
Abstract
Abstract
Neural stem and progenitor cells isolated from embryonic day 13 rat cerebral cortex were immobilized in three-dimensional type I collagen gels, and then the cell–collagen constructs were transferred to rotary wall vessel bioreactors and cultured in serum-free medium containing basic fibroblast growth factor (bFGF) combined with brain-derived neurotrophic factor for up to 10 weeks. Remarkably, the collagen-entrapped cells formed a complex two-layered structure that emulated to a certain extent the cerebral cortex of the embryonic brain in architecture and functionality. The surface layer (layer I) composed primarily of proliferating neural progenitor cells (nestin+, vimentin+, and PCNA+) predominantly expressed functional neurotransmitter receptors for cholinergic and purinergic agonists while differentiating cells (TuJ1+ and GFAP+) in the deeper layer (layer II) contained differentiated neurons and astrocytes and mainly responded to GABAergic and glutamatergic agonists and to veratridine, which activates voltage-dependent Na+ channels. An active synaptic vesicle recycling was demonstrated by neuronal networks in the deeper layer using the endocytotic marker FM1-43. Cell polarization forming the characteristic two-layered structure was found to associate with the bFGF and FGF receptor signaling. These engineered functional tissue constructs have a potential use as tissue surrogates for drug screening and detection of environmental toxins, and in neural cell replacement therapy.
Introduction
However, neural tissue engineering using the cell–collagen constructs remains a significant challenge. The cells in the constructs are typically short-lived due to the difficulty in exchanging oxygen and nutrients, which compromises cells and leads to cell death. In order to assemble single cells into highly organized, functional nerve tissue without restricted diffusion of oxygen and nutrients, simulated microgravity created by NASA-designed rotating wall vessel (RWV) bioreactors has been used in our studies. Cell culture in the simulated microgravity environment offers two beneficial factors: (1) low fluid shear stress, which promotes cell–cell contacts and initiation of differentiating cellular signaling 10 and (2) randomized gravitational vectors, which affect intracellular signal transduction and gene expression. 11 The bioreactor, which promotes aggregation and differentiation, provides an excellent in vitro system to further differentiate bioengineered tissue equivalents, such as skin, 12 cartilage, 13 cardiac cells, 14 prostate, 15 kidney, and liver fragments. 12 The bioreactor has also been used to form a rudimentary tissue-like structure from neural precursor cells. 16
The purpose of the present study is to optimize enabling technologies in achieving the first engineered living tissue construct that resembles embryonic brain tissue cytoarchitecture. We developed the stem cell–collagen–bioreactor culture system that provides the combination of naturally derived polymer scaffolds, simulated microgravity, and soluble factors that supply the physical and chemical cues to guide proliferation and differentiation of neural stem and progenitor cells isolated from the embryonic mammalian brain. In the engineered construct, neural stem and progenitor cells actively proliferate, differentiate into neurons, astrocytes, and oligodendrocytes, and finally form a complex two-layered architecture that emulates the cerebral cortex of the embryonic brain. The engineered tissue-like constructs have a potential use as animal surrogates for drug discovery and detection of environmental toxins, as well as potential use in tissue replacement therapy for cortical tissue disruption by trauma, stroke, or tumor.
Materials and Methods
Neural stem/precursor cell immobilization in collagen gel
Lyophilized collagen (Rat tail tendon, type I; Boehringer-Mannheim, Indianapolis, IN; http://www.roche.com), purchased as a sterile, lyophilized powder, was dissolved to a final concentration of 3 mg/mL by addition of sterile 0.2% v/v acetic acid (pH 3–4) (Fig. 1). Then the collagen solution was diluted with an equal volume of 2× phosphate-buffered saline (PBS; Gibco, Invitrogen, Carlsbad, CA) and a volume of cell media to achieve a final collagen concentration of 0.5 mg/mL (maintaining physiological osmolarity, 250–300 mOsM). After adjusting the pH of the collagen solution to pH 7.4 by the addition of 1 N sodium hydroxide, the solution was chilled in an ice bath to prevent gel formation. Neural stem and progenitor cells were dissociated from the embryonic day 13 (E13) rat cerebral cortex, and then added to pregelled solutions of collagen at the desired density (2 × 105/mL), along with more cell media, if necessary, to obtain a final collagen concentration of 0.4 mg/mL. About 0.4 mL aliquots of the collagen–cell suspension were placed into wells of 24-well tissue culture plates (Fig. 1). 6 The gels were placed in an incubator (37°C, 5% CO2) for 1–2 h to allow gel formation. Once the gel had set, 0.5 mL of neurobasal/B27 (NB/B27) culture medium supplemented with 10 ng/mL bFGF was added to the top of the gels and the matrix was returned to the incubator. The culture medium was changed every 3–4 days. All experiments were carried out at least in triplicate.

Schematic illustrating the in vitro system used to culture cells in collagen gels and bioreactors. Primary neural stem and progenitor cells were isolated from the embryonic rat cortical NE at the beginning of neurogenesis (E13) and immobilized in type I collagen gels. The collagen is used as a scaffold to promote cell attachment, survival, growth, and differentiation in three dimensions. The cell–collagen constructs were loaded into the NASA-designed RWV bioreactor that rotate around a central spin axis at a speed of 10–15 rpm and maintain the forming tissue in a state of continuous free fall or cultured under static conditions. The majority of cells aggregating into a disk-like gel cultured under static conditions died in 4–5 weeks, while RWV cultures promote compact aggregates into a spherical structure, which is transformed into a two-layered cortical tissue-like construct (see Figs. 5 and 6). Color images available online at www.liebertonline.com/ten.
Cell–collagen constructs are maintained in rotary cell culture bioreactors
Neural stem/progenitor cell–collagen constructs, 0.5 mL in size, weighing less than 200 mg each, were loaded into 10 mL disposable rotary cell culture system (RCCS) or RWV bioreactors (Synthecon, Houston, TX; http://www.synthecon.com) (Fig. 1), and cultured in NB/B27 medium containing 10 ng/mL bFGF. Brain-derived neurotrophic factor (BDNF) was added in the second week of culture in the RWV bioreactor. The bioreactor was placed in an incubator (37°C, humidified air, 5% CO2). As a static control, a set of cell–collagen constructs was cultured parallel in a 24-well plate. The constructs were removed from the bioreactor or culture plate at various time points for immunostaining and confocal microscopy.
Determination of cell viability in 3D collagen gel
Cell viability was determined by staining the cell construct using Live/Dead Viability/Cytotoxicity Kit (Molecular Probes, Eugene, OR; http://www.invitrogen.com). Live/dead stain was prepared with 2 μM calcein AM and 4 μM EthD-1 in PBS, and 0.2 mL of the dye solution was added to 0.4 mL gel samples along with 0.2 mL of NB/B27. The samples were incubated with the dyes for 1 h after which the excess dye solution was removed and 0.5 mL of fresh PBS was added for 30 min. For imaging, excess buffer was removed to stabilize the gel sample. Images were captured via the confocal microscopy system described below under “Confocal imaging” (Fig. 2a, b).

Comparison of cell growth and differentiation under static and low gravity conditions. (
Fluorescence immunocytochemistry
In order to identify differentiating neurons and astrocytes in 3D collagen gels, cell cultures were double-immunostained for neuron-specific β-tubulin III (TuJ1) and the astrocytic marker glial fibrillary acidic protein (GFAP). Three-dimensional cultures were fixed in 4% paraformaldehyde in PBS and incubated with a mouse monoclonal class IgG2a anti-TuJ1 antibody (1:500; Covance, Richmond, CA; http://www.covance.com) and a rabbit polyclonal anti-GFAP (1:500; Chemicon, Temecula, CA; http://www.millipore.com). Secondary antibodies were fluorescein isothiocyanate (FITC)–conjugated goat anti-mouse IgG2a (1:50; Southern Biotechnology Associates, Birmingham, AL; http://www.southernbiotech.com) and rhodamine-conjugated donkey anti-rabbit IgG (Jackson Immunological Research, West Grove, PA; http://www.jacksonimmuno.com). The distributions of TuJ1+ and GFAP+ cells were examined using a Leica confocal microscope.
To evaluate the differentiation of multipotent neural stem and progenitor cells in cell–collagen constructs, triple-immunostaining for nestin, TuJ1, and GFAP was carried out with mouse monoclonal class IgG1 anti-nestin (1:500; Chemicon), mouse monoclonal class IgG2a anti-TuJ1 antibody (1:500; Covance), and a rabbit polyclonal anti-GFAP (1:500; Chemicon). Secondary antibodies included rhodamine-conjugated goat anti-mouse IgG1, FITC-conjugated goat anti-mouse IgG2a (1:50; Southern Biotechnology Associates), and 7-amino-4-methyl coumarin-3-acetic acid (AMCA)-conjugated donkey anti-rabbit IgG (1:50; Jackson Immunological Research). Immunostained images were captured using a Leica confocal microscope.
To determine the differences in formation of pre- and postsynaptic specializations between cells cultured in static conditions and in rotary cultures, immunostaining for synapsin I (presynaptic specialization marker) and microtubule associated protein (MAP-2) (postsynaptic specialization marker) was carried out using procedures described previously. 3 Three-dimensional cell–collagen gels were rinsed in PBS and fixed with 4% formaldehyde in 0.1 M phosphate buffer (pH 7.0) for 30 min at room temperature and rinsed in PBS. Cells were exposed to a mixture of rabbit polyclonal anti-synapsin I (1:500; Chemicon) and mouse monoclonal IgG1 anti-MAP-2 (1:300; Sigma, St. Louis, MO; http://www.sigma.com) overnight at 4°C. After exposure to primary antibodies, the cells were rinsed in PBS and incubated for 45 min at room temperature with a mixture of rhodamine-conjugated donkey anti-rabbit IgG and fluorescein-conjugated goat anti-mouse IgG1 (1:50; Jackson Immunological Research). After rinses in PBS, cultures were photographed using phase-contrast and epifluorescence or confocal microscope optics (Leica TCS NT; Leica Microsystems, Heidelberg, Germany; http://www.leicamicrosystems.com). Negative controls with omission of primary antibodies were run simultaneously.
Immunohistochemistry of paraffin-embedded cell–collagen constructs
To evaluate neural progenitor cell differentiation in the cell–collagen–bioreactor system, cell–collagen constructs from various time points were fixed in 4% formaldehyde in PBS (pH 7.0; Invitrogen, Carlsbad, CA; http://www.invitrogen.com) for 10 min at room temperature, followed by thorough rinses in PBS. The constructs were dehydrated through graded alcohol concentrations (70–99.5%), followed by xylene (VWR International, West Chester, PA), and finally embedded in paraffin. Sections of 5 μm thickness were cut for qualitative staining.
Collagen-entrapped cells were examined by immunohistochemistry with well-known cell markers, including proliferation cell nuclear antigen (PCNA) for cell proliferation, nestin for neural progenitor cells, TuJ1 for neuronal progenitors and immature neurons, MAP-2 for mature neurons, synapsin I for synaptic vesicles, and GFAP for astroglial cells. Mouse on mouse (M.O.M.) Immunodetection Kit (Vector, Burlingame, CA; http://www.vectorlabs.com) was used. Target retrieval was performed on formaldehyde-fixed paraffin sections by means of 0.01 M citrate buffer (pH 6.0) in a microwave oven. The sections were then incubated with primary antibodies, which were diluted in Tris-buffered saline supplemented with 1% bovine serum albumin. We used the immunoperoxidase system (LSAB + system; Dako, Carpinteria, CA; http://www.dakousa.com; or DAB Substrate Kit; Vector), which contained biotinylated anti-rabbit, mouse, and goat Ig as a secondary antibody, streptavidin conjugated to horseradish peroxidase as a link agent, and 3,3′-diaminobenzidine (DAB) as a chromogen. Peroxidase-conjugated rabbit anti-rat IgGs were used as the secondary antibody, and DAB was used as a chromogen. After the immunostaining procedures were completed, some sections were lightly counterstained with Mayer hematoxylin (Sigma), and examined with a Zeiss microscope.
Quantitative real-time reverse transcription-polymerase chain reaction
To investigate the effects of simulated microgravity on nestin, TuJ1, and GFAP gene expressions in collagen-entrapped cells, total RNA was isolated from cell–collagen constructs generated under both culture conditions using Trizol (Invitrogen). One μg of RNA from both culture conditions was subjected to reverse transcription using Superscript II and oligo dT according to the manufacturer's protocol (Invitrogen). Samples containing 40 ng of the resulting reverse transcripts were subsequently used for quantitative real-time reverse transcription-polymerase chain reactions (RT-PCRs) using Quantitect SYBR Green PCR master mixture and designed primer pairs in a Bio-Rad iCycler thermal cycler (Hercules, CA). The resulting cDNA was subsequently used for real-time PCR reactions with a Quantitect SYBR Green PCR master mixture (Quantitect SYBR Green PCR kit) and designed primer pairs in an iCycler Thermal cycler (Bio-Rad). The reaction time, cycling conditions, and cycles were as follows: each cycle was set at 95°C for 30 s, 56°C for 30 s, and 72°C for 30 s, and run for 40 cycles. The primer pairs (from 5′ to 3′) were gtaccactggcattgtgatg and agacgcaggatggcatgag for β-actin; tgatgaggaaggagcagagtcagg and aagccatcatcccagagacc for nestin; caagctagctgtcaacatgg and ttcttggcgtcgaacatctg for TuJ1; and tgcagacctcacagacgttg and tctccaaggactcgttcgtg for GFAP. The threshold cycles (CT) were obtained for each sample and subjected to comparative CT methods using β-actin as an endogenous control for calculating the fold change according to the ABI prism 7000 user bulletin's description (Applied Biosystem, Foster City, CA; http://www.appliedbiosystems.com).
Confocal imaging
Images of stained cells were captured via confocal laser scanning microscopy using a confocal microscope (Leica TCS NT; Leica Microsystems). Collagen gel samples were placed in glass-bottomed 35-mm culture dishes for imaging. Sections of the gels, approximately 500 × 500 ×1000 μm, were scanned using a 20 × /0.40 dry plan lens. Excitation wavelengths were provided at 488 nm from an argon laser and at 568 nm from a krypton laser. The output of the confocal microscope is a series of TIFF image files representing the XY image data obtained at each Z focal plane. Combined, these images represent the volume of the gel region examined. The region of interest (ROI) in the gel sample was identified, and the system software was programmed to collect images at either 5 or 10 μm intervals through the thickness of the gel (z-dimension).
Ca2+ imaging
To characterize functional expression of neurotransmitter receptors and voltage-dependent Na+ channels in the layer I and layer II cells, Ca2+ imaging was carried out. Cell–collagen constructs cultured for 3, 7, 14, and 21 days were loaded with 2 μM fluo-3 AM (Molecular Probes) for 1 h at 37°C. The cell–collagen constructs were flattened in a 35 mm dish using a small metal frame, during the Ca2+ imaging. Cells in layers I and II were then sequentially exposed to cholinergic (carbachol), purinergic (ATP), GABAergic (GABA), and glutamategic (glutamate) agonists. Fluo-3-loaded cells were recorded using the Zeiss Attofluor Ratio Vision workstation (Atto Instruments, Rockville, MD; http://www.zeiss.com) equipped with an Axiovert 135 inverted microscope (Carl Zeiss, Thornwood, NY), an ICCD camera (Atto Instruments), and a 20 × /0.8 Plan Apochromat lens (Carl Zeiss). The fluo-3 dye was sequentially excited at 500 ms intervals using a 100 W mercury arc lamp filtered at 488 ± 5 nm, and the corresponding emissions acquired through a 510 nm dichroic mirror and 520 nm long-pass filter set (Chroma Technology, Brattleboro, VT). To collect fluo-3 fluorescence signals, ROIs were drawn electronically around individual cells in the recording field. The fluorescence intensities from up to 99 ROIs were digitized simultaneously with a Matrox image processing board and plotted as line graphs using Attograph for Windows analysis software (Atto Instruments). Changes in cytoplasmic calcium levels (Ca2+c) were monitored as changes in the relative fluorescence of fluo-3, where an increase in fluorescence intensity, measured in arbitrary fluorescence units (a.f.u.), reflected a proportional increase in Ca2+c level.
Staining with endocytotic marker FM1-43
To visualize presynaptic vesicle accumulations, cells were loaded with 10 μM styryl dye (N-3-triethylammonmpropyl)-4-(4-(dibutylamino) styryl) pyridinium dibromide (FM1-43) in depolarizing extracellular solution (50 mM K+) at 37°C. 3 The cells were rinsed with normal physiological medium without dye. Since the FM1-43 staining was performed under saturating conditions, the rate of endocytosis did not affect staining intensity. Cells were imaged with the Zeiss Attofluor Ratio Vision workstation (Atto Instruments) equipped with an Axiovert 135 inverted microscope (Carl Zeiss) and an ICCD camera (Atto Instruments). A subsequent application of depolarizing solution without FM1-43 resulted in a complete destaining of FM1-43 stained puncta.
Results
Simulated microgravity enhances long-term growth of neural progenitor cells in 3D collagen gels
Neural stem and progenitor cells were dissociated from the E13 rat cerebral cortex and immobilized into type I collagen gels. Our previous studies have shown that neural progenitor cells proliferate and differentiate along neuronal and glial lineages in the 3D collagen gels.4,6,8 However, a challenge for growing cell–collagen constructs is the formation of a necrotic core that invariably develops in the center of cell aggregates. The necrotic core contains densely packed dead cells, caused by a restricted supply of oxygen and nutrients (Fig. 2b). To overcome this difficulty, simulated microgravity was created using a NASA-designed RWV bioreactor (Fig. 1). The cell–collagen constructs were loaded into an RWV bioreactor that rotates around its central spin axis at a speed of 10–15 rpm and maintain the forming tissue in a low gravity environment state of continuous free fall. The RWV bioreactors offer low fluid shear stress, which promotes cell–cell contacts and initiation of differentiating cellular signaling, and randomized gravitational vectors, which affect intracellular signal transduction and gene expression.10–12 The cell–collagen constructs were cultured in RWV bioreactors in NB/B27 medium containing 10 ng/mL bFGF for up to 10 weeks. BDNF was added to medium at 10 ng/mL in the second week. Cell viability in 3D collagen gels was quantified by staining immobilized cells using Live/Dead Viability/Cytotoxicity Kit. As shown in the computerized 3D reconstruction of immobilized precursor cells (Fig. 2a), there appeared to be a higher percentage of live cells (green) in the RWV culture compared to the static culture. The plot in Figure 2a shows that cells cultured under static conditions manifested significant growth in the first week, continued growth over the next 2 weeks, and declined gradually in number after the third week. Cells cultured in RWV bioreactors showed a delay in cell growth in the first week and accelerated growth in second week. The enhanced cell survival and growth in rotary cultures could be maintained for up to 10 weeks.
Simulated microgravity promotes long-term differentiation of neural progenitor cells in 3D collagen gels
To evaluate the effects of simulated microgravity on the differentiation of multipotent neural stem and progenitor cells along neuronal and glial lineages, collagen-entrapped cells were examined by immunocytochemistry at different weeks in culture with well-known cell markers, including nestin for neural progenitor cells, TuJ1 for neuronal progenitors and immature neurons, and GFAP for astroglial cells (Fig. 2c, d and Fig. 3). Neural stem and progenitor cells were immobilized in type I collagen gels, and some of them were cultured in RWV bioreactors for up to 10 weeks. Consistent with results described in our previous paper, 8 the appearance of these markers represents major maturational events of neural stem cell development. In both conditions, collagen-entrapped neural progenitor cells showed active neuronal generation followed by astrocyte production (Fig. 2c, d and Fig. 3). However, cells under static conditions expressed proliferating and differentiating markers mostly in the first 3 weeks after which the majority died. In the RWV bioreactors, initially, during the first 2 weeks of culture, a lag phase of cellular differentiation was observed. Accelerated differentiation, with the cells beginning to form large aggregates without death cores, began during the third week (Fig. 2b).

Confocal microscopic characterization of distribution of neural progenitors (nestin+), neurons (TuJ1+), and astrocytes (GFAP+) in layers I and II of a cell–collagen construct. Triple-immunofluorescence staining for nestin, TuJ1, and GFAP of a cell–collagen construct cultured in an RWV bioreactor for 5 weeks. Confocal microscopic images of top (
Simulated microgravity up-regulates nestin, TuJ1, and GFAP gene expressions in 3D collagen gels
Changes in relative nestin, TuJ1, and GFAP gene expressions were compared between static and RWV cell cultures using real-time RT-PCR (Fig. 4a–c). The changes in TuJ1, GFAP, and nestin expressions relative to that of (-actin endogenous control (Fig. 4d) were quantified by fold change. Real-time RT-PCR analysis at 7, 21, and 42 days in culture showed that the expressions of genes encoding TuJ1, GFAP, and nestin were significantly lower in RWV than in static cultures on day 7. However, gene expressions were significantly higher in RWV than in static cultures at days 21 and 42 (Fig. 4a–c). These results are consistent with those from immunocytochemistry, indicating that RWV promoted TuJ1, GFAP, and nestin gene expressions in a long-term culture while a short RWV incubation period (about 1 week) revealed a lag phase in these gene expressions. 8
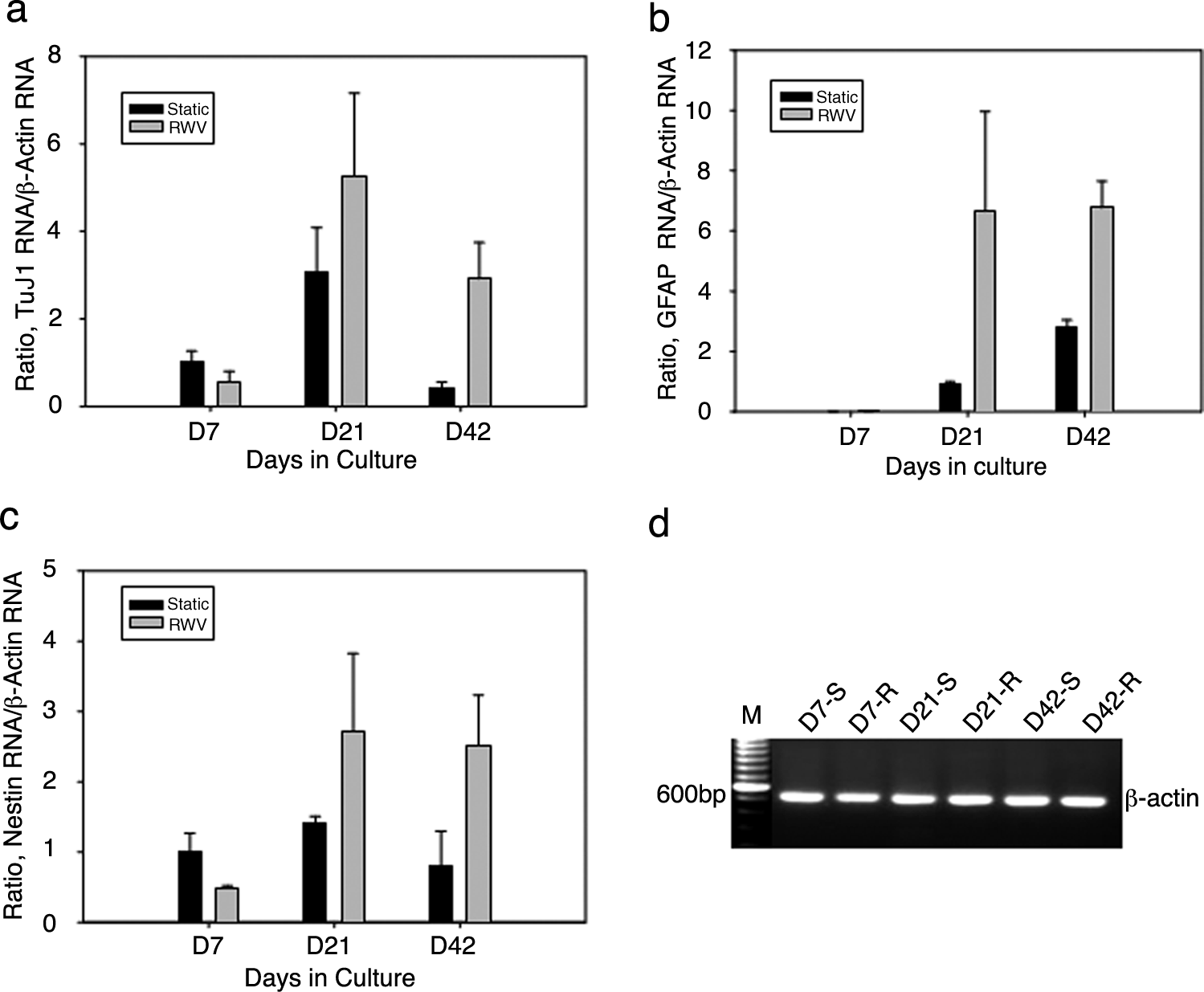
Differential expressions of nestin, TuJ1, and GFAP genes of collagen-entrapped neural stem and progenitor cells cultured under static conditions or in RWV bioreactors. Dynamic changes in nestin, TuJ1, and GFAP gene expressions relative to β-actin occurred under static and RWV cultures using real-time RT-PCR. Although the growth and cell numbers were different in both culture conditions, the same amount of total RNA (1 μg) and reverse transcripts (40 ng) were used to do real-time PCR analysis. Further, a constant gene expression level of a housekeeping gene, β-actin, served as an endogenous control for the quantitative measurements of fold changes between the two conditions among three time points using comparative CT methods (see Materials and Methods section). The constant expression level for endogenous control is essential for the quantitative measurement using comparative CT methods. Three-dimensional static and RWV cultures after 7, 21, and 42 days were examined and compared. (
Cell organization emulates native cortical tissue
As described above, cells grown under conventional static culture conditions expanded faster but degenerated more rapidly with most cells dying after 3 weeks. In contrast, collagen-entrapped neural stem and progenitor cells generated a characteristic cellular shell in rotary cultures over time (Fig. 5). The multicellular shells formed at 5 weeks and could be maintained for up to 10 weeks in rotary cultures. The cell–collagen constructs cultured in RWV bioreactors gradually compacted fourfold to fivefold and became elliptically shaped, tissue-like structures (Fig. 6a). Inspection of the tissue-like structure consistently revealed multiple layers of cells, which were populated by different phenotypes. Immunostaining of transverse sections through the construct was used to identify the different phenotypes (Fig. 6b, c). The surface layer (or layer I, interfacing with the culture medium) composing the surface of the construct consisted of a thin sheet of nestin+, vimentin+, and PCNA+ cells, which appeared to be neural progenitor cells (Fig. 6c, below the white dashed line). The deeper layer (or layer II, identified by a white arrow in Fig. 6b) was composed of a thicker, uneven, and continuous mass of TuJ1+ cells, which appeared to be differentiating neurons. GFAP+ cells were located in both layers I and II (Fig. 6c). In the center core, there was collagen matrix with small scattered aggregates of neurons (TuJ1+) and glial cells (GFAP+). Distribution of GFAP+ cells in layer I could be interesting since populations of GFAP+ astrocytes at the subventricular zone and subgranular layer of dentate gyrus are considered as potential neural stem cells in adult mammalian brain.17,18

Collagen-entrapped neural stem and progenitor cells generate a characteristic cellular shell in rotary cultures over time. H&E staining of transverse sections through cell–collagen constructs cultured in RWV bioreactors (
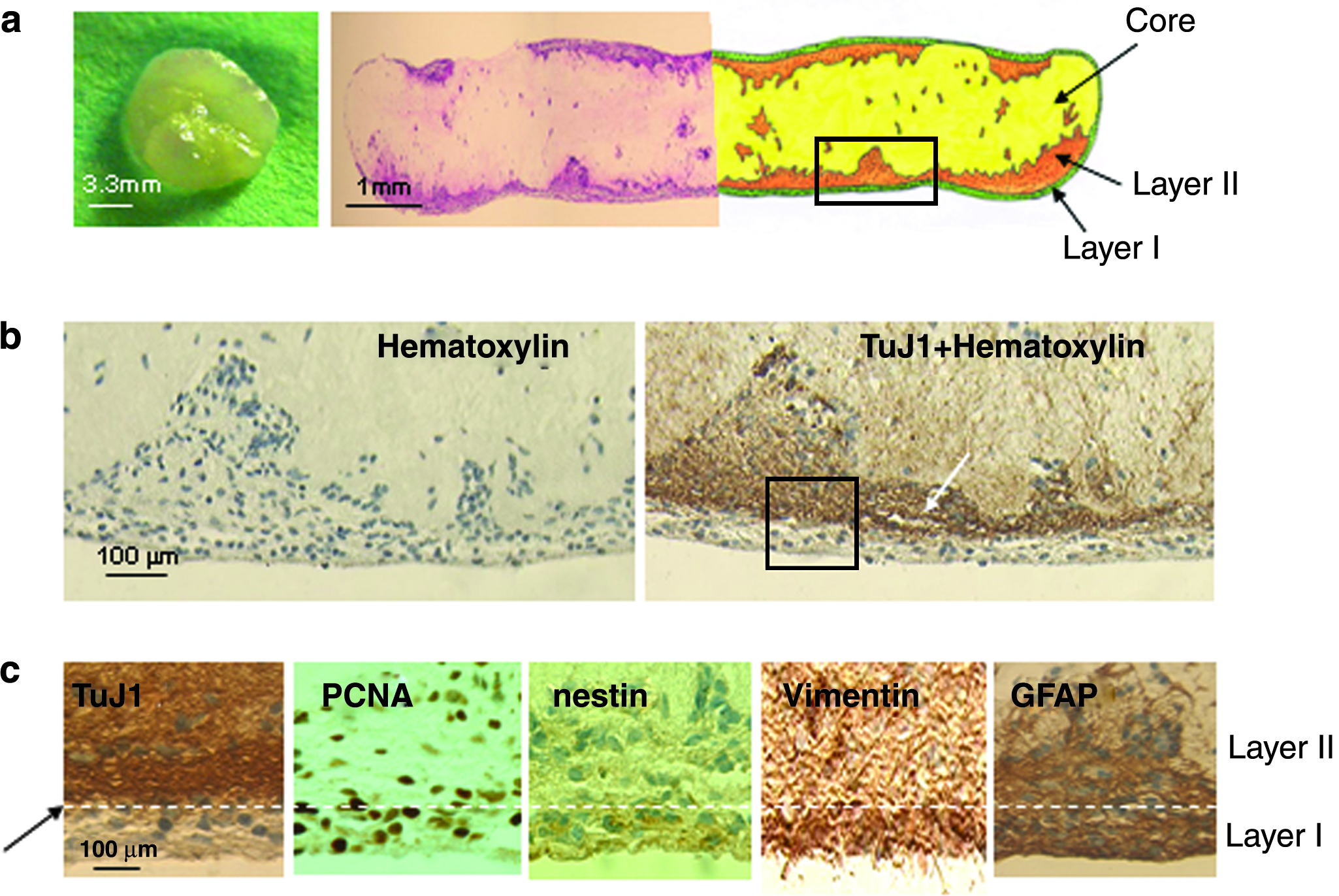
Multiepitope staining shows laminar distributions of immunoreactivities of TuJ1, PCNA, nestin, vimentin, and GFAP present in transverse sections through an intact construct generated by culturing neural stem and progenitor cells in a RWV bioreactor for 5 weeks. (
The cell organization of rotary culture–derived collagen constructs was further analyzed by triple-immunofluorescence staining and confocal microscopy (Fig. 3). Triple staining for nestin, TuJ1, and GFAP in a cell–collagen construct cultured in an RWV bioreactor for 5 weeks showed the predominant distributions of nestin+ in layer I and TuJ1+ and GFAP+ cells in layer II.
The multiple layers of different phenotypes composing the RWV-generated tissue are reminiscent of the developing rat cerebral cortex (Fig. 7). In the developing cortex of the embryonic day 19 rat (Fig. 7a), proliferating neural progenitor cells form the neuroepithelium (NE) (or ventricular zone/subventricular zone) while differentiating neuronal and glial phenotypes are found in the cortical plate/subplate (CP/SP) regions. In the RWV-generated cell–collagen constructs, layer I contained nestin+ and PCNA+ cells (Fig. 7B) corresponding to the NE, while layer II emulated the CP/SP regions. The central core may be similar to the intermediate zone (IZ), which is relatively cell poor. However, in the primary tissue the IZ lies between the NE and the CP/SP. The growth conditions imposed here may have displaced this aspect of normal cytoarchitecture. Alternatively, the central core was composed of differentiating phenotypes that have migrated away from layer II. In this case, the gels were compacted, which is usually attributed to the fact that cells pull on their local ECM, so that although the construct was polarized into a neuroepithelial sheet of neural progenitor cells (layer I) and a deeper-layer region of differentiating phenotypes (layer II), it was missing the IZ. Finally, progenitor cells in layer I, as well as some in layer II, showed an ability to synthesize fibronectin (not shown), which plays a critical role during the early central nervous system (CNS) development.

Similarity in architecture between embryonic day 19 (E19) rat cerebral cortex and RWV-generated embryonic brain-like tissue. (
Distributions of functional transmitter receptors and voltage-dependent Na+ channels, and synaptic vesicle recycling in RWV-generated cortex-like tissue constructs
We investigated the distributions of functional receptors for neurotransmitters and voltage-dependent Na+ channels among cells composing layers I and II using Ca2+ imaging. Sequential exposure to agonists at four major neurotransmitter receptors (cholinergic, purinergic, GABAergic, and glutamatergic) revealed that the vast majority of cells (86%) in layer I responded to ATP, which activates purinergic receptors (Fig. 8a). The rise in cytosolic Ca2+ (Ca2+c) to ATP was high in amplitude, but transient in duration, relaxing to the baseline during agonist exposure. About half (55%) of the layer I cells also exhibited transient Ca2+c responses to carbachol, an agonist at muscarinic cholinergic receptors. Few cells responded to the other agonists and all of these latter cells were clustered together. In contrast, few cells in layer II responded to ATP and those that did exhibited low amplitude responses (Fig. 8b). The majority (∼85%) of layer II cells responded to each of the other agonists including muscimol and glutamate, as well as to a Na+ channel activator veratridine. These differential results on cells in layers I and II parallel previously published results on proliferating precursor cells in the NE and differentiating neurons in the CP/SP. 19 Thus, the two cell layers composing the engineered tissue exhibit functional transmitter receptors and are voltage dependent similar to those developing in vivo.

Stereotypical expression of functional neurotransmitter receptors and voltage-dependent Na+ channels that regulate cytosolic Ca2+ levels among cells in the surface (layer I) and deeper (layer II) layers. Ca2+ imaging was carried out on cells located in the surface (layer I) and deeper (layer II) layers, which were sequentially exposed to 100 μM carbachol, 1 mM ATP, 10 μM muscimol, 50 μM glutamate, and 100 μM veratridine with intervening washes between each application. (
The second functional assay, using an endocytotic marker (FM1-43), showed that cells in layer II, but not those in layer I, were stained with FM1-43, which has been used to label recycling synaptic vesicles in neuronal networks. FM1-43 fluorescence intensity was reduced upon K+ stimulation (Fig. 9b), demonstrating synaptic vesicle recycling at active neuronal circuits. Pre- and postsynaptic specializations were identified by immunostaining for synapsin I and MAP-2. Synapsin I puncta were found surrounding MAP-2–positive cell bodies and dendrites in layer II and were much more abundant than those found in MAP-2+ cells derived from static cultures (Fig. 9a).
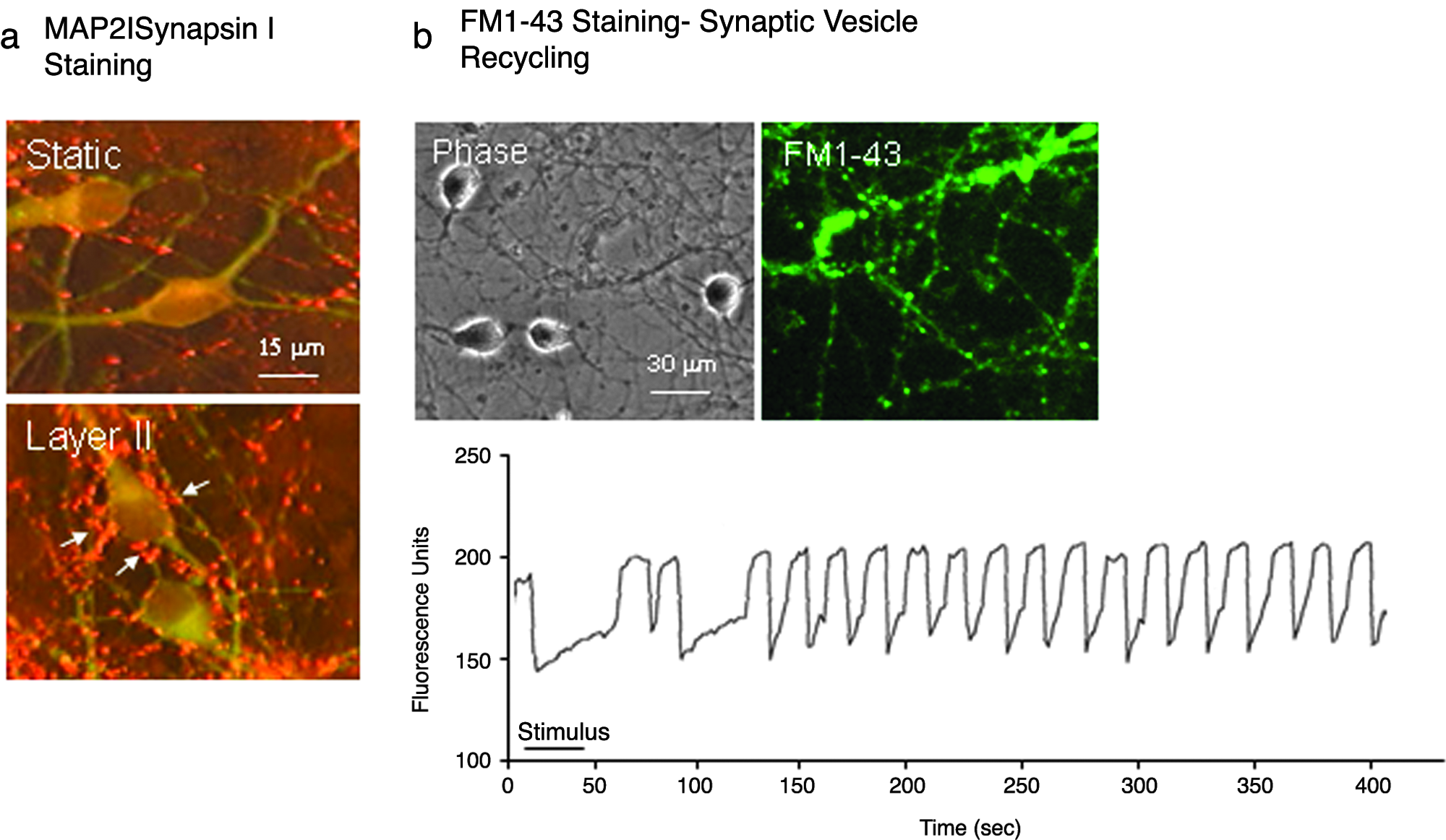
Synaptic formation and synaptic vesicle recycling of neuronal networks in layer II. (
bFGF/FGF receptor signaling is required for layer formation
The mechanisms by which the collagen-entrapped cells spontaneously polarize under microgravity conditions in the rotary culture are unknown. However, it is well known that during embryogenesis morphogenic signals are secreted by cells and morphogen gradients may organize a field of surrounding cells into patterns. 20 The morphogens can be FGFs. Increasing evidence shows that FGF family members are required for the morphogenesis of the neural folds and neural tube during embryonic development of the cortex. FGF ligands and FGF receptors (FGFRs) are expressed by neural progenitor cells from the early phases of morphogenesis.21–23 Therefore, immunocytochemistry for bFGF, FGFR1, and FGFR3 was carried out in sections cut through RWV-generated cell–collagen constructs. bFGF-like, FGFR1-like, and FGFR3-like immunoreactivities were displayed strongly in layer I cells and decreased within the thickness (Fig. 10a). Such an outside-to-inside gradient in expressions of bFGF, FGFR1, and FGFR3 was seen across the construct. The fact that the location of bFGF-, FGFR1-, and FGFR3-positive cells overlapped with that of PCNA-, nestin-, and vimentin-positive cells suggests that proliferating cells in layer I not only produce bFGF but also should respond directly to bFGF.
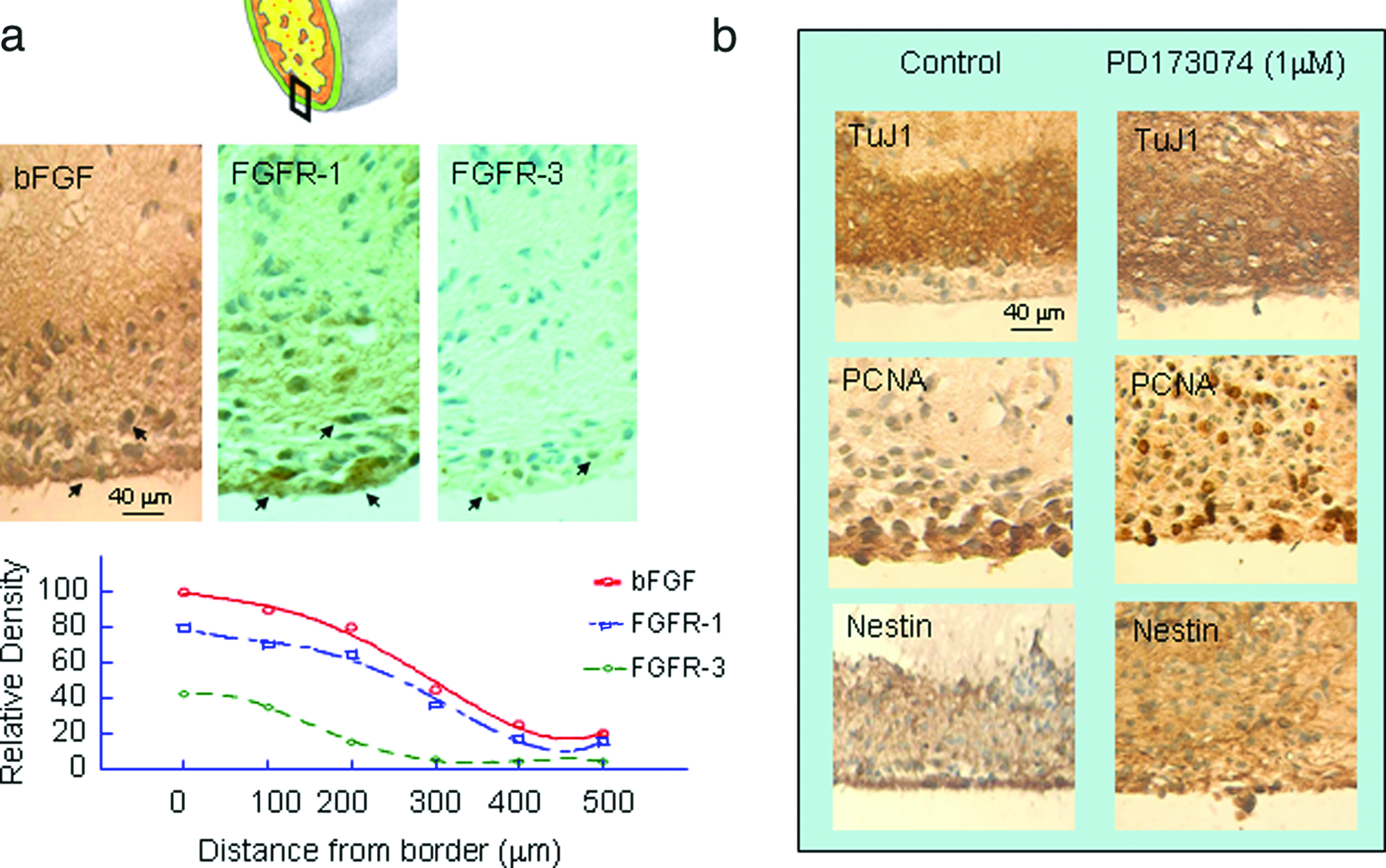
bFGF/FGFR signaling is required for layer formation in the rotary-derived cell–collagen constructs. (
To verify whether or not bFGF/FGFR signaling gradients are required for the formation of layers, a pan-FGFR antagonist, PD173074, was added to the culture medium at concentrations of 100 nM and 1 μM in the second week of culture. Immunostaining for TuJ1, PCNA, and nestin showed that the layer I formation was partially interrupted by addition of 100 nM PD173074 and completely blocked by 1 μM PD173074 (Fig. 10b). These results are consistent with our recent findings demonstrating the critical role of bFGF/FGFR signaling in the patterning of E13 rat telencephalon at the onset of cortical neurogenesis. 23
Discussion
We reported here a novel engineered construct resembling embryonic cortical tissue generated using the stem cell–collagen–bioreactor culture system. This strategy allows construction of multilayered neural tissue whose stratification resembles that of the embryonic rat cerebral cortex. There are at least three major advantages to this strategy for neural tissue engineering. First, the use of neural stem and progenitor cells, which are capable of both self-renewal and phenotypic differentiation into all types of neural cells, has the potential for supplying different cell types under controlled conditions. The neural stem and progenitor cells were isolated from E13 cerebral cortex because at E13, differentiation of the cerebral cortex just begins with the NE containing mostly neural stem and progenitor cells. Second, the use of collagen, which is the most widespread ECM protein in mammalian tissues, provides a scaffold for cell growth in three dimensions. The matrix is useful for the growth of anchorage-dependent cells like neural stem and progenitor cells into neuronal circuits and networks. Third, RWV bioreactors provide simulated microgravity that creates low fluid shear stress and randomized gravitational vectors. Compared to cultures under static, 1 g conditions, the RWV-generated tissues contain cells with persistently higher rates of survival, proliferation, and differentiation. The majority of cells aggregating into spheres in static cultures died after 3 weeks, whereas RWV cultures promoted compact aggregates, which became transformed into a multilayered tissue without a necrotic core. Thus far, the functional engineered tissue-like constructs can be maintained in vitro for up to 10 weeks.
Rotation-mediated aggregation cultures of dissociated embryonic brain cells have yielded sufficient tissue for biochemical and morphological studies. 24 The morphological differentiation observed in aggregates resembles that found in normal brain.23,25 In 26-day-old aggregates, neurons tended to occur in clusters that were located in central regions of the aggregates, whereas astrocytes were more concentrated in the periphery. Rotary cultures of bFGF- or EGF-treated mouse neural progenitor cells generated 2–3-day-old neurospheres 13 and 56-day-old aggregates, 26 in which nestin-positive cells were concentrated in the outer layer and neurons and astrocytes were more packed in core aggregates. All these rotary culture-generated aggregates formed without addition of ECM. In the present study, neural progenitor cells were immobilized into collagen scaffolds before loading into a rotary culture. Collagen is the major class of insoluble fibrous protein in the ECM. As an ECM scaffold, collagen has been widely used as a matrix for the growth of various progenitors.27–29 Since neural progenitor cells, like most mammalian cells, are anchorage dependent, the scaffolds are necessary to serve as ECM equivalents for cell survival, growth, and differentiation. 30 Therefore, neural progenitor cell proliferation, differentiation, and migration are not only under influence of growth factors but are also affected by ECMs.
A major focus of tissue engineering is how to generate functional 3D constructs that are large enough to serve as replacement organs. Three-dimensional constructs cultured in static conditions often contain hypoxic necrotic centers due to the restricted diffusion of nutrients and oxygen, which limit a construct's size and viability function. In order to attain and maintain tissue growth and differentiation in 3D assemblies, a cell–polymer–bioreactor system was used for tissue engineering of cartilage 13 and cardiac tissue. 14 In the present study, neural progenitor cells were expanded and differentiated in a microgravity environment similar to that present in space, which was simulated by a NASA-designed bioreactor. The NASA-designed bioreactor creates a reduced shear environment balanced by adequate mixing to ensure nutrient supply, removal of metabolic wastes, and factors modulating lineage differentiation. The longer survival of larger constructs with well-differentiated neuronal and glial functions demonstrated in our cortex-like tissue constructs may benefit from the use of simulated microgravity.
A fundamental challenge for tissue engineering is to understand organogenesis, the process by which cells self-assemble into tissues and then organs in vivo. Our in vitro 3D model provides a research tool to better understand the complexities of cortical development. One of the issues is to understand the effects of morphogens on cell fate determination and pattern formation during embryogenesis. 31 We demonstrated that neural progenitor cells and progeny may produce bFGF during rotary cultures and that signaling via FGFRs is required for cell survival, proliferation, differentiation, polarization, and layer formation. These are consistent with the role of FGF ligands and FGFRs during the embryonic development.23,32,33
The novel approach described here would be amenable to build different neural tissue models depending on controlled conditions. For example, a combination of bFGF, EGF, and Leukemia inhibitory factor (LIF) would be a potent synergistic stimulation of neural stem cell proliferation that generates a continuously proliferating population of immature cells. In order to generate more excitable neurons and active synaptic activity, an early withdrawal of bFGF and addition of BDNF are necessary. Further, the method described herein would be amenable to build other types of tissue models. As shown previously, a cell–polymer–bioreactor system was used to promote chondrogenesis and cartilage tissue formation. 12 This strategy could also be used to generate functional CNS tissue constructs derived from neural stem and progenitor cells from a variety of species, including humans. Thus, in the future the in vitro model could be scaled-up for clinical use, particularly for tissue replacement therapy for the injured brain and spinal cord and in neurodegenerative diseases. In addition, this method can be applied to the generation of affordable and reproducible in vitro systems with greater control of variables to serve as a possible tissue surrogate for drug screening and detection of environmental toxins.
Conclusions
We demonstrate a novel neural stem cell–collagen–bioreactor system to generate functional cortex-like tissue. This in vitro system includes (1) primary neural stem and progenitor cells isolated from the embryonic rat cortical NE at the beginning of neurogenesis, (2) type I collagen as the ECM protein to provide scaffolds for cell attachment, survival, growth, and differentiation in three dimensions, and (3) NASA-designed RWV bioreactors that offer an optimized microgravity environment. In addition, FGF signaling plays key roles in formation of the complex two-layered construct. The therapeutic implications of the engineered neural tissues and their possible use as tissue surrogate for drug screening and detection of environmental toxins remain to be explored.
Footnotes
Acknowledgments
This work was supported in part by NASA grant NRA 00-HEDS-03–136 to W. Ma. The opinions and assertions contained herein are the private ones of the authors and are not to be construed as official or reflecting the views of the Department of the Navy.