
Abstract
The use of animal sera for the culture of therapeutically important cells impedes the clinical use of the cells. We sought to characterize the functional response of human mesenchymal stem cells (hMSCs) to specific proteins known to exist in bone tissue with a view to eliminating the requirement of animal sera. Insulin-like growth factor-I (IGF-I), via IGF binding protein-3 or -5 (IGFBP-3 or -5) and transforming growth factor-beta 1 (TGF-β1) are known to associate with the extracellular matrix (ECM) protein vitronectin (VN) and elicit functional responses in a range of cell types in vitro. We found that specific combinations of VN, IGFBP-3 or -5, and IGF-I or TGF-β1 could stimulate initial functional responses in hMSCs and that IGF-I or TGF-β1 induced hMSC aggregation, but VN concentration modulated this effect. We speculated that the aggregation effect may be due to endogenous protease activity, although we found that neither IGF-I nor TGF-β1 affected the functional expression of matrix metalloprotease-2 or -9, two common proteases expressed by hMSCs. In summary, combinations of the ECM and growth factors described herein may form the basis of defined cell culture media supplements, although the effect of endogenous protease expression on the function of such proteins requires investigation.
Introduction
A major impediment to the re-implantation of culture-expanded hMSCs for therapeutic use in vivo, however, is that current culture methodologies require the use of animal sera to provide appropriate nutrients, growth factor, and ECM components to support cell viability and growth. This poses a number of risks, including the xenobiotic transfer of disease, and in turn accounts for the increasing regulatory constraints on the use of cells that have been cultured in animal sera before therapeutic transplant. 13 A number of studies have applied various strategies to attempt to address this issue, including the use of patient-derived sera14,15 and platelet rich plasma (a rich source of growth factors).16–18 However, regulatory bodies find such undefined media components undesirable. Thus there is an imperative to develop effective culture media formulations suitable for ex vivo expansion of hMSCs without the use of animal products or non-defined components.
The major biological component of bone is type I collagen, but adult bone is also rich in other glycoproteins such as fibronectin and to a lesser degree vitronectin (VN).19,20 In addition, insulin-like growth factor (IGF)-I and -II, along with IGF binding protein-5 (IGFBP-5) and transforming growth factor beta 1 (TGF-β1), are among the most highly expressed growth factors in bone.21–25 Recently, the IGFs, together with the IGFBPs, have been shown to interact with each other and the matrix protein VN in vitro and to support functional responses in a number of epithelial cell types26–29 and osteoblasts. 30 TGF-β1 has also been shown to associate with VN and retain functional effects on cells. 31 Furthermore, interactions between the ECM and members of the IGF system and other growth factors, such as TGF-β1, are believed to have wide-ranging effects on hMSC function and bone tissue growth and turnover,32,33 raising interest in the potential use of these compounds for novel tissue-engineered therapeutics. Importantly, hMSCs are known to express a range of integrins, including αvβ3 integrins (the VN receptor), and growth factor receptors, including the IGF-IR and receptors for TGF-β1. 34 Some studies suggest that VN can induce osteogenic differentiation of hMSCs,35,36 whereas TGF-β1 has been shown to mediate proliferative responses in hMSCs through the induction of IGFBP-3 and IGF-I expression, which then act in an autocrine manner. 33 Thus the effects of TGF-β1 on hMSC and osteoblast function are complex and poorly understood.
Little information is available concerning the effects of IGFBP-5 on hMSCs despite findings that hMSCs express IGFBP-3 and -5 and IGF-I 33 and that the pro-differentiation steroid dexamethasone down regulates IGFBP-5 and IGF-I. 37 However, IGFBP-5 and its proteolytic fragments have been shown to confer IGF-independent effects on osteoblast cell function in mouse and human, including proliferation and phosphorylation of serine residues of a 420-kDa membrane-bound putative IGFBP-5 receptor.38–40 Therefore, this study was directed at investigating the functional effects of VN together with IGFBP-3 or -5 and IGF-I, or VN and TGF-β1, on hMSC functions such as metabolic activity, protein synthesis, and culture morphology.
Materials and Methods
Isolation and culture of hMSC from donor bone marrow samples
Bone marrow samples were collected, after informed consent and with hospital and university ethics committee approvals, from four elderly patients approximately 80 years of age (1 male and 3 female) presenting to The Prince Charles Hospital or The Holy Spirit Northside Hospitals in Brisbane for total knee or hip joint replacement surgical procedures as a treatment for osteoarthritis. Typically 2 to 5 mL of bone marrow was collected into tubes containing 5 mL of phosphate buffered saline supplemented with 200 U/mL of heparin and placed on ice for transport. Individual marrow samples were repeatedly passed through a 19 g needle to dislodge clumped cells, diluted to 30 mL with phosphate buffered saline, and passed through a 100-μm filter before overlaying onto 15 mL of Lymphoprep (Axis-Shield Poc As, Oslo, Norway). Samples were centrifuged at 400 g for 35 min at 20°C with zero brake, and the mononuclear cell fraction was transfered into fresh tubes containing 20 mL of Dulbecco's modified Eagle medium (DMEM; Invitrogen/GIBCO, Auckland, New Zealand) supplemented with 10% fetal calf serum (FCS; ThermoTrace, Noble Park, Vic, Australia), 50 U/mL penicillin/streptomycin (Invitrogen/GIBCO), and 5 μg/mL gentamycin (Invitrogen/GIBCO) (DMEM+). The cells were then seeded into tissue culture flasks at 1.6 × 105 cells/cm2 and incubated in a humidified 5% carbon dioxide (CO2)/95% air atmosphere at 37°C. The culture medium was changed after 5 days in the first instance and twice weekly thereafter. After four to six passages, the cells used in this study were characterzsed for their multilineage potential.
Preparation of tissue culture plates with VN and growth factors for metabolic activity and total cell protein assays
hMSC metabolic activity and total cell protein were assessed in the presence of solution-phase combinations of VN, IGFBP-3 or -5, and IGF-I or TGF-β1. Specifically, 52 μL of 1 μg/mL (52 ng/well) or 3.346 μg/mL (174 ng/well) VN and serum-free DMEM (sf-DMEM) solution or sf-DMEM alone was added to 96-well tissue culture plates and incubated for 2 h in a humidified 5% CO2/95% air atmosphere at 37°C. After incubation, 43.5 μL of 0.16 μg/mL (7 ng/well) TGF-β1 or 1.6 μg/mL (70 ng/well) IGFBP-3 or -5 in 0.5% bovine serum albumin and sf-DMEM, alone or in combination with 0.4 μg/mL (17.4 ng/well) IGF-I, were added to wells with or without VN and incubated at 4°C overnight. Plates were allowed to return to room temperature before seeding of cells. The final concentrations of the added proteins after cell seeding (final volume of 195.5 μL) were 3.55 nM or 11.9 nM for VN, 1.43 nM for TGF-β1, 11.9 nM for IGFBPs, and 11.9 nM for IGF-I.
Metabolic activity assay
Sub-confluent cultures of hMSCs were harvested using trypsin digestion, and 5000 cells were transfered into each well of a 96-well plate containing various combinations of VN, IGFBP-3 or 5, and IGF-I or TGF-β1 in a final volume of 195.5 μL. The cells were incubated under standard culture conditions for 24, 48, or 72 h before removal of conditioned medium and addition of 110 μL of 1:10 v/v WST-1 reagent (Roche Applied Sciences, Castle Hill, NSW, Australia) and sf-DMEM solution to each well. Absorbance readings at 450 to 650 nm were taken after 2 h of incubation.
Total protein assay and morphological analysis
Sub-confluent cultures of hMSCs were harvested, seeded into 96-well plates, and allowed to incubate as described above. The conditioned medium was recovered and the monolayers fixed using 4% w/v paraformaldehyde, incubated for 20 min, and subsequently stained with 0.1% w/v crystal violet. The monolayers were destained by immersion in a beaker of gently circulating tap water until stain visibly ceased to wash from the cell layer. The plates were then allowed to air dry overnight, and images of culture morphology were digitally captured at ×100 magnification using a Nikon Coolpix digital camera (Nikon, Lidcombe, NSW, Australia) mounted on a Nikon TE100 phase contrast microscope (Nikon). The crystal violet stain was then solubilized in 100 μL of 10% acetic acid and the absorbance determined at 595 nm.
The percentage of culture surface area covered was determined using Image J version 1.40d (National Institutes of Health, Bethesda, MD) 41 with McMaster Biophotonics Facility (MBF) plugins. 42 A 700- × 700-pixel (0.86 mm2) area of each image was analyzed using the following settings.
Apply band pass filter; to remove variation in background lighting.
Apply color balance set to 0.75; to convert background to white.
Convert image to 8-bit greyscale.
Manually apply threshold to maximally highlight area covered by cells.
Apply de-speckle function twice to remove non-specific noise from the background.
Analyze particles with particle size (mm2) set to 0.0001 to 1.0 for area fraction.
Images were saved as TIFF files and the area data exported to Excel for further analysis.
Gelatin zymography
Conditioned medium recovered from representative wells, as described above, was stored at −20°C until analyzed. Samples were analyzed for matrix metalloproteinase (MMP)-2 and MMP-9 gelatinolytic activity. Briefly, 4 μL of 5X loading buffer (0.05% bromophenol blue, 50% glycerol, 10% sodium dodecyl sulfate (SDS), 0.5 M Tris base, pH6.8) was added to 16 μL of conditioned medium samples, loaded onto freshly prepared 4% stacking/10% resolving polyacrylamide gels containing 1 mg/mL porcine gelatin (Sigma-Aldrich, St Louis, MO) and subjected to electrophoresis in Tris-glycine running buffer (25 mM Tris base, 246 mM glycine, 0.1% SDS, pH 6.8) under non-reducing conditions at 200 V and 4°C. The gels were then washed in 2.5% Triton-X100 for 30 min and then again for 60 min to remove SDS before incubation in buffer (50 mM Tris base, 10 mM calcium chloride, 50 mM sodium chloride, pH 7.6) for 48 h at 37°C. The gels were stained using 0.25% Coomasie Blue and destained in 10% acetic acid and 40% methanol (vol/vol) to reveal gelatinase activity, indicated as clear bands and discriminated according to molecular weight and comparison with MMP-2 and MMP-9 standards (Chemicon Pty/Ltd, Boronia, Vic, Australia).
Statistical analysis
Results are from four separate experiments (4 donors) performed in at least triplicate and are expressed as the corrected absorbance as a percentage of the 24-h –VN control (–VN) ± standard error of the mean. This repeated-measures design was analyzed using a linear mixed model in which donor was considered a random effect. If differences were detected between treatments using the global test, post hoc t-tests were conducted on the estimated marginal means to compare specific treatments from the 72-h data. Image analysis of morphological data was performed using Dunnett's test. Treatments were considered different when p < 0.05. It should be noted that the means and standard errors in Figures 1-3 do not represent those used in the linear mixed model, which accounts for the time course nature of the experiment and technical and biological sources of variation in the replicates. The symbols represent differences between treatments.

Human mesenchymal stem cell (hMSC) metabolic activity response to transforming growth factor beta 1 (TGF-β1) under serum-free conditions is dependent on vitronectin (VN) concentration. hMSCs were incubated for up to 72 h at 37°C with a combination of 3.55-nM or 11.9-nM VN and 1.43-nM TGF-β1. Cells seeded into wells without VN (–VN) or with standard growth medium (containing 10% fetal calf serum (FCS)) were employed as controls. Metabolic activity was measured using WST-1 reagent with absorbance read at 450 nm using 650 nm as reference. Results were corrected using data from sample blanks (serum-free medium and WST-1 alone) and are expressed as the mean corrected absorbance as a percentage of the 24-h –VN control (depicted by horizontal line) ± standard error of the mean. Significant difference between individual treatments and the 72-h –VN control or between individual treatments where connected by line was determined using a post hoc t-test and is indicated by α (p < 0.05) or β (p < 0.01). Significant differences between individual treatments and the 10% FCS control is indicated by * (p < 0.05) or # (p < 0.01).
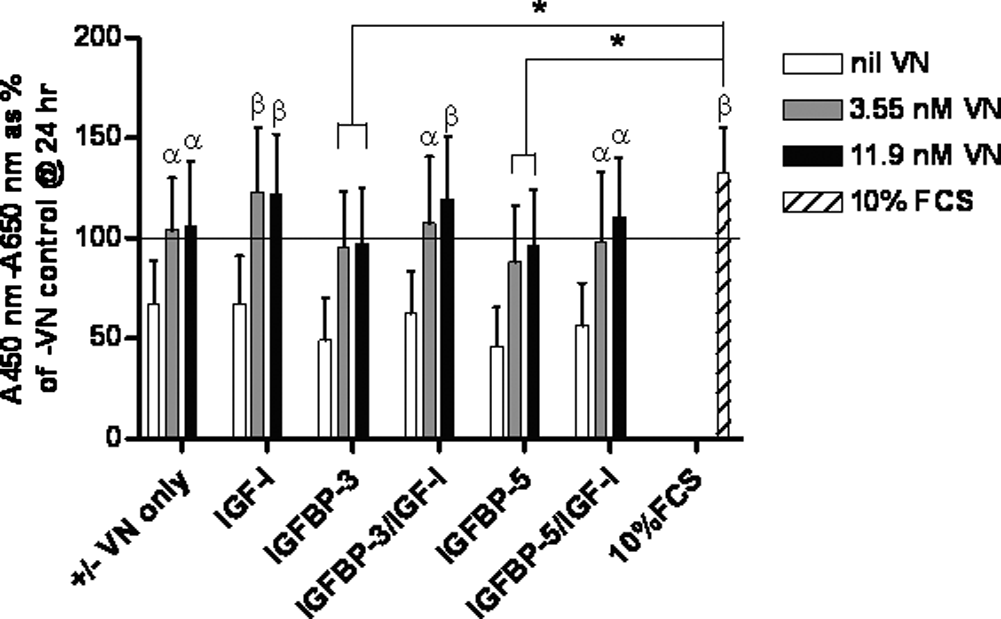
Vitronectin (VN) facilitates human mesenchymal stem cell (hMSC) metabolic activity in the presence of insulin-like growth factor binding protein (IGFBP)-3 or -5 and insulin-like growth factor (IGF)-I in serum-free conditions. hMSCs were incubated for up to 72 h at 37°C with various combinations of 3.55-nM or 11.9-nM VN, 11.9-nM IGFBP-3 or -5, and 11.9-nM IGF-I. Cells seeded into wells without VN or with standard growth medium (containing 10% fetal calf serum (FCS)) were employed as controls. Metabolic activity was measured using WST-1 reagent with absorbance read at 450 nm using 650 nm as reference. Results were corrected using data from sample blanks (serum-free medium and WST-1 alone) and are expressed as the mean corrected absorbance as a percentage of the 24-h control without VN (–VN) (depicted by horizontal line) ± standard error of the mean. Significant difference between individual treatments and the 72-h –VN control was determined using a post hoc t-test and is indicated by α (p < 0.05) or β (p < 0.01). Similarly significant differences between individual treatments and the 10% FCS control is indicated by * (p < 0.05) or # (p < 0.01).
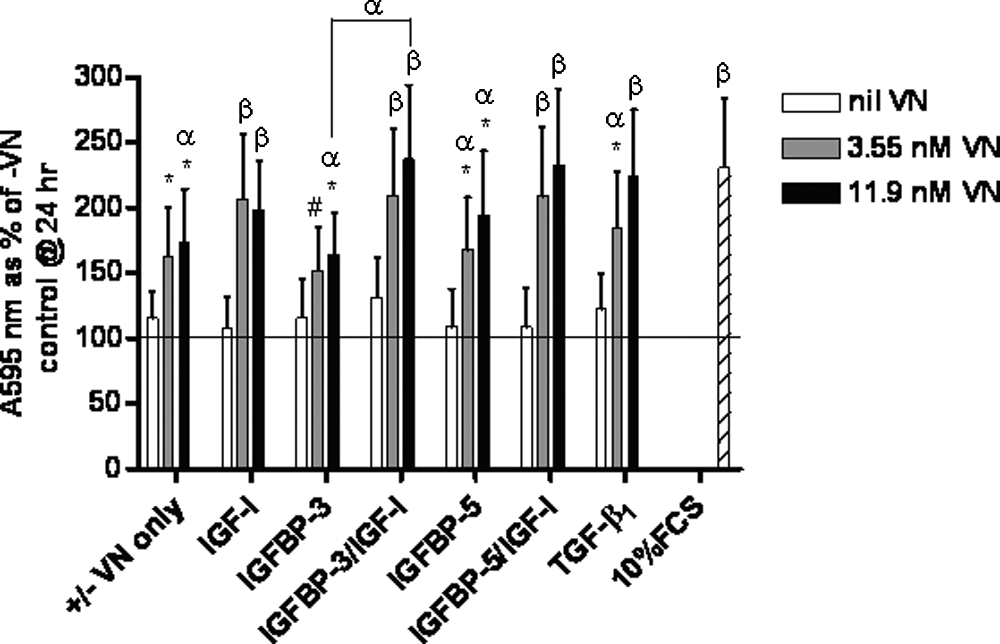
Vitronectin (VN) facilitates insulin-like growth factor binding protein (IGFBP) and insulin-like growth factor (IGF)-I or transforming growth factor beta 1 (TGF-β1)–mediated increase in total cell protein by hMSCs under serum-free conditions. Human mesenchymal stem cells (hMSCs) were incubated for up to 72 h at 37°C with various combinations of 3.55-nM or 11.9-nM VN, 11.9-nM IGFBP-3 or -5, and 11.9-nM IGF-I or 1.43-nM TGF-β1. Cells seeded into wells without VN or with standard growth medium (containing 10% FCS) were employed as controls. Cells were fixed, stained with crystal violet, and washed before the stain was liberated with 10% acetic acid and absorbance read at 595 nm. Results were corrected using data from sample blanks (10% acetic acid alone) and are expressed as the mean corrected absorbance as a percentage of the 24-h –VN control (depicted by horizontal line) ± standard error of the mean. Significant differences between individual treatments and the 72-h–VN control or between individual treatments where connected by line was determined using a post hoc t-test and is indicated by α (p < 0.05) or β (p < 0.01). Significant differences from the 10% FCS control are indicated by * (p < 0.05) or # (p < 0.01).
Results
hMSC metabolic activity response to TGF-β1 in serum-free conditions is dependent on VN concentration
TGF-β1 has been shown to associate with VN in vitro while retaining TGF-β1–mediated cell function and is also known to elicit a number of effects in hMSCs.31–33,43 Therefore, we wished to determine whether TGF-β1, together with VN, could influence hMSC culture metabolic activity. We measured the metabolic activity of hMSC cultures at 24, 48, and 72 h and found that, after 72 h, there was no difference in the response of cultures to 3.55-nM or 11.9-nM VN alone (Fig. 1). However, we observed that, after 72 h, the metabolic activity of hMSC cultures exposed to TGF-β1 plus 3.55-nM or 11.9-nM VN was significantly greater than hMSC cultures exposed to the 10% FCS control (p < 0.05 and p < 0.01, respectively) and to VN alone (p < 0.01) (Fig. 1). The metabolic activity of hMSC cultures exposed to TGF-β1 alone was equivalent to that assayed for the 72-h –VN control. These data suggest that TGF-β1 plus VN may act synergistically to stimulate hMSC culture metabolic activity over 72 h and that this response is dependent on VN concentration.
VN facillitates hMSC metabolic activity in response to IGFBP-3 or 5 and IGF-I in serum-free conditions
Given that hMSCs are known to express IGFBP-3 and -5 and IGF-I, 33 we wished to determine the effect of VN, IGFBP-3 or -5, and IGF-I on the metabolic activity of hMSC cultures in serum-free conditions. Similar to the response observed for TGF-β1, IGF-I plus 3.55-nM or 11.9-nM VN resulted in significantly higher hMSC culture metabolic activity than the 72-h –VN control (p < 0.01). Similarly, IGFBP-3/IGF-I plus 3.55-nM or 11.9-nM VN resulted in significantly greater metabolic activity than the 72-h –VN control (p < 0.05 and p < 0.01, respectively) as did IGFBP-5/IGF-I (p < 0.01) (Fig. 2). Although cultures exposed to IGFBP-3 or −5 plus 3.55-nM or 11.9-nM VN expressed greater metabolic activity after 72 h than the 72-h –VN control, results for these treatments failed to reach statistical significance. Although these treatments resulted in statistically lower responses than hMSC cultures exposed to 10% FCS (p < 0.05), all other treatments containing VN were found to induce statistically equivalent responses to cells cultured in 10% FCS (Fig. 2). Taken together, these data indicate that VN was the major factor contributing to hMSC culture metabolic activity and that the addition of IGFBP-3 or IGFBP-5 plus IGF-I could not significantly enhance the response over that obtained by VN alone or VN plus IGF-I (Fig. 2).
VN facilitates IGFBP-3 or -5/IGF-I– or TGF-β1–mediated increase in total cell protein by hMSCs in serum-free conditions
To determine whether hMSC cultures exposed to the above-mentioned proteins exhibited cumulative effects, we measured the relative total cellular protein. After 72 h, the difference in VN concentration did not affect the relative total protein accumulation within the monolayer culture of hMSCs unless VN was absent (Fig. 3). However, cultures exposed to the 11.9-nM VN-alone treatment were found to have significantly higher total protein than the –VN control (p < 0.01), whereas cultures exposed to IGF-I plus VN at either concentration resulted in significantly higher total protein accumulation than in the –VN control (p < 0.01). We also found that, of the IGFBPs tested, only IGFBP-5 plus 3.55-nM VN enhanced total protein accumulation (p < 0.05). hMSCs exposed to IGFBP-3 or -5 plus 11.9-nM VN had significantly higher levels of total protein than the –VN control (p < 0.05), as did cultures exposed to IGFBP-3 or -5/IGF-I with either concentration of VN (p < 0.01). hMSCs exposed to IGFBP-3/IGF-I plus 11.9-nM VN contained significantly more total protein than cells cultured in IGFBP-3 plus 11.9-nM VN alone (p < 0.05) (Fig. 3).
We found that TGF-β1 with 3.55-nM and 11.9-nM VN enhanced hMSC total protein accumulation significantly more than the –VN control (p < 0.05 and p < 0.01, respectively) or the TGF-β1–only treatment (p < 0.01). No significant difference was observed in total cellular protein accumulation between the 10% FCS control and treatments containing VN except 3.55-nM and 11.9-nM VN only (p < 0.05), IGFBP-3 with 3.55-nM VN or 11.9-nM VN (p < 0.05 and p < 0.01, respectively), IGFBP-5 with 3.55-nM VN or 11.9-nM VN (p < 0.05), and TGF-β1 with 3.55-nM VN (p > 0.05) (Fig. 3).
Taken together, these data indicate that addition of IGFBP-3 or -5 together with IGF-I plus VN or, similarly, TGF-β1 plus 11.9-nM VN can stimulate an increase in total protein over 72 h equivalent to 10% FCS and that this response is largely dependent on the presence of VN.
IGF-I and TGF-β1 induce aggregation of hMSC culture morphology under serum-free conditions
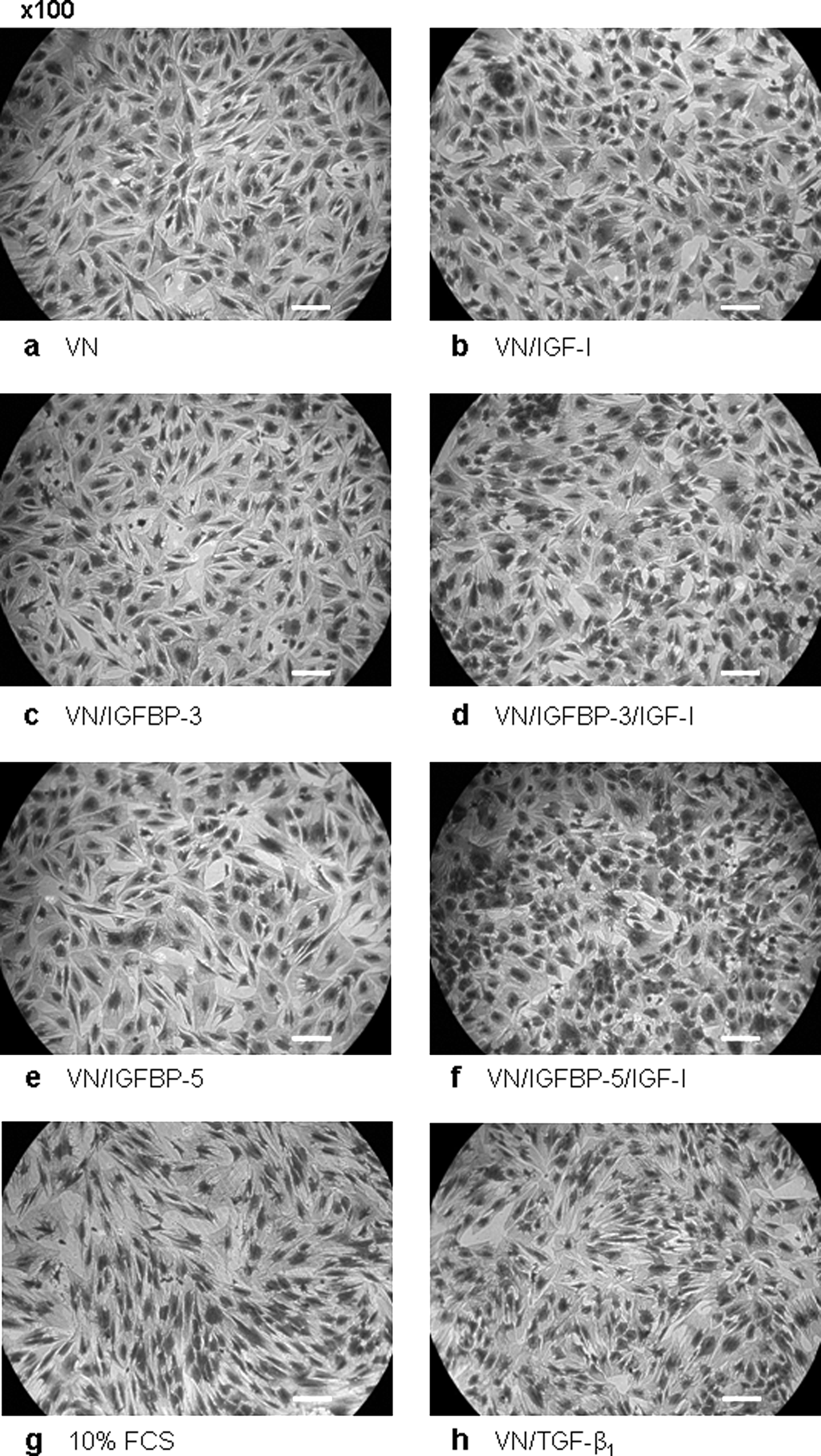
Examination of culture morphology revealed that, after 24 h, few viable cells remained in cultures without VN (not shown). In contrast, cultures exposed to treatments containing 3.55-nM VN (Fig. 4a) or 11.9-nM VN (not shown) were viable and relatively evenly distributed across the culture vessel surface, with most cells appearing spread and well attached (Fig. 4a.1–6,8). However, by 48 h, cultures exposed to 3.55-nM VN plus IGF-I or TGF-β1 (±either of the IGFBPs) began to retract or aggregate, leaving large gaps in the monolayer (Fig. 4b.2,4,6,8). Cultures that were exposed to 3.55-nM VN plus IGFBPs retracted only slightly (Fig. 4b.3,7,5). After 72 h, hMSCs exposed to 3.55-nM VN plus IGF-I or TGF-β1 had further contracted, leaving large areas of culture surface devoid of cells (Fig. 4c.2,4,6,8), whereas cultures exposed to 3.55-nM VN plus IGFBP-3 or -5 had developed similar but much smaller gaps in their respective monolayers (Fig. 4c.3,5). The aggregation effect was not observed in cultures exposed to 3.55-nM VN alone (Fig. 4a.1,b.1,c.1). Similarly, hMSCs exposed to 10% FCS remained relatively confluent throughout the culture period (Fig. 4a.7,b.7,c.7). These data indicate that, in the presence of 3.55-nM VN in a serum-free environment, IGF-I and TGF-β1 can induce time-dependent aggregation and contraction of hMSC cultures.

Insulin-like growth factor (IGF)-I and transforming growth factor beta 1 (TGF-β1) induce aggregation of human mesenchymal stem cell (hMSC) culture morphology under serum-free conditions. Sub-confluent cultures of hMSCs were harvested using trypsin digestion before seeding into wells of 96-well plates containing various combinations of 3.55-nM vitronectin (VN) and 11.9-nM IGF binding protein (IGFBP)-3 or -5 and 11.9-nM IGF-I or 1.43-nM TGF-β1 at a density of 5000 cells/well. Cells seeded into wells without VN or with standard growth media (containing 10% fetal calf serum) were employed as controls. Plates were incubated for 24 h (
IGF-I– and TGF-β1–mediated hMSC culture aggregation is dependent on VN concentration
In contrast to the morphological changes observed in hMSC cultures exposed to 3.55-nM VN in the presence of IGF-I or TGF-β1, hMSC cultures exposed to 11.9-nM VN did not exhibit any evidence of aggregation and disruption of the monolayer or gap formation, even after 72 h (Fig. 5b,d,f,h). Similarly, hMSC cultures exposed to IGFBP-3 or -5 in the presence of 11.9-nM VN or cultures exposed to 11.9-nM VN alone had not undergone aggregation (Fig. 5a,c,e). Thus, at higher concentrations, VN appears to have an antagonistic effect on the time-dependent aggregation of hMSC cultures exposed to IGF-I and TGF-β1.
Insulin-like growth factor (IGF)-I– and transforming growth factor beta 1 (TGF-β1)–mediated human mesenchymal stem cell (hMSC) culture aggregation is dependent on vitronectin (VN) concentration. Sub-confluent cultures of hMSCs were harvested using trypsin digestion before seeding into wells of 96-well plates containing various combinations of 11.9-nM VN (
To further characterize the observed aggregation phenomena, we performed image analysis of the digital photomicrographs taken after 72 h. ImageJ was used to convert a select area (0.86 mm2) of each image to binary code (black and white), and the percentage of surface area covered by the hMSC cultures in response to each treatment was determined. Representative images illustrating this process are included in Figure 6a. hMSCs exposed to IGF-I or TGF-β1 occupied significantly less area than the cultures exposed to –VN control, in which cells were cultured without VN (p < 0.01) (Fig. 6b). Similarly, cultures treated with IGF-I, IGFBP-5 and IGF-I (p < 0.05), or IGFBP-3 and IGF-I (p < 0.01) covered significantly less surface area than cultures treated with 3.55-nM VN alone. Although TGF-β1 also caused cells cultured in the presence of 3.55-nM VN to cover less surface area, this was not statistically significant. In contrast, the surface area that hMSCs occupied was greater than 70% when cultured with 11.9-nM VN. Furthermore, all treatments at this VN concentration yielded statistically equivalent cell coverage to VN alone (Dunnett's test) (Fig. 6b). Taken together, these data further support our interpretation that the higher VN concentration abrogated the reproducible culture aggregation phenomenon that we observed at the lower VN concentration.

Insulin-like growth factor (IGF)-I and transforming growth factor beta 1 (TGF-β1)–mediated decrease in human mesenchymal stem cell (hMSC) surface area coverage is dependent on vitronectin (VN) concentration. Digital images of crystal violet stained cultures were analyzed using ImageJ (National Institutes of Health, Bethesda, MD)
41
and McMaster Biophotonics Facility plugins
42
to determine the percentage of area covered by hMSCs in response to various combinations of 3.55-nM or 11.9-nM VN and 11.9-nM IGFBP-3 or -5 and 11.9 nM-IGF-I or 1.43-nM TGF-β1 after 72 h. A representative image is shown to illustrate the conversion of a defined area of the original image to binary in preparation for area analysis (
VN, IGFBP-3 or -5 and IGF-I or TGF-β1 do not effect the accumulation of MMP-2 and MMP-9 in hMSC-conditioned medium
Mesenchymal cells are known to express a variety of proteases, including members of the MMP family of proteins. 44 IGF-I and TGF-β1 are known to regulate expression of proteases in cells of mesenchymal origin, 45 prompting us to hypothesize that the aggregation and contraction phenomenon that we observed in the presence of IGF-I or TGF-β1 was due to the action of secreted proteases. We investigated whether the presence of IGF-I or TGF-β1 influenced the secretion of MMP-2 or -9 by hMSCs using gelatin zymography. To our surprise, we observed no difference in MMP-2 or -9 activity between any of the treatments (Fig. 7a–d). Indeed, our data from hMSCs cultured in serum-free media suggests that hMSCs constitutively secrete MMP-2 and -9 (Fig. 7a). These data indicated that IGF-I and TGF-β1 did not influence the secretion of MMP-2 and -9 into the conditioned medium and therefore that it is unlikely that either of these proteases was directly responsible for the aggregation of hMSC cultures in response to IGF-I or TGF-β1.
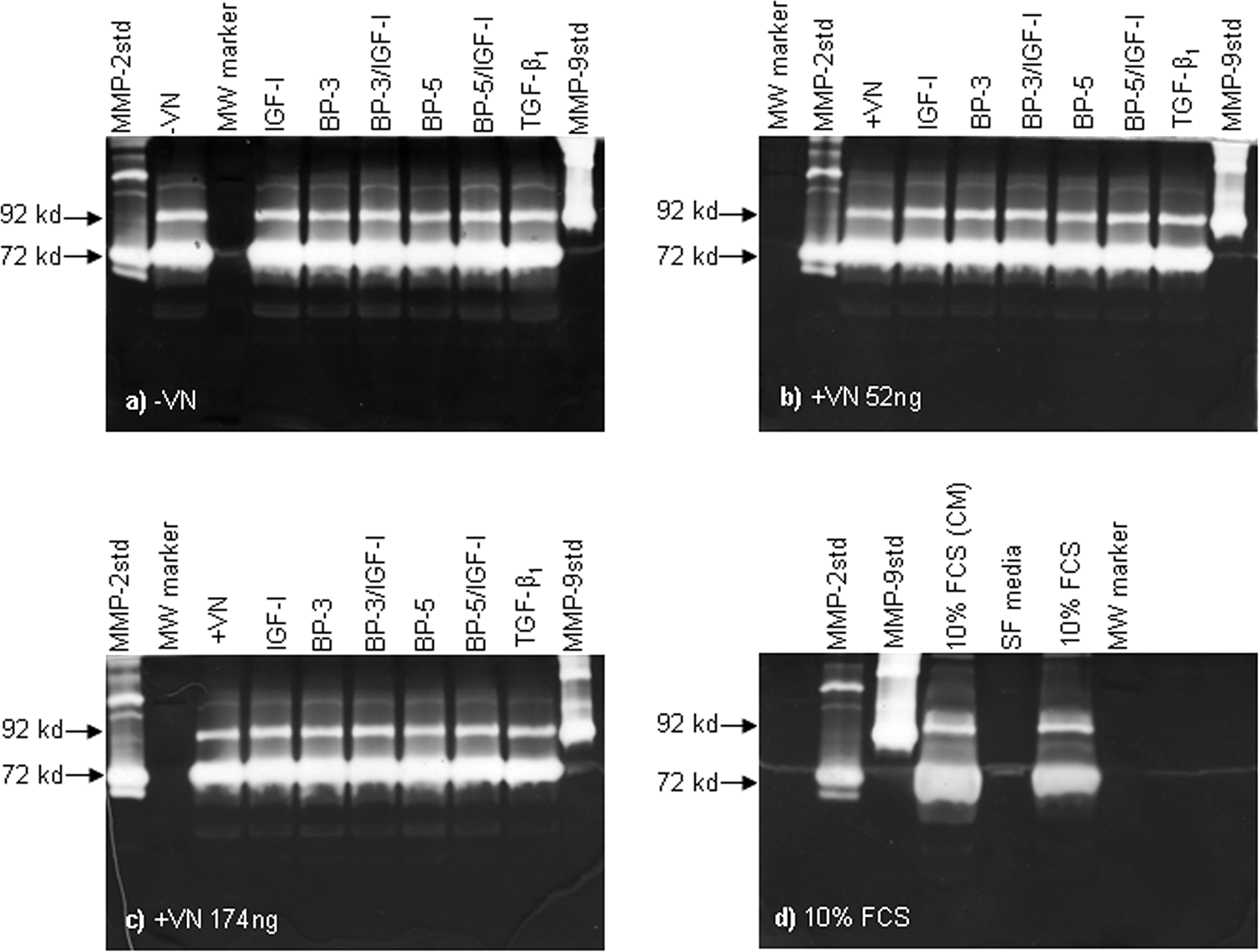
Vitronectin (VN), insulin-like growth factor binding protein (IGFBP)-3 or -5 and insulin-like growth factor (IGF)-I or transforming growth factor beta 1 (TGF)-β1 does not effect the accumulation of matrix metalloprotease (MMP)-2 and MMP-9 in human mesenchymal stem cell (hMSC)-conditioned medium. Conditioned medium was kept from representative wells of 72-h cultures from separate donors before fixing of cells for crystal violet staining during the total protein assays as detailed above. Collected samples (20 μL) were run on 10% gelatin zymograms before washing of gels in 2.5% Triton-×100 and then incubated for 48 h at 37°C in activation buffer. The zymograms were then stained for 2 h with Coomasie blue R250 stain followed by de-staining. Gelatinase activity was then visualized as clear bands and discriminated by comparison with purified MMP-2 and MMP-9 standards.
Discussion
The successful in vitro culture of hMSCs is still largely dependant upon the use of relatively undefined animal-derived products, particularly FCS. Thus, as regulatory authorities require increasingly stringent standards for human therapeutic agents, 13 the development of culture methodologies for hMSCs that include highly defined medium components, of non-animal origin, has become a vital area of of tissue engineering.14,46–49 We report here studies into the functional effect of the ECM glycoprotein VN in association with various combinations of IGFBP-3 or -5, IGF-I, or TGF-β1 on hMSC behavior in a serum-free environment.
First, we measured the metabolic activity of hMSCs exposed to the various combinations of VN and growth factors and found that VN was the major influence on hMSC metabolic activity. The metabolic activity of cultures without VN was substantially less than that assayed in the 24-h –VN control. We interpret this to indicate that cell adhesion was vital in this system in order for hMSCs to respond to exogenously added growth factors. Our observation that there was no difference in the response of hMSCs exposed to IGF-I plus either of the IGFBPs plus VN or to IGF-I plus VN alone, corroborates the findings of a previous study with hMSCs which found no difference in hMSC number when treated with either IGF-I, IGFBP-3 or IGF-I + IGFBP-3. 33 Although the responses reported by Kvieborg et al. 33 were significantly greater than the control treatment alone, in contrast to our observations, the authors initially cultured the cells in the presence of 10% FCS and replaced this with medium supplemented with 0.5% FCS and IGF-I, IGFBP-3, or both after 48 h. Thus, the elevated response (FCS in medium only) may have been due in part to (additional) unknown factors present in the 0.5% FCS that were absent in our study. 33
We found that only TGF-β1 plus 3.55-nM or 11.9-nM VN stimulated a significant increase in metabolic activity, greater than the activity supported by 10% FCS (Fig. 1). This indicates that TGF-β1, via downstream signalling effectors, has a direct stimulatory effect on hMSC metabolic activity. Alternatively, TGF-β1 may indirectly stimulate metabolic activity by up-regulating the expression of IGFBP-3 and IGF-I, as previously demonstrated. 33 The latter possibility is intriguing because the responses of hMSC cultures exposed to exogenously added IGFBP-3 plus IGF-I were equivalent to 10% FCS, whereas the responses to TGF-β1 plus either concentration of VN were significantly greater than the response to 10% FCS (Fig. 1, 2). Nevertheless, the IGFBP-3 used in this study was produced in Escherichia coli and was non-glycosylated (∼35 kDa), whereas hMSCs exposed to TGF-β1 produce 38-kDa and 42-kDa isoforms of IGFBP-3, representing partially and fully glycosylated isoforms.33,50 Thus the basis for the difference in response remains unclear. The, differences in glycosylation status of the IGFBP-3 and the level of expression of IGFBP-3 or IGF-I may play an important role in the cellular response. In addition, human lung fibroblasts have been shown to increase bromodeoxyuridine incorporation in response to VN plus TGF-β1 than TGF-β1 alone. This response was shown to be due to greater αvβ3 integrin expression and induced complex formation between TGFβIIRs and αvβ3 integrins, causing upregulation of cyclin D1 and synergistically enhanced proliferation. 51 Taken together, these data support our finding that hMSC metabolic activity is synergistically enhanced in response to TGF-β1 plus 3.55-nM VN and is further enhanced by higher concentrations of VN (11.9 nM). It is difficult to dismiss an increase in integrin binding sites; (e.g., cell surface αvβ3 integrins), and perhaps an autocrine–paracrine response to endogenously expressed IGFBP-3 and IGF-I is responsible. However, further investigation into the specific mechanisms underpinning this response may enable specific conclusions about the role of TGF-β1, VN, IGFBP-3, or IGF-I and their respective receptors to be drawn.
Although, 11.9-nM VN plus IGFBPs plus IGF-I resulted in significantly greater hMSC total protein accumulation than in the controls, Jia and Heersche reported that IGF-I stimulated osteoprogenitor cell proliferation, 52 whereas other authors have reported that IGF-I has no effect on hMSC proliferation or differentiation. 53 Andress and Birnbaum have shown that an osteoblast-derived isoform of IGFBP-5 enhances IGF-I–stimulated osteoblast mitogenesis. On the other hand, purified IGFBP-3 has been reported to (dose-dependently) inhibit IGF-I–stimulated osteoblast mitogenesis in a dose-dependant manner. The authors postulated that this was due to sequestration of IGF-I by IGFBP-3, inhibiting IGF-I association with the IGFIR. 25 IGF-I together with the bone matrix protein osteopontin, an αvβ3 integrin ligand, has been shown to stimulate porcine smooth muscle cell (pSMC) growth, a response that was further enhanced to levels equivalent to 10% FCS with the addition of IGFBP-5. 54 Furthermore, interaction between IGFBP-5 plus VN has been shown to be critical for the enhancing effect of IGFBP-5 on IGF-I–mediated pSMC DNA synthesis. 54 Indeed, previous research within our laboratory has confirmed and expanded the concept of enhanced cellular response to complexes composed of IGFBP-5 plus IGF-I bound to VN.26–28
Interestingly, the time-dependent manner in which hMSC cultures exposed to 3.55-nM VN and IGF-I– or TGF-β1–expressed culture contraction or aggregation phenomena (Fig. 4a–c) appeared initially to be the result of cell adhesion or cell death. However, closer analysis of the total protein data revealed that there was little or no difference in total protein content between these treatments and that of hMSCs exposed to 3.55-nM VN alone (Fig. 3). We interpret this to suggest that there is no significant change in cell number during this time. Importantly, cultures that were exposed to 11.9-nM VN appeared to be protected from this effect (Fig. 5). Image analysis (Fig. 6a), which showed that, after 72 h, there was generally less surface area coverage by cells exposed to IGF-I or TGF-β1 and 3.55-nM VN than to VN alone and that 11.9-nM VN abrogated this effect further supported these observations (Fig. 6b). The aggregation of mouse osteoblastic cells exposed to TGF-β1 has been described previously. 55 Similarly, MC3T3-E1 cells cultured in serum-free medium and exposed to TGF-β1 developed culture morphologies identical to those of our hMSCs. However, when the MC3T3-E1 cells were cultured on a type I collagen substratum, the effect was greatly diminished. 56 Karsdal et al. have also reported that activation of extracellular signal–regulated kinase 1/2 (ERK1/2) and p38 mitogen-activated kinase (MAPK) is greater in response to TGF-β1. Activation of ERK1/2 and MAPK may partly explain the increase in hMSC metabolic activity observed in this study. Importantly, specific inhibitors revealed that the p38 MAP kinase pathway, but not the ERK1/2 or Janus kinase (JNK) pathways were involved in the MC3T3-E1 morphological change observed in response to TGF-β1. 56
The fact that TGF-β1 is known to induce plasminogen activator inhibitor (PAI-1) expression may also explain this phenomenon in part.57,58 PAI-I has recently been shown to mediate cell detachment from VN substrata by initiating endocytosis of urokinase–urokinase receptor alpha v integrin complexes. 59 This conincides with our observation that the aggregation response was attenuated in the presence of higher VN concentration (i.e. availability of more integrin binding sites). This is further supported by the finding that by over expressing the number of αvβ3 integrins on the surface of chinese hamster ovary (CHO) cells, PAI-I mediated cell detachment is drastically reduced. 59 To the best of our knowledge, the study reported herein is the first to describe this phenomenon in hMSCs and demonstrate that, by manipulating the concentration of VN, the observed response can be attenuated. Although a similar response has been reported for osteoblasts exposed to parathyroid hormone, which adopted a stellate morphology,56,60 we are unaware of any studies other than the present study that report a similar response to IGF-I. The specific molecular mechanisms underlying the observed responses remains to be elucidated.
We initially speculated that the observed growth factor–mediated aggregation may be due to secretion of proteases that affected hMSC attachment to the substratum. However, we found that there was no correlation between the functional expression of MMP-2 or -9, as determined using gelatin zymography, and the growth factor–induced hMSC aggregation. Similar results were observed in MC3T3-E1 cells exposed to TGF-β1; TGF-β1 was found not to affect the expression of MMP-2 but induced MMP-13 expression into the conditioned medium. 56 Given that hMSCs exposed to no treatment whatsoever (the -VN control) expressed MMP-2 and -9, we concluded that hMSCs constitutively expressed both proteases (Fig. 7a, Lane 2). This appears to be relatively common for MMP-2 across a number of different cell types. 44 Interestingly VN has been shown to induce MMP-2 expression in melanoma cells, 61 and MMP-2 and -9 are known to interact or cooperate with αvβ3 integrins to regulate cell migration and invasion.62,63 Others have reported that over-expression of tissue inhibitor of metalloproteinases (TIMP)-2, an MMP-2 inhibitor, results in reduced MMP-2 activity (as expected) and restores melanoma cell attachment and spreading on a VN substratum, also showing that activated MMP-2 did not degrade the VN substrate. 64 Indeed, solution-phase VN has also been shown to be insensitive to MMP-2–mediated proteolysis. 61 We did not investigate the expression of TIMPs by hMSCs in this study; however, investigation of the fate of the proteins used in this study are required to more fully understand the dynamics within such serum-free culture environments.
In conclusion, the data presented in this study indicate that VN is a valuable matrix component that provides significant initial support for the serum-free culture of hMSCs. In addition, the concentration of VN is an important factor in the maintenance of the culture integrity, and further investigation into the effect of endogenously expressed proteases on the functionality of added matrix components, growth factors, or other constituents is required. Furthermore, in the presence of other factors, such as TGF-β1, additional stimuli can be achieved. Lastly, specific investigation of the mechanisms of TGF-β1 action on hMSC function in the serum-free environment is required.
Footnotes
Acknowledgments
The authors would like to thank Professor Ross Crawford and the medical staff at the Holy Spirit Northside and Prince Charles Hospitals for the collection of bone marrow samples used in this study. Thanks also to Dr. Cameron Hurst of the IHBI, QUT, for his useful advice and discussions regarding the statistical analysis, and Dr. Zareen Yameen, for much of the hMSC characterization work. The authors are also grateful for research funding from the QUT Strategic Collaborative grant scheme and Tissue Therapies P/L.