
Abstract
Collagen-fibronectin gels containing Bcl-2–transduced human umbilical vein endothelial cells (Bcl-2-HUVEC) implanted in the abdominal walls of immunodeficient mice form mature microvessels invested by host-derived smooth muscle cells (SMC) by 8 weeks. We tested the hypothesis that coengraftment of human aortic SMC (HASMC) could accelerate vessel maturation. To prevent SMC-mediated gel contraction, we polymerized the gel within a nonwoven poly(glycolic acid) (PGA) scaffold. Implanted grafts were evaluated at 15, 30, and 60 days. Acellular PGA-supported protein gels elicited a macrophage-rich foreign body reaction and transient host angiogenic response. When transplanted alone, HASMC tightly associated with the fibers of the scaffold and incorporated into the walls of angiogenic mouse microvessels, preventing their regression. When transplanted alone in PGA-supported gels, Bcl-2-HUVEC retained the ability to form microvessels invested by mouse SMC. Interestingly, grafts containing both Bcl-2-HUVEC and HASMC displayed greater numbers of smooth muscle α-actin–expressing cells associated with human EC-lined arteriole-like microvessels at all times examined and showed a significant increase in the number of larger caliber microvessels at 60 days. We conclude that SMC coengraftment can accelerate vessel development by EC and promote arteriolization. This strategy of EC-SMC coengraftment in PGA-supported protein gels may have broader application for perfusing bioengineered tissues.
Introduction
Cotransplantation of primitive mesenchymal cells (e.g., 3T3 cells) has been reported to stabilize vessels formed from human EC. 8 We wondered if differentiated SMC would perform this same function, and possibly with greater efficiency. Coengraftment of fully differentiated SMC with EC in 3D collagen gel–based systems for the development of tissue-engineered microvessels has been limited by SMC-mediated gel contraction. A potential strategy to overcome the biomechanical limitations of protein gel systems is the incorporation of a biodegradable scaffold material at the time of cell suspension and gel polymerization. Perhaps the most widely studied biomaterial suitable for such applications is poly(glycolic acid) (PGA). PGA is a commercially available product that is currently FDA-approved for a number of applications and has been used extensively in both microvascular and large vessel tissue engineering.10,15–18 The degradation and inflammatory characteristics have been well described, and the byproducts of PGA degradation are known to have little effect on the viability of vascular SMC or EC.15,19,20
Here, we describe a PGA-stabilized system for coengraftment of Bcl-2-HUVEC and enhanced green fluorescent protein (EGFP)–transduced human aortic SMC (HASMC) in protein gels. We find that the PGA scaffold can effectively prevent protein gel contraction by HASMC. Unlike unsupported protein gels, subcutaneous implantation of PGA scaffold–supported protein gels induces a host foreign body response and a transient angiogenic response, even in the absence of engrafted cells. In PGA-supported gels containing only HASMC, some transplanted HASMC interact with the scaffold material, tightly associating with PGA polymer fibers, while other HASMC associate with host-derived angiogenic vessels and act to prevent their regression. PGA does not interfere with vessel formation by implanted Bcl-2-HUVEC. In implants containing both HASMC and Bcl-2-HUVEC, many HASMC are recruited to human EC-lined vessels, promoting early formation of arteriole-like vessels at 15 days and enhancing the extent of microvessel remodeling into larger caliber vascular conduits by 60 days.
Materials and Methods
Cell procurement and graft implantation
All human cells were obtained using protocols approved by the Yale Human Investigation Committee. All animal experiments were performed according to Yale Institutional Animal Care and Use Committee protocol number 2006-07863.
Cells
HUVEC were isolated, cultured, and transduced to express the caspase-resistant D34A mutant form of Bcl-2, as previously described. 6 The efficiency of transduction, which routinely reached greater than 90% without selection, was confirmed by flow cytometry of permeabilized cells with an anti-Bcl-2 antibody (1:10; BD Biosciences, San Diego, CA). HASMC were isolated by explant outgrowth from minced human thoracic aorta, as described previously. 21 For transduction of SMC, EGFP DNA was inserted in the LZRSpBMN vector (provided by Dr. G.P. Nolan, Stanford University, Palo Alto, CA). The retroviral vector DNA containing EGFP was directly transfected into the Phoenix packaging cell line by Lipofectamine 2000 (Invitrogen, Carlsbad, CA) and selected using puromycin (10 μg/mL; Calbiochem, San Diego, CA). Supernatants were collected in M199 media (Gibco, Grand Island, NY) containing 10% fetal bovine serum (FBS) with 2 mM L-glutamine, and 100 U/mL penicillin. Supernatants were then filtered using a 0.45 μm syringe filter (BD Biosciences) and supplemented with polybrene (8 μg/mL; Sigma-Aldrich, St. Louis, MO). The efficiency of transduction, which routinely reached 55–65%, was assessed by flow cytometry.
Preparation of protein gels
Bcl-2-HUVEC (3.0 × 106 cells/mL final concentration) and HASMC (1.5 × 106 cells/mL final concentration) were suspended in rat tail type I collagen (final concentration 1.5 mg/mL; BD Pharmingen, Bedford, MA) with 1 × M199 (diluted from 10 × ), 75 μg/mL fibronectin, prepared from human plasma, 2.5% 1M HEPES, and 2.2 mg/mL
Graft analysis
Approximately, one half of each graft was fixed in formalin and paraffin embedded for staining with hematoxylin and eosin (H&E) or analyzed by immunohistochemistry. Six μm formalin-fixed, paraffin-embedded tissue sections were used for immunohistochemical analysis of harvested tissue. Primary antibodies reactive with Bcl-2, GFP, or smooth muscle cell–specific alpha-actin (α-SMA) were used for enumeration and characterization of vascular profiles (Bcl-2 1:100 [BD Pharmingen], α-SMA 1:100 [Novocastra, Newcastle upon Tyne, UK], GFP 1:500 [AbCam, Cambridge, MA], and Mac-3, an antibody specific for mouse macrophages, 1:100 [BD Biosciences]). A species-appropriate biotinylated secondary antibody (1:100; JacksonImmuno, West Grove, PA) was used to detect the primary antibody, and an avidin-binding complex and AEC detection kit used for color development (Vector Laboratories, Burlingame, CA).
Vessel density and microvascular morphometry was done by manual counting of a mid-graft tissue sample by an observer (B.R.S.) blinded to the experimental protocol. Vascular profiles were characterized by positive staining with human or mouse CD31, or anti-Bcl-2 staining, in structures with an identifiable vascular lumen greater than 5 μm. All vessels in 10 random, high-power fields (54 × 54 μm) were counted for each graft to determine vessel density. In some experiments, all vessels within five 200 × fields were enumerated and/or characterized. Statistical analysis was performed using an ANOVA with Bonferroni post hoc correction.
Results
As described previously, Bcl-2-HUVEC suspended in simple protein gels may be cultured for 7 days or more without significant loss of viability, forming a network of simple cords and tubes. 6 However, culture of HASMC in simple protein gels, with or without Bcl-2-HUVEC coculture, results in contraction of the gel, evident by 24 h and pronounced by 48 h (Fig. 1a, b). Gel contraction continues for at least 72 h of culture, resulting in a dense ball of collagen (data not shown). Such contracted gels have proven unsuitable for implantation. By contrast, when a degradable, nonwoven PGA scaffold is incorporated at the time of gel polymerization (Fig. 1c), the collagen gel remains stably attached to the tissue culture well edges throughout the same time period (Fig. 1d). The presence of PGA does not adversely affect the viability of Bcl-2-HUVEC or HASMC, incorporated separately or in combination (Fig. 2A–C). Based on these observations, we proceeded to implant PGA-supported gels containing human vascular cells to assess their in vivo behaviors.

Effects of nonwoven PGA scaffold support on 3D in vitro culture of vascular cells. Coengraftment of Bcl-2-HUVEC and HASMC in collagen-fibronectin gels results in initial gel contraction by 24 h (

Effects of PGA on cell viability in 3D in vitro culture. The presence of the PGA scaffold does not reduce the number of viable cells in 3D cultures of Bcl-2-HUVEC or HASMC and significantly increased viability in Bcl-2-HUVEC–HASMC cocultures (
In a small pilot experiment, we observed that PGA-supported protein gels containing both HASMC and Bcl-2-HUVEC developed many more large caliber microvessels than PGA-supported gels containing Bcl-2-EC alone. We then set out to confirm this effect in a larger experiment. PGA-supported protein gels were implanted in a subcutaneous position in the abdominal wall of C.B-17 SCID/bg mice using the same technique as has been previously described for unsupported protein gels.6,12 Constructs were implanted that contained no vascular cells, Bcl-2-HUVEC only, HASMC only, or Bcl-2-HUVEC with HASMC. Grafts were explanted for evaluation at 15, 30, or 60 days. By gross examination, acellular gels supported by the degradable PGA scaffold appeared to be only minimally perfused at 30 days postimplantation (Fig. 3A). PGA-supported protein gels containing EGFP-transduced HASMC appeared somewhat more extensively perfused than acellular PGA-supported gels at the same time points (Fig. 3B). We reasoned that the observed angiogenesis might have resulted from a foreign body reaction. Immunohistochemical analysis of constructs harvested at 15 days confirmed the presence of a mouse macrophage response to the PGA, characterized by Mac-3–positive mononuclear and multinucleated foreign body giant cells (Fig. 3C). Such responses were not seen in our previous experiments using acellular protein gels lacking a PGA scaffold.6,12 Thus, it seems likely that PGA fibers induce a foreign body reaction, and that mouse macrophages participating in this reaction induce an angiogenic response.22–24 As detected by immunohistochemical staining of tissue sections from PGA-supported gels containing HASMC only, some EGFP-positive HASMC were seen to be closely associated with the fibers of the scaffold while other HASMC incorporated into the walls of host-derived microvessels (Fig. 3D).
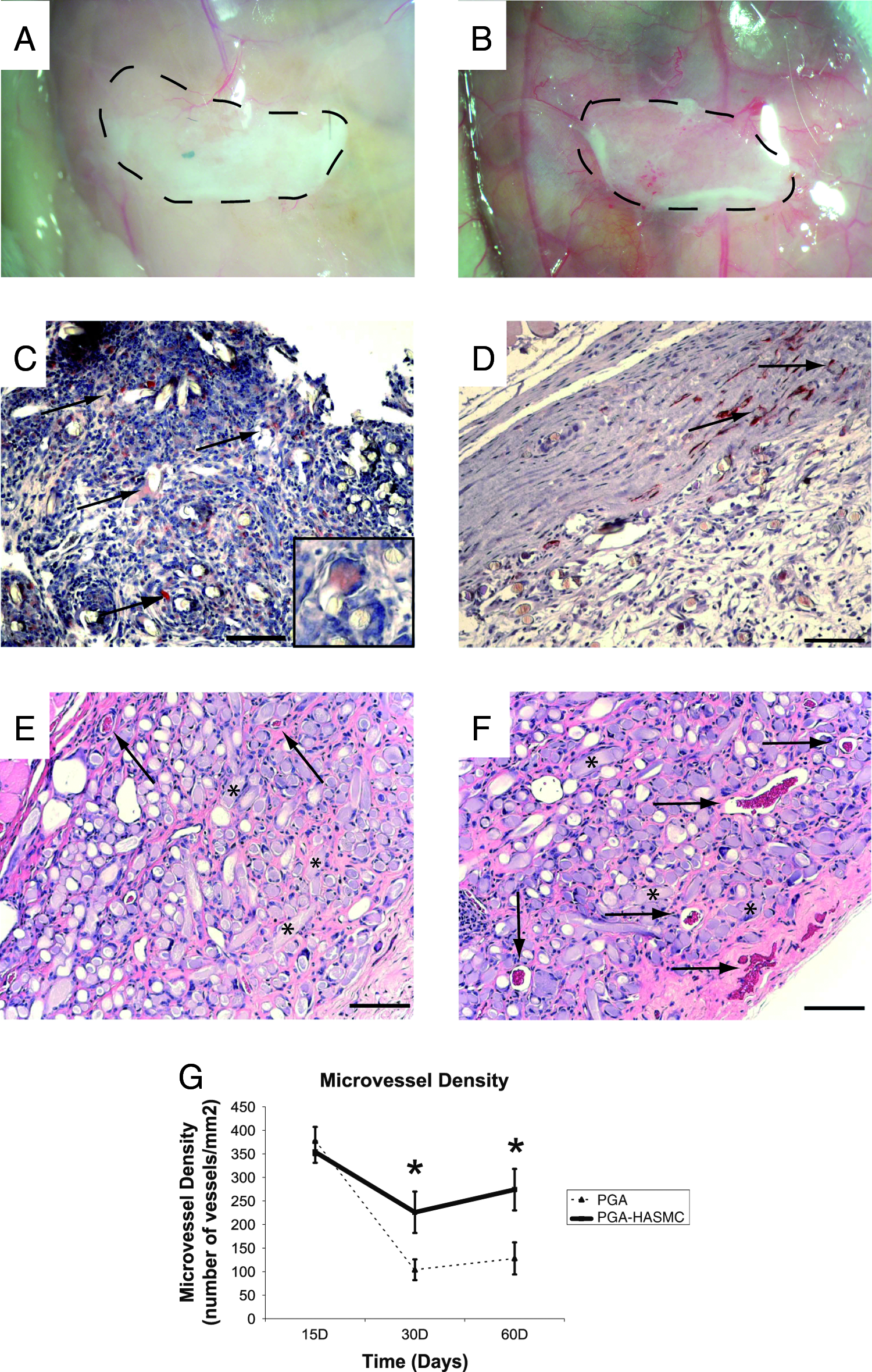
Subcutaneous implantation of acellular and PGA-supported gels containing HASMC. Acellular gels containing PGA scaffold material appeared to contain very few host-derived vessels after 30 days implantation by gross examination (
H&E staining of formalin-fixed, paraffin-embedded tissue sections of acellular PGA-supported gels revealed the presence of the PGA scaffold material at 30 days postimplantation within a somewhat vascularized collagen/fibronectin matrix (Fig. 3E; arrows identify microvessels, and asterisks indicate PGA fiber remnants). In comparison to acellular PGA-supported protein gels, histologic evaluation of HASMC-containing PGA-supported protein gels confirmed the findings of gross examination and revealed a greater number of host-derived microvessels at 30 days postimplantation (Fig. 3F; arrows identify microvessels, and asterisks indicate PGA fiber remnants).
The ability of transplanted HASMC to increase vessel number at 30 days could arise from further stimulation of host angiogenesis or from prevention of vessel regression, or both. To address this issue, we compared acellular grafts with grafts containing only HASMC at various times. Quantitation of vessel profiles within the protein gel matrices revealed no effect of HASMC on vessel number at 15 days. However, PGA-supported gels containing HASMC maintained the number of host-derived microvessels present in the gel at 30 and 60 days, whereas a significantly reduced number of microvessels were present in acellular gels at later time points (Fig. 3G). These data favor the interpretation that the major effect of HASMC on host-derived microvessels, in this setting, is stabilization.
In general, the degree of perfusion of PGA-supported gels that lack EC, with or without HASMC, was slight compared with our previous experience with Bcl-2-HUVEC in unsupported protein gels. PGA-supported grafts containing Bcl-2-HUVEC or Bcl-2-HUVEC combined with HASMC grossly appeared to be much more vascularized as early as 15 days postimplantation (data not shown) with robust vascularization at 30 days. Such gels were obviously more well perfused than either acellular constructs or constructs containing only HASMC.
By histology, the number of microvessels present within PGA-supported gels containing Bcl-2-HUVEC versus Bcl-2-HUVEC with HASMC was indistinguishable at 15 days (data not shown). After 30 days of implantation, there were microvessels present throughout the collagen/fibronectin gel matrix in PGA-supported gels containing Bcl-2-HUVEC alone (Fig. 4A). Following 60 days of implantation, in PGA-supported gels containing Bcl-2-HUVEC alone, there was complete PGA scaffold degradation and persistent human microvessels, many invested with host-derived mural cells, present throughout the remaining protein gel (Fig. 4B).
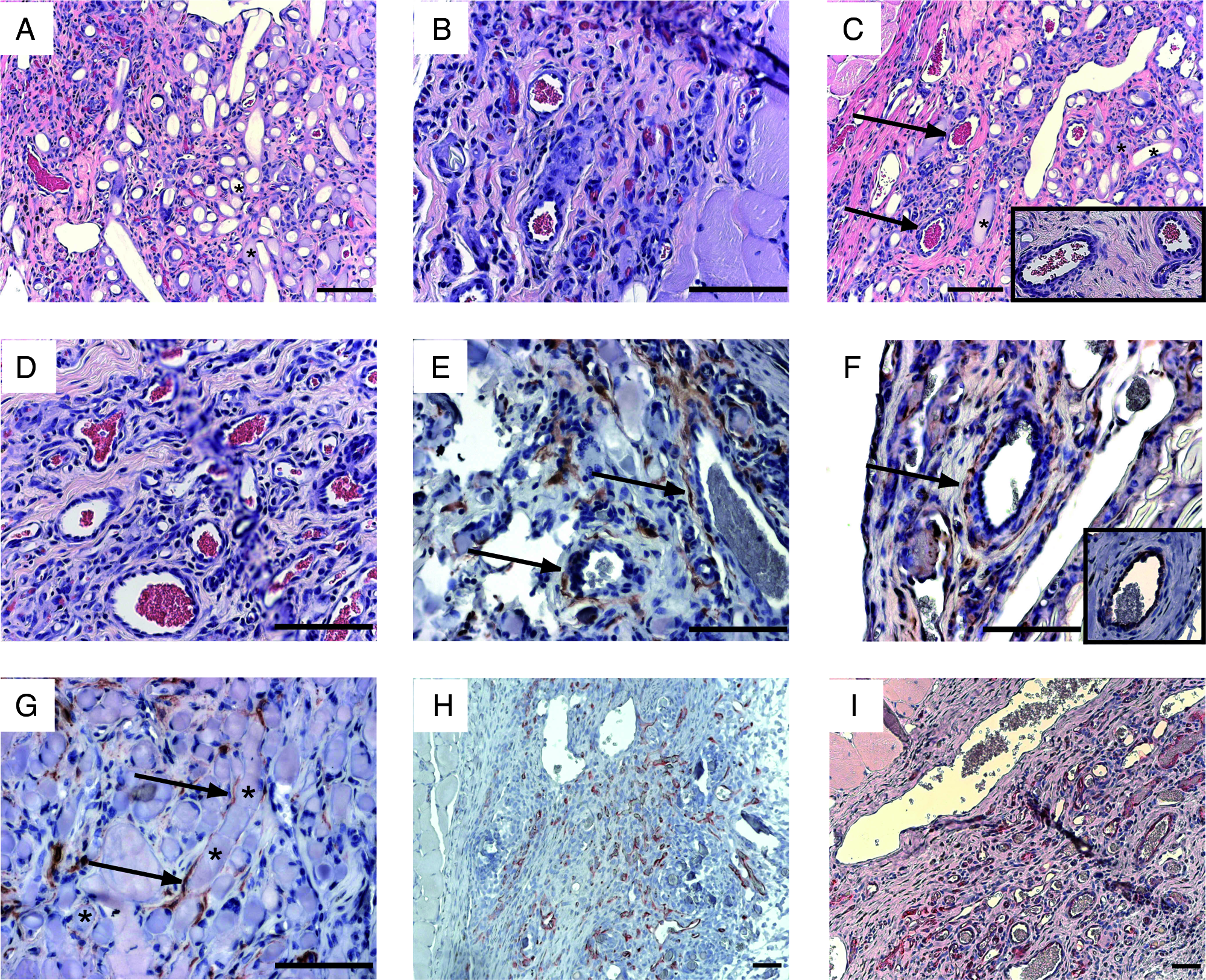
Morphologic and immunohistochemical analysis of PGA-supported gels containing Bcl-2-HUVEC and/or HASMC. H&E staining of tissue sections from PGA-supported gels containing Bcl-2-HUVEC alone following 30 days implantation illustrates partial scaffold degradation and robust vascularization (
When compared with PGA-supported gels containing Bcl-2-HUVEC alone, microvessels in the Bcl-2-HUVEC–HASMC coengrafted gels showed more extensive SMC investment of arteriole-like structures at 15 days (data not shown), 30 days (Fig. 4C, arrows, panel inset), and 60 days. After 60 days of implantation, when compared with PGA-supported gels containing Bcl-2-HUVEC alone, H&E-stained sections of PGA-supported gels containing coengrafted Bcl-2-HUVEC and HASMC (Fig. 4D) revealed the presence of many more large caliber microvessels within the protein gel constructs.
Morphometric analysis and quantitation of the vessels present within the PGA-supported gels containing either HUVEC or coengrafted HUVEC and HASMC revealed that there were no significant differences in the size distribution of the microvessels present within the gels at 15 or 30 days (Fig. 5A, B). The total microvessel density within the PGA-supported collagen/fibronectin gels was also similar at 15 and 30 days for Bcl-2-HUVEC and Bcl-2-HUVEC–HASMC–containing gels (Fig. 5D).

Morphometric analysis of microvessels present within PGA-supported protein gels. While vessel size distribution within grafts containing either Bcl-2-HUVEC or Bcl-2-HUVEC and HASMC was similar at 15 days (
As we predicted from our pilot experiment, there was a significant increase in the number of vessels larger than 50 μm (2.6 ± 0.5 vs. 1.3 ± 0.2), in the HUVEC–HASMC coengrafted gels when compared to the PGA-supported gels containing HUVEC alone at 60 days postimplantation (Fig. 5C). The total in-graft microvessel density was also observed to be significantly greater in HASMC-containing PGA-supported gels at 60 days when compared with PGA-supported gels containing only Bcl-2-HUVEC (655 ± 60 vessels/mm2 vs. 450 ± 76 vessels/mm2) (Fig. 5D). Human EC-lined microvessels appeared to be increased in number at 60 days, but this change was not statistically significant (Fig. 5E).
Immunohistochemical evaluation of PGA-supported gels containing both Bcl-2-transduced HUVEC and HASMC revealed the integration of transplanted HASMC in the walls of arteriole-like and venule-like vascular structures within the implanted protein gel (Fig. 4E). After 60 days, EGFP-positive HASMC could be located in the media of well-formed arterioles within the protein gel (Fig. 4F). Immunohistochemical staining of serial tissue sections with an antibody specific for Bcl-2 demonstrated that the arteriole containing EGFP-positive HASMC was lined by Bcl-2–positive EC (Fig. 4F, panel inset). As was the case in gels lacking EC, in PGA-supported gels containing coengrafted Bcl-2-HUVEC and HASMC, some engrafted HASMC could also be found tightly associated with the polymer fibers of the PGA scaffold; EGFP-positive cells were identified wrapping around and running along single fibers throughout the graft (Fig. 4G). Anti-Bcl-2 staining of formalin-fixed, paraffin-embedded tissue sections revealed extensive microvascular remodeling of Bcl-2-HUVEC–lined (human) microvessels from postimplant day 15 (Fig. 4H) to day 60 (Fig. 4I). This microvascular remodeling was characterized by development of large dilated venules and smaller arteriolar structures with concentric layers of mural cells. These results indicate that the process of polymer degradation did not negatively affect the development or persistence of human microvessels within the protein gel matrix.
The number of α-SMA–positive microvessels was also found to be significantly greater at 15, 30, and 60 days postimplantation in the Bcl-2-HUVEC–HASMC coengrafted protein gels when compared to protein gels containing Bcl-2-HUVEC alone, suggesting an increased presence of arteriolar structures and increased remodeling kinetics within the PGA-supported protein gels coengrafted with both HUVEC and HASMC (Fig. 5F). Cumulatively, these data show that cotransplantation of HASMC along with Bcl-2-HUVEC both accelerates development of SMC-invested arterioles and promotes formation of large caliber microvessels.
Discussion
We describe a system for the successful coculture and cotransplantation of EC and SMC in 3D collagen/fibronectin gels supported by a nonwoven, degradable PGA matrix. The increased mechanical strength imparted by incorporating a degradable scaffold material at the time of collagen gel polymerization allowed for protein gel integrity sufficient for cell survival in 3D culture—an obstacle that has previously hindered the simultaneous study of EC and SMC in 3D gel systems. 22 The contractile nature of SMC has also generally prevented their inclusion in long-term tissue-engineered microvascular networks based in simple protein gel systems, as the tension generated by the engrafted SMC on the collagen fibrils of the gel leads to gel contraction and cell death.23,24 This problem, too, was overcome by incorporation of a nonwoven PGA scaffold at the time of cell suspension and gel polymerization. In other words, use of a PGA scaffold prevents gel contraction, increases in vitro cell viability, and allows for the coengraftment of human EC and SMC, allowing their effects on microvessel formation to be studied in vivo. Using this system, we show that transplanted SMC associate with EC-lined microvessels, enhance arteriolization, and increase the formation of large caliber microvessels.
As noted above, the presence of a PGA scaffold within cell-containing collagen/fibronectin gels led to increased cell survival and viability for Bcl-2-HUVEC alone, HASMC alone, or Bcl-2-HUVEC with HASMC. While this result was, perhaps, predictable in the case of HASMC-containing gels, where the scaffold reduced gel contraction, this was an unexpected observation in gels containing only EC. The basis of this effect is uncertain, but it is encouraging for the use of PGA in tissue-engineered constructs. To our knowledge, incorporation of a supporting scaffold material within a 3D collagen/fibronectin gel has not been previously reported to increase EC viability in vitro.
PGA-supported protein gels display properties in vivo not observed in unsupported protein gels of the same composition.6,12,25,26 This is likely due to the inflammatory nature of the incorporated PGA scaffold utilized in these experiments. The inflammatory effects of PGA are well described,15,19,20,27 and an infiltration of Mac-3–positive mouse macrophages was observed localized around the fibers of the PGA scaffold. Macrophages are likely to contribute to the host angiogenic response observed, and while the initial host angiogenesis is likely due, in large part, to the response of the foreign material, it is clear that the presence of HASMC within the collagen/fibronectin matrix alters the phenotype of these microvessels, rendering them, at least temporarily, more stable and preventing angioregression as observed in acellular grafts containing PGA. 28 In other words, the presence of HASMC in collagen/fibronectin supported by a PGA scaffold leads to host vessel persistence within the gel that is not observed in the absence of HASMC.
We are not the first investigators to use synthetic polymers to support protein gels for EC transplantation. Microvessel formation by Bcl-2–transduced human dermal microvascular EC in Matrigel™ has been described using porous poly(lactic acid) sponges.5,29 However, in these experiments, vessel formation was only assessed at relatively early time points (7–21 days), and neither long-term vessel stabilization nor persistence was examined. Further, these experiments did not include contractile cells in the implants, so vessel maturation was dependent on recruitment of host-derived mesenchymal/mural cells. We believe that ours is the first report to employ such scaffolds for the specific purpose of implanting human vascular SMC.
The presence of HASMC within PGA-supported protein gel matrices resulted in the presence of more microvascular structures in gels containing HASMC alone, as well as in gels that contained coengrafted HASMC and Bcl-2-HUVEC. While, we feel it is likely that the presence of HASMC led to a stabilization and remodeling of the microvessels that develop within the protein gel, our data are also consistent with the possibility that HASMC continually induce new vessel growth (angiogenesis), even while the process of angioregression occurs. Thus, it is possible that gels bearing HASMC contain more microvessels due to this persistent generation of new microvascular structures that balance the rate of spontaneous regression.
The inclusion of HASMC with Bcl-2-HUVEC led to the development of a phenotypically unique microvascular network, characterized by the increased presence of large caliber microvessels with a more arteriole-like phenotype. The population of mural cells used in these experiments is an important consideration. Vascular SMC are heterogenous and also differ from pericytes associated with capillaries. Our experiments utilized cells isolated from the media of the thoracic aorta, a population of cells believed to arise from the neural crest.30–32 It remains to be seen whether other mural cells, such as SMC from other arterial locations or pericytes, isolated from different tissue sites, would have the same phenotypic effects on developing tissue-engineered microvessels as those isolated from the ascending thoracic aorta. For example, pericytes from continuous capillaries might favor the formation of continuous capillaries, whereas specialized pericytes, such as Ito cells of the liver or mesangial cells of the renal glomerulus, might induce the formation of specialized capillaries characteristic of these organs.8,33–37 Coengraftment of EC with other such specialized mural cells may provide for much more intricate control of directed angiogenesis or directed self-assembly of tissue-specific microvascular networks, a process we have termed microvasculogenesis.
In summary, we have developed a new system of PGA scaffold–supported collagen/fibronectin protein gels for coengraftment of EC and contractile vascular SMC. These studies offer proof-of-principle that vascular SMC can accelerate, stabilize, and promote remodeling of tissue-engineered microvessels. Our system opens the possibility of directed microvasculogenesis through coengraftment of EC with selected populations of vascular mural cells.
Footnotes
Acknowledgments
The authors would like to thank Louise Benson, Gwendolyn Davis, and Lisa Gras for their help in isolation of HUVEC and animal husbandry.
This work was supported by grants NIH-RO1-HL085416 (B.R.S., S.M.J., W.M.S., and J.S.P.) and NIH-PO1-HL70295 (G.T. and J.S.P.)