of somatostatin and ghrelin expression, one of the more interesting results of the study was that the proendocrine response to Matrigel was dramatically increased following culture of mPDEC on Matrigel, which stimulated the formation of 3D cell clusters, as compared to within Matrigel, which stimulated the formation of 3D cysts. One major difference between the two conditions was that the clustered cells were more rounded than those that formed cysts, which were more flattened. As mentioned previously, there is reason to believe that these differences in cell shape could be important. In addition, as mPDEC formed clusters, cells are brought together in a more compact form that can facilitate (or result from) new cell–cell interactions. In MEC, signaling through integrins has been demonstrated to affect the formation of cell–cell adherens junctions and the establishment of cell polarity, suggesting that these pathways are intertwined. 48 While lumen-containing cysts are associated with the differentiated phenotype of MEC in vitro and are consistent with the in vivo morphology of differentiated acini, 21 cell clusters (as opposed to cysts) more closely resemble the structure of islets, which aggregate via cadherins during fetal development. 49 Consistent with our results and with a role for cell–cell interaction differentiation, restoration of the 3D configuration of primary cultures of fetal islet cells 50 and a fetal pancreatic endocrine cell line 51 resulted in a rise in endocrine hormone gene expression compared to monolayers.
Implications for tissue engineering of islets
From a tissue-engineering perspective, a reasonable goal is to maximize the expression of endocrine hormones, especially insulin. While the total expression of somatostatin and ghrelin was increased following overlay of mPDEC with Matrigel or passage of mPDEC onto Matrigel (Table 1), the passage of mPDEC decreased the expression of Ngn3. As Ngn3 levels are related to endocrine expression under a given culture condition, understanding and preventing this decrease in Ngn3 would be useful. There are at least two nonexclusive explanations for the decrease of Ngn/HPRT following passage: (1) preferential proliferation of non-Ngn3-expressing cells and (2) preferential death of Ngn3-expressing cells. Consistent with the first explanation, the decrease in Ngn3/HPRT levels was most dramatic following passage of mPDEC onto or into collagen, and into Matrigel or laminin, environments that also appeared to promote the highest degree of mPDEC proliferation, while the decrease in Ngn3/HPRT was more minor following passage of mPDEC onto Matrigel and NTP, conditions that did not appear to stimulate proliferation. Selective proliferation, however, cannot fully explain the decrease in Ngn3/HPRT levels as total Ngn3 levels decrease following overlay or passage under all conditions (Tables 1 and 2). Taken together, the data suggest that preferential death of Ngn3-expressing cells and preferential proliferation of noninfected cells both may occur, although to different extents in different culture conditions. In our work, the use of ectopic expression of Ngn3 in mouse cells is a model cell type to study PDEC to endocrine differentiation; ectopic expression of Ngn3 is not required for endocrine differentiation of human PDEC.3,27 Thus, the decrease in Ngn3 expression may not be an issue in efforts to engineer human islets.
Although the long-term tissue engineering goal is to efficiently produce insulin-producing β cells, we believe that the induction of somatostatin and ghrelin reflects a more general effect of laminin and 3D morphogenesis on islet hormone expression, and that given the appropriate cues (as discussed previously 28 ), mPDEC are capable of differentiating toward a β-cell phenotype. As both total Ngn3 expression levels (Tables 1 and 2) and Ngn3/HPRT levels (Figs. 4B vs. 7B) appeared to decrease under these conditions relative to control mPDEC grown on collagen, it is extremely unlikely that the expansion of Ngn3-expressing islet precursors is responsible for the proendocrine effect observed. A more likely possibility, which would also be consistent with the observed increases in somatostatin/Ngn3 and ghrelin/Ngn3 levels (Figs. 4C and 7C), is that 3D morphogenesis and laminin act together to increase the percentage of mPDEC that respond to Ngn3 or the efficiency of cellular response to Ngn3.
The present study, to the best of our knowledge, is the first to systematically show that laminin and 3D morphogenesis act together to enhance islet differentiation in vitro. Considering that laminin is expressed in the pancreatic ducts during fetal islet development 52 and has been reported to be upregulated during adult islet neogenesis, 53 and that morphogenesis is a hallmark of these processes, the results might also reflect an in vivo dependence of laminin-mediated islet differentiation on 3D morphogenesis. Although there is still a great deal to learn about the mechanisms by which these signals are transduced, these factors should be considered in the design of more rational protocols aimed at generating tissue-engineered islets. Of specific interest is the observation that islet differentiation is most robust under conditions that permit mPDEC to assemble into 3D cell aggregates resembling native islets. These results suggest that methods to actively promote the formation of islet-like cell clusters over cyst-like structures or monolayers would benefit the formation of tissue-engineered islets. Further analysis of the mPDEC model, including the downstream signals initiated by laminin and how cell shape, cell–cell interactions, and transcription factor expression interact with this pathway, could lead to an improved understanding of the factors controlling islet development in vivo and to the discovery of additional, more specific modifications that could optimize the production of tissue-engineered islets in vitro.
Footnotes
Acknowledgments
We would like to thank the NIH for their financial support (Diabetes and Endocrinology Research Center Pilot & Feasibility Grant #P30-DK19525, K.J.G.; RO1 #HL 64388-01A1, K.J.G.; Training Grant #HL-07954-01, M.I.B.).