
Abstract
Angiopathies are one of the leading underlying causes of morbidity in diabetic patients. Poorly managed blood glucose levels contribute to vascular defects that manifest themselves in numerous different clinical conditions, including diabetic retinopathy, nephropathy, peripheral artery disease, and compromised wound healing. The angiopoietin family (Angs 1–4) has been shown to play a critical role in the growth and maintenance of vasculature. Here we evaluate the efficacy of a new Ang-based peptidomimetic compound, Vasculotide, on diabetic-related wound healing. Stimulation of endothelial cells (ECs) with Vasculotide results in activation of the Ang receptor, Tie 2, and its associated signaling pathways. This activation promoted biological responses such as EC survival, migration, and matrix metalloproteinase 2 (MMP2) production. We show that Vasculotide alone and in combination with vascular endothelial growth factor (VEGF) results in the production of well-arborized vessels supported by myogenic cells. Using an excisional skin–wound model produced on the back of diabetic B6.Cg-m(+/+)Lepr(db)/J (db/db) mice, we found that Vasculotide-treated wounds presented with decreased wound closure times (p < 0.05) and dramatic increases in granulation tissue (p < 0.01). Although the potential of this novel proangiogenic compound in treating microvascular dysfunction is not strictly limited to topical administration, we provide mechanistic evidence as a proof of principle in support of its efficacious use in diabetic wound healing.
Introduction
Angs are characterized structurally by an N-terminal super clustering domain (SCD) followed by a coiled-coil domain (CCD) and a C-terminal fibrinogen-like domain (FLD).3,4 Functional studies have highlighted a role for the SCD and CCDs in forming high-order homotypic Ang multimers. 5 The specific nature of these multimers is variable and seems to be unique to each Ang family member. Binding specificity of the Angs for the Tie 2 receptor has been ascribed to the FLD. Taken together, data suggests that it is the unique structural attributes of each Ang family member that promote differential clustering of Tie 2, and the pleiotropic physiological effects of Angs 1–4.5–8
Recently several short Tie 2 binding peptides were identified using a phage display screening approach. 9 Here we use the core sequence of one of those peptides and knowledge of optimal Tie 2 receptor clustering5,6 to specifically engineer a novel receptor agonist. We detail the construction of this novel proangiogenic compound, its target specificity for the Tie 2 receptor, and its use in several in vitro and in vivo angiogenesis assays. In particular, we assess the utility of Vasculotide in the treatment of excisional full-thickness wounds performed in the context of the db/db mouse strain.
Materials and Methods
Cell culture and western blotting
Pooled human umbilical vein endothelial cells (HUVECs) (Lonza, Basel, Switzerland) were grown on six-well or 10 cm plates (Nunc, Roskilde, Denmark) coated with gelatin (Sigma, Oakville, Canada) as described. 10 Cell lysate preparation and western blotting were performed as previously described.11,12
Antibodies for western blotting, immunofluorescence, and immunohistochemistry (IHC)
The following commercially available antibodies were used: monoclonal anti-Tie 2 (Pharmingen, Missisauga, Canada), polyclonal anti-pY992 Tie 2 (Cell Signalling Technology, Pickering, Canada), polyclonal anti-Mapk (Cell Signalling Technology), monoclonal anti-phospho Mapk (Cell Signalling Technology), polyclonal anti-Akt (Cell Signalling Technology), polyclonal anti-pS473 Akt (Cell Signalling Technology), monoclonal anti-pS1177 eNos (BD Biosciences, Missisauga, Canada), polyclonal anti-pY397 focal adhesion kinase (Fak) (Biosource, Burlington, Canada), polyclonal anti-Fak (Santa Cruz Biotechnology, Santa Cruz, CA), polyclonal anti-smooth muscle actin-Cy3 direct conjugate (Dako, Glostrop, Denmark), polyclonal anti-PECAM (Pharmingen), polyclonal anti-smooth muscle actin 1 (Sigma).
Peptide synthesis and biotinylation
Solid Phase Peptide Synthesis (Fmoc-SPPS) was utilized to manufacture Vasculotide peptide. Briefly, peptides were synthesized using Applied Biosystems' ABI433A Peptide Synthesizer (Foster City, CA) according to manufacturer's instructions with slight modification. Biotin was conjugated to the above T7 peptide by using EZ-link-PEO-maleimide-biotin (Pierce, Rockford, IL) according to the manufacturer's instruction.
Stimulations
HUVECs, EaHy926, 13 Cos1, 14 and C166 15 cells were stimulated in full serum containing growth media (for HUVECs: F12K media, 10% FBS, 0.1 mg/mL heparin, 0.01 ng/mL VEGF, 10 ng/mL EGF, 5 ng/mL basic fibroblast growth factor [bFGF], and 1× pen-strep) (for EaHy926, Cos1, and C166: DMEM, 10% FBS, and 1× pen-strep) for indicated times with angiopoietin 1 (R&D Systems, Minneapolis, MN), or VEGF (R&D Systems). Biotinylated Vasculotide peptide was either left unclustered or clustered with a 4:1 molar ratio of peptide:avidin in PBS for 2 h at 4°C. Stimulations with Vasculotide were performed in full serum containing growth media for 15 min unless otherwise indicated.
Boyden chamber migration assay
HUVECs were seeded at a density of 8.4 × 104 cells in 500 mL of F12 media plus 0.1% FBS in the upper chamber of an 8-μm-pore modified Boyden chamber (Falcon, Franklin Lakes, NJ), and migration assays were performed as described. 16 Student's t-test was used to test the statistical significance with a 95% confidence interval. All experiments were performed with three replicates each, and five random counts were taken per replicate.
Zymography
HUVECs cultured in F12 media plus 0.1% FBS were stimulated for 16 h with various factors as indicated. Conditioned media was centrifuged to remove cellular debris and prepared for gel electrophoresis using nonreducing conditions as described by Voskas et al. 17
Matrigel plug assay
Growth factor–reduced matrigel (BD Biosciences), impregnated with various factors, was injected subcutaneously into the flank region of 3-month-old CD1 mice (Jackson Laboratories, Bar Harbor, ME). Fourteen days postimplantation and immediately prior to sacrifice, the vascular tracing agent tomato lectin-FITC (100 μg) was injected i.v. and allowed to circulate for 10 min. Plugs were surgically resected and fixed in 4% paraformaldehyde (PFA) for 16 h. Images of vascular topology were taken using a Zeiss dissecting microscope at 4× magnification. Upon fixing, all plugs were whole mount stained with anti-Sma1-Cy3. Plugs were analyzed on a Zeiss Axiovert 100 M confocal microscope (Carl Zeiss, Toronto, Canada) for three-dimensional image reconstruction, and photos were processed using LSM Image Browser and Adobe Photoshop 7.0. Morphometric quantification of vessel characteristics was performed by skeletonizing confocal image projections with Image Processing Tool Kit 5.0 (IPTK 5.0). Student's t-test was used to test the statistical significance with a 95% confidence interval. All experiments were performed with three replicates each, with at least six representative image stacks.
Wound healing assay
Pathogen-free, 9-week-old B6.Cg-m(+/+)Lepr(db)/J (db/db) mice were purchased from Jackson Laboratories. Circular, full-excisional, 6-mm-diameter wounds were performed according to animal committee approval guidelines. Indicated factors were suspended in sterile Intrasite Topical Gel (Smith and Nephew, Hull, England) at noted concentrations. Treatments were as follows: PBS (n = 8), bFGF (R&D Systems) (10 μg/mL; n = 8), clustered Vasculotide (20 nM; n = 4), clustered Vasculotide (5 nM; n = 4). Preparations were applied at day 0 (time of wounding), days 2, 4, and 6. Standardized images were taken on days 0, 2, 4, 6, and 7 using a Canon EOS digital Rebel camera. Image J (NIH) was used to manually outline wound perimeter from which total pixel counts (wound area) were determined. Mice were sacrificed on day 7, and full wound beds including margin were excised and fixed in 4% PFA. Wounds were bisected, and each half was prepared for either paraffin or frozen sections. Wax sections were processed according to standard protocols for hematoxylin and eosin (H&E) and Masson's trichrome stain. Images were captured using a Leica DMLS compound light microscope and Pixel Link camera at 5× and 20×. Student's t-test was used to test the statistical significance.
Histological score
Histologic sections were fixed in 4% paraformaldehyde, embedded in paraffin, and sectioned and stained with H&E and Masson's trichrome for histologic evaluation. Tissue sections were evaluated by a pathologist for reepithelialization, granulation tissue formation, and inflammation (Table 1).
Adenoviral infections
Cos1 or C166 cells cultured in DMEM, 10% FBS, and 1× pen-strep were infected (multiplicity of infection [MOI] 30) overnight with adenoviruses encoding one of either EGFP or Tie 2. Infection efficiency was confirmed by epifluorescence (EGFP), or by immunoblot (Tie 2).
Cell death ELISA
HUVECs were maintained in F12 media plus 0.1% FBS for 16 h in the presence of various concentrations of unclustered or clustered Vasculotide. Analysis of apoptosis was performed using Cell Death ELISA Plus (Roche, Missisauga, Canada) according to manufacturer's specifications.
Pull-down assay
EaHy926 endothelial cell (EC) lysate prepared in Phospho Lipase Cγ lysis buffer 18 was mixed with either biotinylated Vasculotide or biotinylated irrelevant peptide. Biotinylated peptides were isolated with avidin agarose, washed three times in Phospho Lipase Cγ lysis buffer, 18 boiled in 2× SDS loading buffer, resolved by polyacrylamide gel electrophoresis (8% gel, 37.5/1 acrylamide/bisacrylamide), and subsequently tested for their ability to precipitate Tie 2 via standard immunoblot analysis.
Results
Construction of Vasculotide
T7 peptide (-His-His-His-Arg-His-Ser-Phe-) 9 was modified by addition of an amino terminal cysteine to facilitate sulfhydryl-mediated addition of a 29 Å biotinylated polyethylene glycol (PEG)–maleimide moiety (Fig. 1a). Postsynthesis clustering of this biotinylated peptide-PEG with avidin, referred to hence forth as Vasculotide, gave rise to an obligate tetrameric compound.
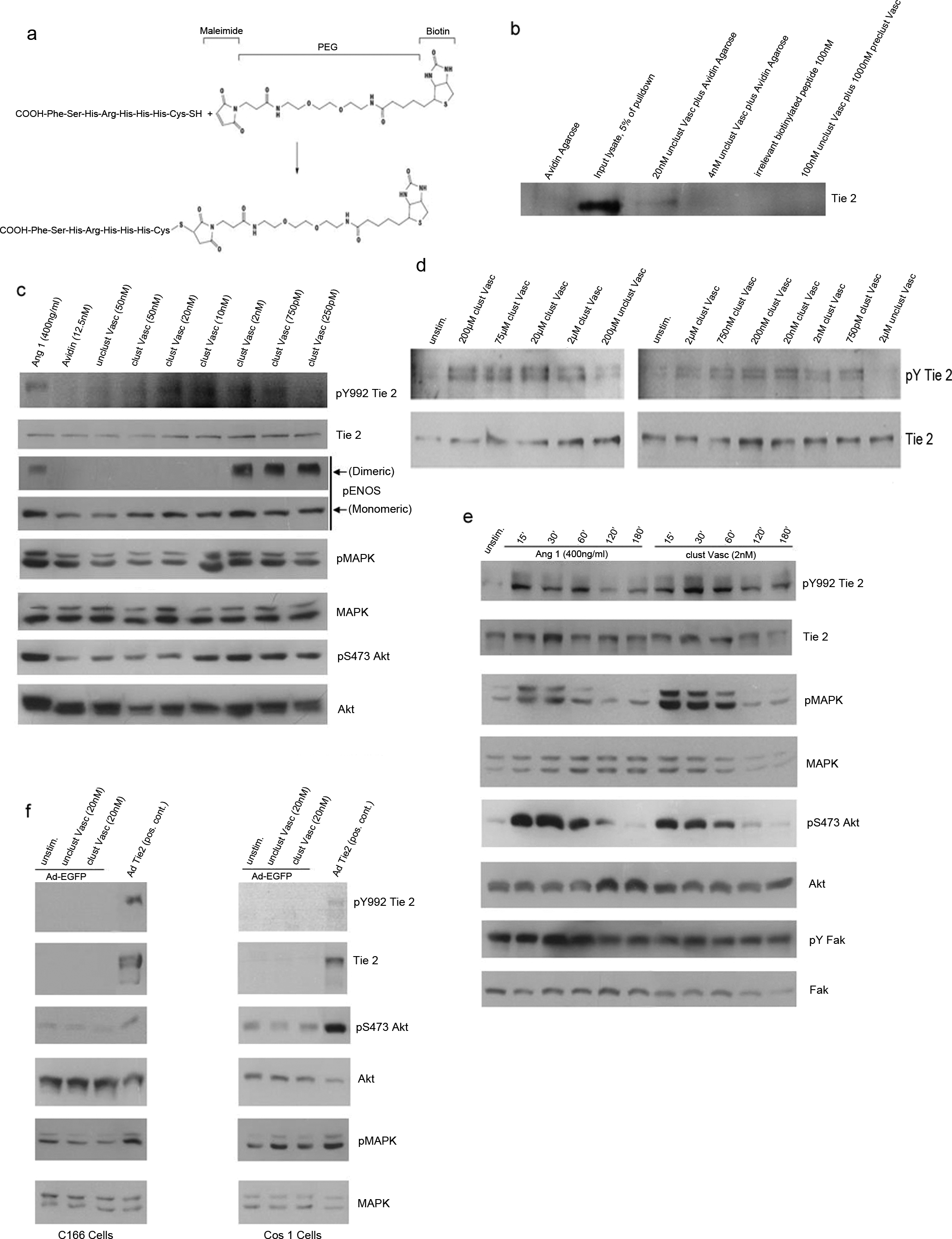
Biochemical analysis of clustered Vasculotide. (
Vasculotide binds the Tie 2 receptor
The ability of Vasculotide to bind the Tie 2 receptor, despite the engineered modifications, was tested using an in vitro pull-down assay. Nonclustered Vasculotide was mixed with whole cell lysate isolated from EaHy926 ECs. Vasculotide, but not irrelevant biotinylated peptide, was able to precipitate Tie 2 (Fig. 1b). Addition of a 10-fold excess of Vasculotide clustered with soluble avidin was able to compete for available Tie 2 and was able to abolish this interaction (final lane).
Clustered Vasculotide activates Tie 2 and associated downstream pathways
Having established that Vasculotide was able to bind Tie 2, we tested its ability, when clustered with avidin in a 4:1 ratio (Vasculotide:avidin), to activate the Tie 2 receptor and several well-established downstream signaling pathways.19–21 Immunoblot analysis of stimulated human umbilical vein endothelial cell (HUVEC) extracts revealed that clustered Vasculotide activates Tie 2 (pY992 Tie 2) at concentrations ranging from 20 nM to 750 pM, with 5–10 nM appearing optimal (Fig. 1c). Significantly, only when clustered with avidin, was vasculotide able to stimulate Tie 2 phosphorylation (see avidin alone and unclustered Vasculotide). Coincident with receptor activation, we show phosphorylation of several known Tie 2–responsive proteins including Mapk, Akt, and eNos. Paradoxically, high concentrations of clustered Vasculotide are not capable of activating Tie 2 receptor activity in HUVECs (see clustered Vasculotide 50 nM). We reasoned that this was due to the fact that high ratios of clustered Vasculotide:Tie 2 would decrease receptor clustering. This unusual dose response has also been noted with preparations of concentrated Ang1–conditioned media (unpublished observations) and by others using purified recombinant Ang1.22–27 To test this hypothesis, we utilized EaHy926 ECs which express much higher levels of Tie 2. Using this cell line we were able to demonstrate specific activation of the Tie 2 receptor at concentrations ranging from 200 μM to 750 pM (Fig. 1d), suggesting that the ratio of clustered Vasculotide:Tie 2 is critical for optimal activation.
Clustered Vasculotide displays activation kinetics similar to native Ang1
To examine Tie 2 activation kinetics, we stimulated HUVECs with native Ang1 or clustered Vasculotide for various times. Low-concentration clustered Vasculotide (2 nM) was comparable to Ang1 (400 ng/mL) stimulation in its ability to activate Tie 2 phosphorylation. Overall, activation of the receptor by either Ang1 or clustered Vasculotide followed a very similar time course, with phosphorylation returning to almost basal levels sometime after 2 h (Fig. 1e). Again we noted marked increases in pMAPK and pAkt. Previously Ang1 and 2 have been shown to activate Fak, in a Tie 2-independent manner through direct engagement of αVβ1 integrin.28,29 To test whether clustered Vasculotide was capable of activating this arm of the Ang signaling cascade, we examined the activation of Fak. As previously shown, Ang1 time-dependently activated Fak, whereas clustered Vasculotide had no significant effect. These results are suggestive of a more direct role for Vasculotide in eliciting Tie 2–specific signaling.
Clustered Vasculotide displays specificity for the Tie 2 receptor
To more formally address the specificity of clustered Vasculotide for the Tie 2 receptor, we performed reconstitution experiments in Cos1 fibroblast cells and C166 ECs, both of which are phenotypically null for Tie 2. Cells infected with recombinant adenovirus encoding EGFP or Tie 2 were either left unstimulated or stimulated with Vasculotide or clustered Vasculotide. To broadly examine the possibility that clustered Vasculotide was activating Tie 2–independent downstream signaling pathways, we examined activation of MAPK and Akt in the EGFP-infected cells. Neither Vasculotide nor clustered Vasculotide significantly induced phosphorylation of Mapk or Akt above that noted in the nonstimulated samples (Fig. 1f). As expected, overexpression of Tie 2 in either of the cell types resulted in its constitutive activation (see pY992 Tie 2) as well as activation of Mapk and Akt. This fact precluded us from stimulating these cells any further with Vasculotide or clustered Vasculotide (data not shown). Since Mapk and Akt are highly activated downstream of a host of cell surface receptors, we reason that these results are highly suggestive of a Tie 2–specific and dependent role for clustered Vasculotide in these events.
Clustered Vasculotide promotes EC survival, migration, and MMP2 release
Ang1 and, to a lesser degree, Ang2 have been shown to protect ECs from various different apoptosis-inducing conditions including serum withdrawal.30,31 Based on the fact that clustered Vasculotide strongly activates the antiapoptotic protein Akt, we wanted to determine if it could protect ECs from serum withdrawal–induced death. Analysis of cell apoptosis via cell death ELISA revealed a statistically significant decrease in serum withdrawal–induced cell death when treated with clustered Vasculotide at 500 pM (p = 0.002) compared to nontreated cells (Fig. 2a). Nonclustered Vasculotide and avidin alone had no effect on survival.
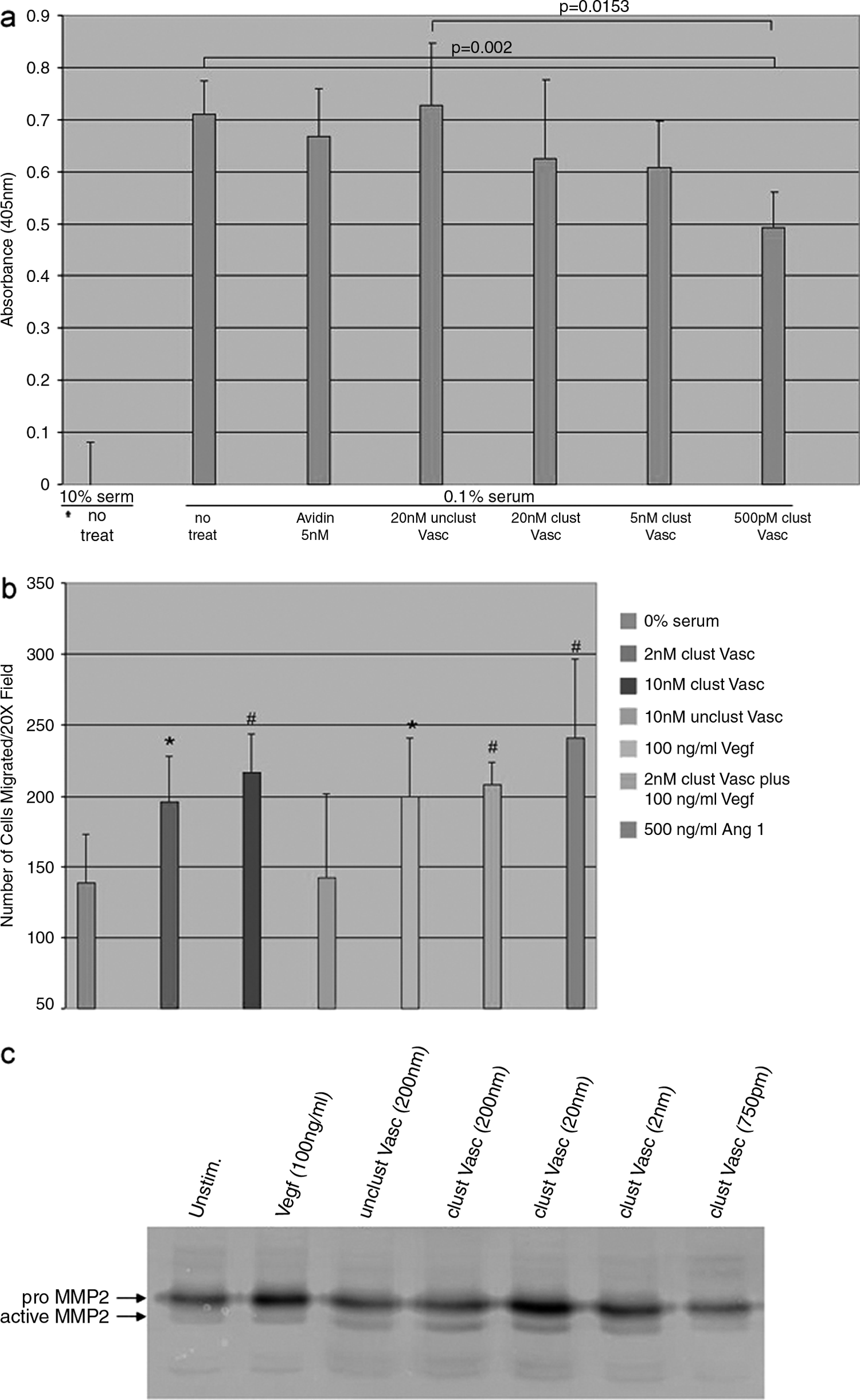
Measurement of the in vitro angiogenic activity of clustered Vasculotide. (
Previous studies underline a role for Ang1 in promoting EC migration and release of the matrix degrading enzyme matrix metalloproteinase 2 (MMP2).19,32 To test if Vasculotide, like Ang1, was capable of promoting these processes, we initially examined its influence on migration. Using a modified Boyden chamber assay, we subjected HUVECs to various chemotactic agents including Ang1, Vasculotide alone, or in combination with VEGF. Clustered Vasculotide applied at concentrations of 2 nM and 10 nM promoted significant increases in chemotactic cell migration when compared to no treatment (Fig. 2b). Noted migration was not significantly different than that seen in samples treated with VEGF or Ang1. Nonclustered Vasculotide did not promote migration above that seen in the untreated sample.
In vivo, ECs cannot migrate until they secrete enzymes necessary to break down components of the extracellular matrix. Here we employed gelatin zymography to determine if clustered Vasculotide promoted secretion of MMP2. Analysis of conditioned media revealed that cells treated with clustered Vasculotide secreted a protein that dose-dependently degraded gelatin at a molecular weight that was consistent with proenzyme and active enzyme forms of MMP2 (Fig. 2c). Taken together with the results of the migration analysis, it appears that application of clustered Vasculotide might offer distinct proangiogenic activities.
Clustered Vasculotide promotes in vivo angiogenesis
To examine the in vivo angiogenic potential of clustered Vasculotide, we employed a growth factor–reduced Matrigel plug assay. Matrigel was impregnated with either clustered Vasculotide or one of several different control factors and injected subcutaneously into adult CD1 mice. After 14 days, and prior to sacrifice, all mice were injected with FITC-lectin to facilitate vessel identification and quantification. Upon surgical removal of the plugs, a robust angiogenic response was seen in the membranous capsule that surrounded the plugs impregnated with clustered Vasculotide, VEGF, or clustered Vasculotide plus VEGF (Fig. 3a). In the case of the clustered Vasculotide samples, clear, large, well-arborized vessels were present on the surface. These vessels were well branched and contained a continuum of small and large arterioles and venules. Consistent with literature accounts, 33 vessel growth promoted by the addition of VEGF alone was apparent although the nature of these vessels was highly tortuous. Vessels located in the proximal tissue surrounding the VEGF-containing plugs also looked enlarged and inflamed upon examination (data not shown). Application of clustered Vasculotide in combination with VEGF did not seem to increase the overall vascular density, but rather contributed to decreases in the level of the tortuosity seen with VEGF alone. To more fully address and assign quantitative vessel parameters to the different treatment groups, the plugs were fixed and interrogated with laser confocal microscopy analysis combined with postprocessing image analysis (IPTK 5.0; Reindeer Graphics, Asheville, NC). No significant vessel growth was noted in plugs containing Matrigel or Matrigel impregnated with nonclustered Vasculotide (Fig. 3a–d). Plugs containing VEGF, clustered Vasculotide, or clustered Vasculotide plus VEGF displayed robust induction of angiogenesis. Statistically, VEGF, clustered Vasculotide, or combinations thereof did not differ in total vessel number, length, or branch points (Fig. 3d). As was noted in the membranous capsule surrounding the plugs, samples that contained clustered Vasculotide displayed more organized branching patterns and less tortuosity than the plugs containing VEGF alone. Ang1 has been reported to potentiate the recruitment of myogenic support cells 34 and inhibit VEGF-induced vascular permeability through mechanisms that are not yet well understood. 35 To determine if Vasculotide, like Ang1, was capable of initiating pericyte recruitment, we performed whole mount immunofluorescence against smooth muscle actin 1 (Sma1). Matrigel plug analysis of VEGF-driven angiogenesis revealed a relative absence of Sma1-positive staining cells (Fig. 3c). Samples containing clustered Vasculotide alone or in combination with VEGF displayed a significant increase in overall Sma1 staining. Sma1 staining was tightly associated with the exterior of the vessels and was enriched on larger vessels. Taken together these results demonstrate potent in vivo angiogenic effects of clustered Vasculotide.
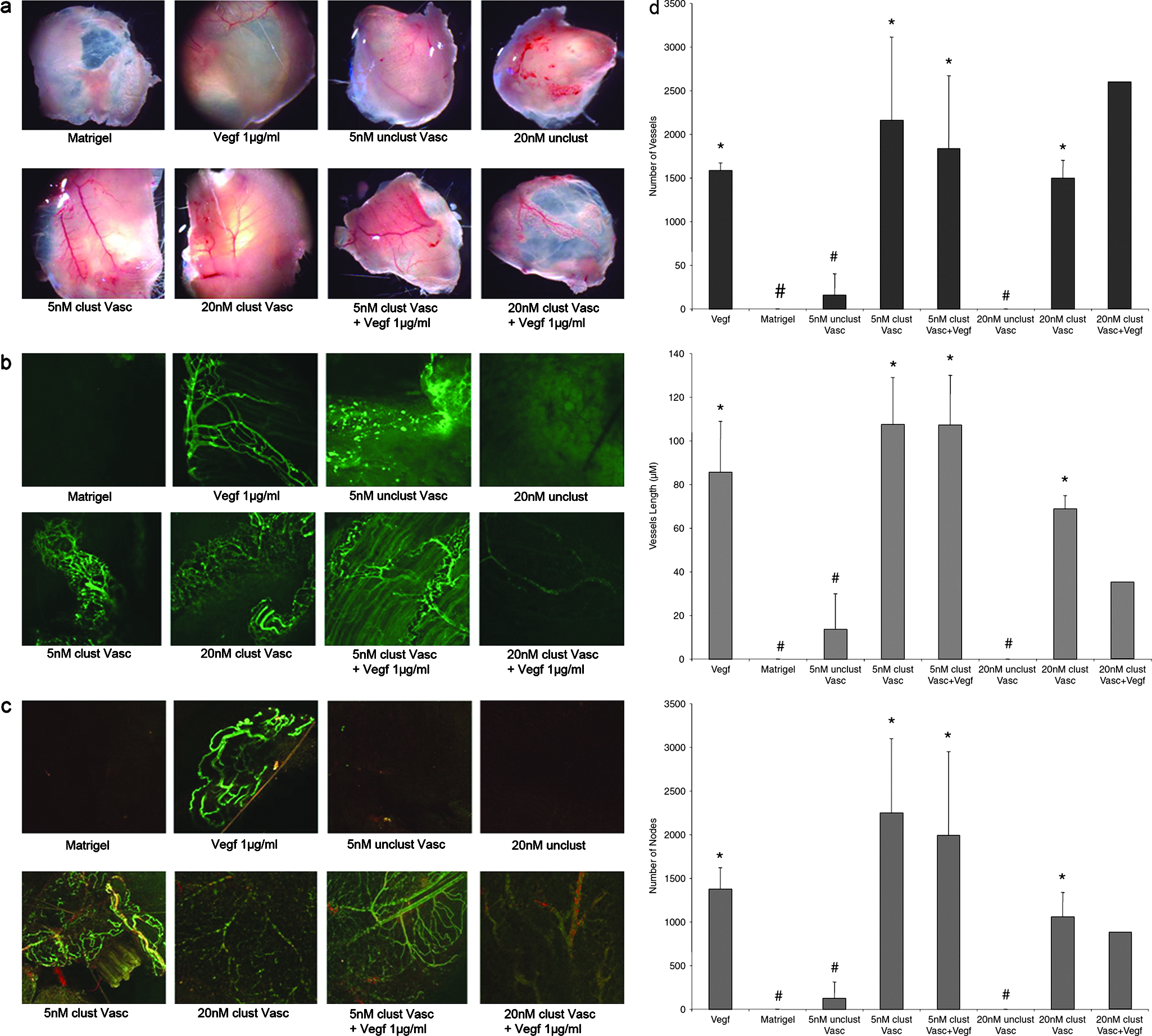
Clustered Vasculotide promotes a robust in vivo angiogenic response. (
Clustered Vasculotide promotes wound healing in genetically diabetic mice
Chronic wounds, such as neuropathic or neuroischemic foot ulcers, are prevalent in diabetic patients. These wounds may arise from the patient's inability to sense injury (neuropathic) or due to microvascular defects (neuroischemic). Animal models of type II diabetes exist and closely recapitulate wound healing defects seen in human diabetics. Here we used B6.Cg-m(+/+)Lepr(db)/J (db/db), a strain of diabetic mouse that presents with impaired wound healing, to test if clustered Vasculotide could improve wound closure times. Circular (6-mm diameter), full-excisional wounds were made equal distance apart on the dorsal side of the mice. Intrasite gel (Smith and Nephew), impregnated with PBS, bFGF, or clustered Vasculotide, were applied on the day of wounding (day 0), days 2, 4, and 6. Images of the wounds were taken on the same days and on the final day of the assay, day 7 (Fig. 4a). Significant decreases in wound margin in the clustered Vasculotide samples (20 nM, p = 0.05, n = 4) were noted as early as day 2 (Fig. 4a, b). Coincident with improved wound closure was a change in the appearance of the wounds treated with clustered Vasculotide. These wounds presented with a lighter, more mucoid–looking scab (Fig. 4c). Margin closure in the clustered Vasculotide–treated wounds (20 nM and 5 nM) continued at an accelerated pace for the duration of the assay. As was the case in vitro, clustered Vasculotide applied at lower concentration seemed to offer more benefit (see days 4 and 6; 20 nM vs. 5 nM; p = 0.05 compared to p = 0.01, respectively) (Fig. 4a, b). Although this may seem unusual, similar results were also noted with topical application of becaplermin (Regranex), a topical preparation of modified platelet derived growth factor (PDGF). 36 Although Vasculotide provided the fastest healing at early time points, by day 7 of treatment, wound closure promoted by bFGF at 10 μg/mL was statistically indistinguishable from clustered Vasculotide at 20 nM and 5 nM. Paraffin-embedded cross sections of all wounds stained with H&E and Masson's trichrome (Fig. 4d) were given to a pathologist for independent blind analysis. Scores ranging from 0 to 4 were assigned for each of the following: reepithelialization, granulation tissue and neovascularization, and inflammation (see Table 1 for detailed scoring criteria). Differences between vehicle and bFGF, although apparent in wound closure times, existed only at the level of inflammation (p = 0.0486) (Fig. 4e). Application of clustered Vasculotide resulted in delayed reepithelialization (20 nM, p = 0.0004 and 5 nM, p = 0.029) but promoted profound increases in regranulation and neovascularization (20 nM and 5 nM, p = 0.0006). Inflammation noted in the clustered Vasculotide samples was only slightly increased at 20 nM (p = 0.034) and was not significantly different from vehicle at the 5 nM dose (p = 0.0979). Overall collagen deposition, as assessed by Masson's trichrome staining, was dramatically increased at both of the clustered Vasculotide concentrations compared to the vehicle and bFGF samples. Immunohistochemical analysis of the wound beds with antibodies directed against PECAM1, ICAM 1, and Sma1 demonstrated clear differences between clustered Vasculotide–treated and untreated samples (Fig. 4f). Wounds treated with clustered Vasculotide displayed dramatic increases in the number of PECAM1-positive vessels within the granulation tissue. Importantly many of the new vessels in the granulation tissue were accompanied by Sma1-positive support cells. Further, these vessels were also positive for ICAM 1, a marker of activated ECs, 37 demonstrating an active wound healing process was occurring. The increase in vessel density was surprisingly not accompanied by an increase in lymphangiogenesis as detailed by similar numbers of podoplannin-positive vessels (data not shown). Based on pathological and immunohistochemical analysis we conclude that the improved wound closure times noted in the clustered Vasculotide samples are a likely consequence of strong contraction effects brought about by increases in collagen deposition and neovascularization.
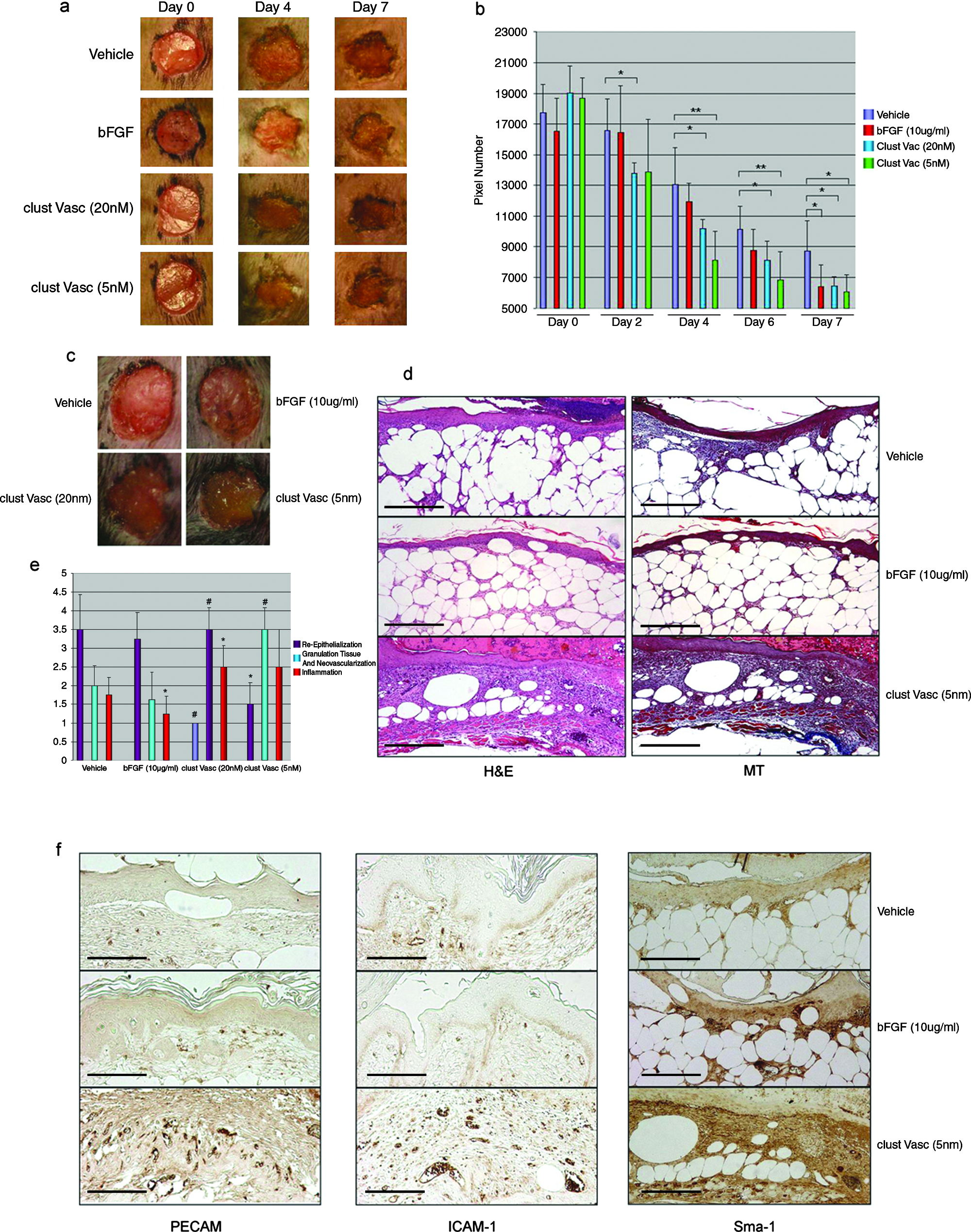
Clustered Vasculotide promotes accelerated wound closure in genetically diabetic mice. (
Discussion
Herein we describe the rationale design of a novel Ang mimetic. In vitro analysis of its properties profile effects that very closely resemble other native Angs. Specifically, we show activation of Tie 2 and signaling pathways that have been classically described downstream of Tie 2 including MAPK, AKT, and eNOS. However, unlike the effects of Ang1, we illustrate with the use of reconstitution experiments that Vasculotide uncouples Tie 2 signaling from integrin-dependent signaling. We report that treatment of primary ECs with clustered Vasculotide results in an induction of cell migration, MMP2 release, and protection from serum withdrawal–induced apoptosis. In vivo we show, when impregnated in Matrigel, clustered Vasculotide promotes a robust proangiogenic response that is marked by the production of well-arborized vessels that stain positively for the myogenic support cell marker, Sma1. Significantly, we show that clustered Vasculotide, when combined with VEGF, does not increase vessel number, vessel length, or number of branch points, but rather mitigates the tortuosity and improves pericyte recruitment to vessels above that which is seen with the addition of VEGF alone. We have examined the utility of clustered Vasculotide in the treatment of experimentally induced wounds using a widely accepted animal model of diabetes mellitus type II and find that clustered Vasculotide offers accelerated wound closure when compared to vehicle or bFGF controls. Using this model of impaired wound healing, we conclude that the improved wound closure times noted for clustered Vasculotide derive from increased granulation tissue and neovascularization of the wound.
Although surgical approaches to correct and restore collateral blood flow to ischemic tissues have steadily improved, many pathological states exist due to defects in small vessels. These vessels are not amenable to correction with surgical approaches, and even if they were, the underlying defects that lead to the dysfunctional vasculature are likely to remain. Establishment of a functional vascular plexus in the case of diabetic foot wounds is thought to be a rate limiting step necessary to promote efficient wound healing. Consistent with this notion, Galiano et al. recently presented promising work that demonstrates that therapeutic augmentation of vascular-specific growth factor, VEGF, can indeed provide wound healing improvements in a mouse model of diabetic wound healing.2,38 While the proangiogenic activities of VEGF have been profusely examined in the literature, thus far, limitations stemming from induction of vascular permeability and vessel tortuosity have prevented its clinical exploitation. Therapeutic modulation of the Tie 2 receptor represents an alternative novel vascular target that may confer benefits that exist beyond which most other single growth factors demonstrate due to its highly pleiotropic nature.39–41 Support for this concept stems from a report in which Kampfer et al. find that while Ang2 levels are constitutively elevated in the db/db diabetic mouse, there is further induction of Ang2 and VEGF expression upon wounding and in the early phases of wound healing. 1 They propose that Ang2 and VEGF collaborate to promote vessel destabilization and neovessel formation. However, they report that the increase in VEGF is transient, and in the absence of elevated VEGF, it is thought that persistently high levels of Ang2 contribute to further vessel destabilization and regression. 1 Given these findings, therapeutic augmentation of Ang1 or treatment methodologies that include application of an Ang1 mimetic in diabetic wound healing are predicted to be beneficial in reestablishing a balance in the Tie 2 signaling axis.
While we clearly show Vasculotide-dependent activation of Tie 2, it remains unknown whether the downstream effects perfectly recapitulate those of Ang1, Ang2, or some combination thereof. In fact, this question has proven difficult to address, as distinguishing features downstream of Ang1- or Ang2-dependent activation of the Tie 2 receptor are not well elucidated and appear highly context dependent. Recently, we and others have shown that Ang2, originally thought to be a Tie 2 antagonist, is more likely a partial agonist of the Tie 2 receptor, and that under conditions of endothelial stress, Ang2 possesses many of the same endogenous functions of Ang1, including resistance to vascular leak and apoptosis.10,42 Although the exact origin(s) of the differences and similarities that exist between the actions of Ang1 and 2 are poorly understood, unique structural characteristics differentiate these two ligands. Vasculotide was engineered to structurally mimic binding and clustering properties of Ang1. The strength, to which Vasculotide is able to activate the Tie 2 receptor, as tested in EC culture, suggests that it behaves as intended. However, in vivo, the activities of Vasculotide appear to resemble something of a hybrid between the known actions Ang1 and 2. For instance, use of Vasculotide in Matrigel plug assays demonstrates a potent induction of neovascularization, an activity that requires initial vessel destabilization and more closely resembles the reported actions of Ang2. While in the same assay we note that plugs containing Vasculotide are well supported by periendothelial cells, an event previously ascribed to Ang1.34,43 In this sense our results are consistent with those of Cho et al. 7 who report that use of their chimeric form of Ang1, COMP Ang1, both promotes neovascularization of the cornea and recruits perivascular support cells. One possible explanation for these results derives back to the unique structural characteristics of native Ang1 and 2. In the case of Cho et al., 7 synthesis and purification of COMP Ang1 gave rise to a heterogeneous mixture of multimers, with pentamers comprising the most abundant species. The synthesis of Vasculotide results in an obligate tetrameric structure. However, no formal studies have been conducted to assess the stoichiometric binding of Vasculotide to Tie 2. As such it remains unknown whether one molecule of Vasculotide is capable of binding four Tie 2 monomers. Factors that contribute to functional tetramerization of Tie 2 by Vasculotide are theorized to involve the angle of T7 peptide presentation as determined by avidin structure, the flexibility and length of the PEG molecular spacer, and the ratio of Vasculotide to free Tie 2 receptor available for binding. Given these considerations, it is possible that Vasculotide may be capable of engaging the Tie 2 receptor monomerically, dimerically, trimerically, or tetramerically. Data presented suggest that Vasculotide may mimic the effects of Ang1 and to a lesser degree Ang2 and, as such, may predominantly engage the Tie 2 receptor as a tetramer. Studies are currently underway to examine different clustering configurations aimed at manipulating biological outcomes mediated by Vasculotide.
Footnotes
Disclosure Statement
No competing financial interests exist.