
Abstract
Abstract
Currently, most of the research on how to encourage stem cells to replace missing tissues focuses on biochemical control, such as signaling by growth factors. In addition to basic questions, such as how are stem cells induced to differentiate into particular cell types, also inherent in those studies are practical questions about how to identify, grow, induce, and safely deliver stems cells to the proper target. At the Forsyth Center for Regenerative and Developmental Biology, we are examining a different set of signals, specifically bioelectric signals (the regulated movement of ions across membranes), including membrane voltage, pH, and gap junction activity and gating. We have found strong evidence that bioelectrical signals function at many critical, early points, both up- and downstream of transcriptional regulation, during the processes of normal morphogenesis and adult stem cell–based regeneration. Examples described include gap-junction-dependent regulation of stem cell identity in a flatworm, proton-flux-regulated establishment of left-right asymmetry in vertebrates, and proton-flux-initiated regeneration of a complex structure that includes spinal cord—the tadpole tail—in frogs.
Introduction
When I found so astonishing a power placed within my hands, I hesitated a long time concerning the manner in which I should employ it. Although I possessed the capacity of bestowing animation, yet to prepare a frame for the reception of it, with all its intricacies of fibres, muscles, and veins, still remained a work of inconceivable difficulty and labour.
Bioelectricity is a familiar process, at least in the context of nerve conduction and control of heartbeats; EEGs and EKGs are records of bioelectrical phenomena, but all cells generate and use bioelectricity for a variety of tasks, including “housekeeping” functions and processes related to the differentiated state of the cell. This ubiquitous bioelectricity occurs over much longer time scales than the above two examples of action potentials, but also results from the regulated movement of ions in and out of the plasma membrane. Ion translocators, including channels and pumps, both of which are made of protein subunits, make that movement possible. When a charged element such as an ion moves, that is a current; when charges are kept separate, that is a potential, called voltage. The term bioelectricity encompasses all of these biologically generated electrical phenomena.
The study of bioelectrical phenomena in development has a long history (see review in note 2). Particularly relevant to the work presented here are studies of cancer, regeneration, and embryonic patterning.3–9 The following is a review of three published studies10–12 that build on the work of those pioneers by using modern molecular techniques to show how bioelectricity is a fundamental component of a variety of developmental processes, including control of stem cell differentiation, patterning of a vertebrate body axis, and initiation of regeneration. I will use these examples as illustrations of the contention that research on this other aspect of cell physiology promises to provide insights equal in consequence to those coming from the study of cell and tissue biochemistry.
Bioelectricity and the Differentiation of Stem Cells
The flatworm is a bilaterally symmetrical animal with three tissue layers, multiple organ systems, and a complex brain and central nervous system. It also possesses impressive powers of regeneration; 13 a worm can be cut into many, many pieces, and each piece can regenerate into a complete worm. The basis of this ability is the presence, throughout the animal, of adult stem cells called neoblasts. Like all stem cells, their ability to divide and give rise to two different kinds of daughter cells (another stem cell, and a cell that will now differentiate into a particular cell type) distinguishes them. The process by which these cells replace missing tissue begins when the stem cell receives an as-yet-unidentified signal to proliferate. Once in the new location, the non-stem daughter cells then begin the process of differentiation into replacement tissue.
The significance of this ability to regenerate goes beyond maintenance and tissue repair, however, because the worms use the same mechanism for asexual reproduction. Flatworms can reproduce by pinching themselves in half to produce two partial worms, an anterior half and a posterior half. Stem cells in both halves are then recruited to regenerate whichever half is missing; the neoblasts of the anterior half regenerate a tail, whereas the neoblasts of the posterior half regenerate a head. In the same way, if the middle trunk section of a worm loses head and tail, the stem cells will regenerate both, each at its appropriate end (Fig. 1). To achieve the correct pattern, cells at the cut edge must receive information from that portion of worm to which they are attached, otherwise they could regenerate the wrong end. This passing of information cannot be a local phenomenon because the same cells, if they are next to a posterior cut edge, signal the edge cells to make tail, but if they are next to an anterior cut edge, they will signal the edge cells to make a head. Thus, the information that patterns the regenerating cells must be a long-range signal. Because of this requirement, Levin and colleagues hypothesized that gap junctional communication (GJC), which can carry long-range information, might play an essential role in the ability of worms to regenerate. 14

Cartoon illustrating the process of regeneration of flatworm pieces. Stem cells, called neoblasts, proliferate and regenerate the missing tissue, correctly putting the head at the anterior end and the tail at the posterior, regardless of where the original piece comes from.
Gap junctions are like tunnels connecting the cytoplasm of adjacent cells. 15 When open, they allow passage of ions and small molecules between the two cells; thus, they form a conduit that results in the two cells sharing bioelectrical, and some biochemical, properties. In addition, membrane charges affect the open–closed state of a gap junction; thus, the electrical state of the adjoining cells affect GJC even in situations when current does not carry the signal.
Reagents that inhibit GJC indicate that GJC is important for planarian regeneration, but this must be confirmed using the high level of specificity available with molecular genetics. One way to achieve protein-specific loss of function is RNAi. RNAi is a nucleic acid that, because of a complimentary sequence of nucleotides, can interact directly with the native mRNA for the protein of interest, a process that leads to the degradation of the native RNA. Once the sequence of the RNA of interest is known, an RNAi can be designed and injected that will cause the destruction of the relevant native mRNA. Thus, RNAi is a highly specific inhibitor.
RNAi experiments in the flatworm Schmidtea mediterranea designed to knock down each type of gap junction individually (there are 12 in the planaria studied10–16 ) showed that it is the gap junction protein smedinx-11 that is required for normal regeneration to take place. The loss-of-function RNAi data, in addition to showing that GJC is required for regeneration, also showed that GJC is required for the stem cell population to maintain itself. These and a variety of other experiments led to the conclusion that the GJC mediated by smedinx-11 is required for the proliferation of the stem cell population and therefore for its maintenance.
GJC in S. mediterranea is also critical for signaling to the stem cells where they are in the animal. As mentioned above, a trunk piece will normally regenerate a head at the end where the head was amputated and a tail at the end where the tail was amputated (Fig. 1). This means that there is a signal that tells the stem cells near the head end that they are at the anterior of the trunk and therefore should regenerate the anterior tissues. Another signal tells the posteriorly located stem cells to regenerate posterior tissue. The correct pattering of this signaling, that is, the correct distribution of information about the anterior–posterior (AP) axis of the worm, is also dependent on GJC. This was shown by inhibiting all GJC, through addition of heptanol to the water in which the trunk pieces were regenerating their heads and tails. Under these conditions, a significant number of worms regenerate with heads at both ends (Fig. 2). The interpretation of that result is that GJC is part of the signaling that prescribes posterior structures, because in its absence, anterior structures develop at both ends. Thus, GJC is involved not only in the regulation of stem cells, but also in the process of patterning the entire organism along its AP axis.
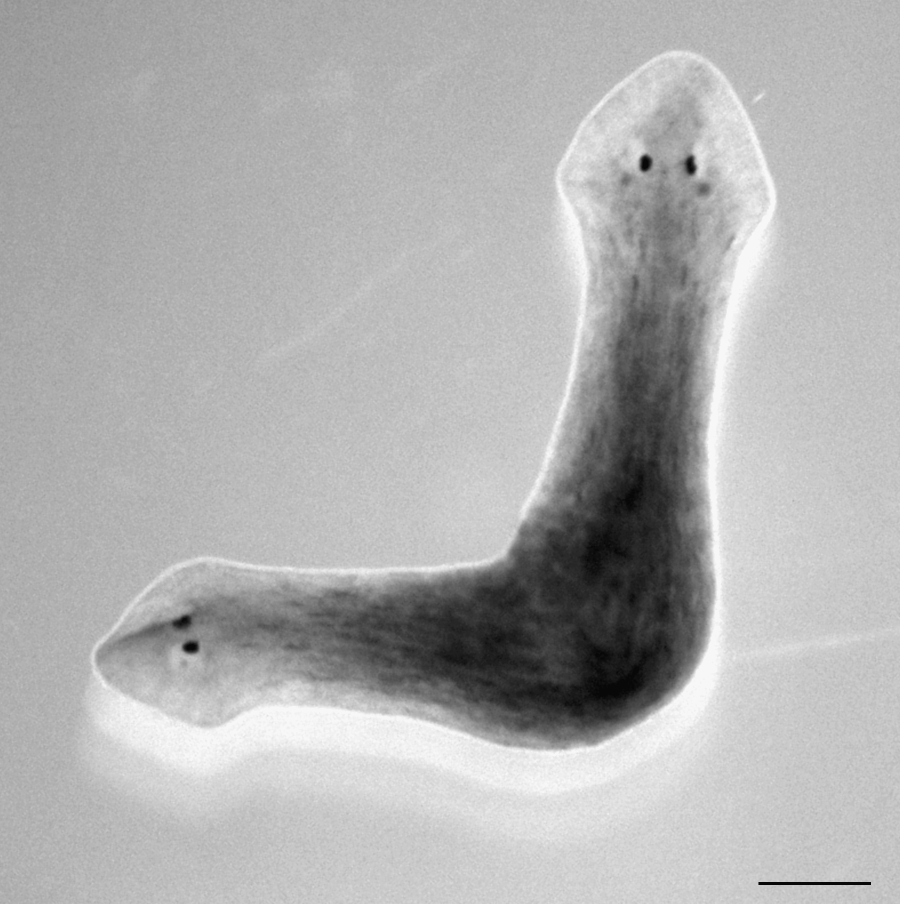
Photo of two-headed Schmidtea mediterranea induced by adding heptanol to the water during the regeneration of the trunk piece. This inhibition of gap junctional communication has re-specified the fate of cells. Scale bar =500 μm.
These data reveal that bioelectrical or very small molecule–based communication between stem cells and the cells that surround them may prove to be an essential aspect of stem cell biology, especially relevant to engineers because, as indicated in the Introduction, control of stem cell proliferation and differentiation are high-priority goals. Understanding of the in vivo gap junction-mediated bioelectrical signaling involved in these processes will surely provide important and practical information for those seeking to mimic the physiology and morphogenesis of stem cell–based tissues. For example, GJC could be used to control communication between cells or to rationally establish fields of equalized membrane potential, nutrients, and other small molecules.
Bioelectricity and the Orientation of the Heart
Like the development of the AP polarity in worms, establishment of the other two axes, dorsal–ventral (DV) and left–right (LR), is an example of what developmental biologists call patterning, and like the study of stem cells, research on patterning is focused primarily on the role of chemical signaling factors. We have found that, in this context as well, bioelectrical events play pivotal roles in morphogenesis, specifically, the creation of biased LR asymmetry of vertebrates.
Vertebrates, such as humans, are bilaterally symmetrical, a term that probably derives from our appearance from the outside (e.g., two eyes, two arms, two legs). However, on the inside, there is striking asymmetry to our organs; usually, our hearts and spleens are on the left, the two lobes of our livers are subdivided along different planes, and even our brains are sided. 17 This asymmetry is found in all bilaterally ‘symmetric’ animals, but what is more interesting is that, in most cases, this asymmetry is biased, that is, it is always aimed in the same direction. In other words, we are not a 50-50 mix of left hearts and right hearts. This bias is difficult to explain in terms of functionality, because mirror-image reversals occur, a phenomenon called situs inversus, and those animals appear to function normally. It is also difficult to explain developmentally. Brown and Wolpert proposed one well-accepted model for how the bias might develop. 18 Their idea is that two axes of a chiral (handed) molecule—they called it the F molecule—could be aligned with the AP and DV axes, and that would force the third axis of the molecule to point in one direction along the LR axis. This accomplishes the task of biasing asymmetry. The discovery that situs inversus in mice can result from a mutation in a molecule called dynein (subsequently dubbed left–right dynein, or LRD) supported this model. This led to the finding that LRD is found in cilia, small hairs that spin, and the discovery that cilia with normal LRD spin clockwise. In mice, this clockwise rotation of a particular field of cilia is a critical step in turning the asymmetry of a molecule, LRD, into the biased LR asymmetry of the whole organism. Although this is a fascinating result, ciliary rotation cannot be the basis of LR asymmetry in all vertebrates19,20 simply because, in other animals, such as frogs and fish, LR asymmetry is biased before the appearance of the first cilia, whereas other asymmetric organisms (e.g., Drosophila) do not have these cilia at all. Our study of the creation of LR asymmetry in frogs, fish, and chicks led us to propose a complimentary model based on asymmetric localization of bioelectrical events in the early embryo.
We know that patterning of the Xenopus embryo cannot be the result of transcription factor activity because transcription does not begin until later, during cleavage; that is, at the time of AP, DV, and LR axis determination, there is no transcription, and maternal RNAs and proteins carry out all processes. These patterning events are epigenetic phenomena. For example, the AP and DV axes are determined relative to the point where the sperm fused with the egg at fertilization. The first cell division, known as the first cleavage, then separates the future left and right sides of the organism. We have found that loss of function of certain bioelectric processes during the earliest cleavage divisions can remove the bias in the asymmetry; that is, they can randomize the sidedness of the bias. The earliest LR difference of a known biochemical signaling molecule does not appear until later in development.
These early bioelectric processes were discovered in a hierarchical drug screen. This kind of screen is an iterative process employing ever-more-specific ion-translocator inhibitors to narrow down the list of candidate fluxes for further studies. By showing that a broadly acting disruptor of the activity of a certain ion does not affect the process, this screen rapidly rules out that ion. A practical guide to this kind of screen is described in note 21. Screens by Levin and coworkers showed that the processes relevant to the early LR biasing event involve the flux of potassium and hydrogen ions across the plasma membrane of early cells.11,22 Two hydrogen-ion pumps, the V-ATPase and the H/K-ATPase, and two potassium-ion channels, KvLQT 23 and Kir4.1, mediate that specific flux. 24 All of these ion translocators are electrogenic; that is, their activity leads to a separation of charge across the membrane. Because positively charged hydrogen ions (H+ or protons) and positively charged potassium ions travel out of the cell, the result is a net positive charge on the outside of the membrane and a net negative charge on the inside. This is a membrane voltage, and by convention, net negative on the inside is considered a negative membrane voltage. Immunohistochemical localization of subunits of these translocators show that they, unlike the vast majority of proteins, can be found in only one cell after the first division. This biased localization is still visible in the four- and the eight-cell embryo, when it can be deduced that they are specifically localized on the future right side (Fig 3). The activity of these pumps has been inferred from direct measurements of proton efflux; in an eight-cell embryo, proton efflux from the cell on the future right-ventral side of the embryo is double the efflux from the left-ventral cell. This difference can be erased if the proton-pumping V-ATPase is inhibited. 11 This is consistent with the above-mentioned localization of the V-ATPase subunits to the future right side of the embryo. These and other data led us to the following model of bioelectric biasing of the LR axis. 25 Localization of proton pumping to the right side of the embryo leads to a difference in membrane voltage across the LR axis; the future right side is more negative (because more protons are leaving those cells), and the future left side is more positive (because fewer protons are leaving those cells). In addition, the excess pumping of H+ ions on the right side leads to cells of higher cytoplasmic pH on the right side. Finally, Levin and Mercola showed that there is an indirect gap-junctional pathway connecting the left-ventral cell to the right-ventral cell that runs through all of the other cells but does not connect those two cells directly. (If it did, there would be no way to establish differences in membrane voltage because of the unfettered movement of ions between the two cells.) 26 Because of that gap junctional path, there is a charge gradient and a pH gradient across all the cells that runs from the most negative right-ventral cell to the most positive left-ventral cell. Our data suggest that both of these gradients are required for normal LR asymmetry. 11

Asymmetric localization of V-ATPase subunits in an eight-cell-stage frog zygote. Shown is a section through an eight-cell Xenopus embryo. The dark color of the right ventral cell is the result of immunohistochemistry using an anti-V-ATPase subunit c antibody and alkaline phosphatase–conjugated secondaries. The staining reveals localization of this subunit to this cell. Scale bar = 250 μm.
A membrane voltage gradient can provide the force needed to separate molecules by their net charge; this is completely analogous to the separation of molecules by the voltage of an electrophoresis apparatus. A small positively charged molecule is moved toward the negative “pole” of the embryo's voltage gradient during the early cleavage stages, and it accumulates exclusively in the right-ventral cells. That molecule is the neurotransmitter serotonin.25,27 Further experiments showed that it is the asymmetry in the localization of serotonin that is dependent on V-ATPase activity. 11 Finally, we hypothesize that it is the high pH of the right-ventral cell that stabilizes the serotonin that, by the 16-cell stage, is gone from all the other cells. Thus, the interpretation of these data is that biased bioelectrical events lead to biased asymmetry in the localization and stabilization of a chemical known to be part of the LR patterning cascade.11,28
The theme that this example illustrates is that large-scale patterning, in this case an entire axis of an embryo, can require regulated ion flux. That knowledge could be useful for tissue engineers working to create non-homogeneous tissues; for example, it is possible that manipulating the electrical environment of a replacement tissue, for example, by inducing electrophoresis using an externally applied voltage, could aid in the process of correctly positioning different components of a tissue within a particular scaffold.
Bioelectricity as a Trigger of Regeneration
Another model for tissue engineering is the process of regeneration. Like the original development of a tissue, understanding of the mechanisms of regeneration could provide crucial clues for the engineer seeking to design and control the creation of replacement tissue. Like studies of development, the search for the chemical cues emitted and perceived by cells, particularly stem cells, dominates studies of regeneration. The roles of growth and transcription factors in particular are the subject of important studies elucidating the pathways that lead to regeneration. Once again, our approach is to study the bioelectrical phenomena associated with this process, and we have found that, like the important events of development described above, important events of regeneration are also bioelectrical. Indeed, our studies indicate that ion flux is sufficient to initiate the process of regeneration, thus revealing the exciting possibility that manipulation of bioelectrical events could stimulate cells to carry on the rest of the regeneration process without the necessity for us to understand other details of the phenomenon, such as upstream genetic signals.
The tail of the tadpole of the frog Xenopus laevis comprises skin, muscle, vasculature, notochord (the “spine”), and nervous system, including spinal cord and peripheral nerves. Until developmental stage 45 (about 7 days at 18°C), and then again after stage 48, the tail will regenerate if it is amputated in the posterior half of its length (Fig. 4A). The process begins with the healing over of the amputation site and the formation of a triangular clump of cells in the center of the healing edge, called the regeneration bud. The new tail then grows out, and the above-listed cell types develop from tissue-specific adult stem cells in the tissue anterior to the cut site. The anterior-most end of the new tail is the most mature, whereas the distal tip retains some of the undifferentiated character of the regeneration bud as it is pushed further away from the tadpole. After 8 to 10 days, the regenerated tail is essentially indistinguishable from the tail of uncut controls. The stages between 45 and 48 are called the refractory period, 29 because if the tail is amputated during this time, it can not regenerate, although normal healing takes place (Fig. 4B). The biological reason for this period is not known, but it corresponds to the time in development when the tadpoles begin to feed.
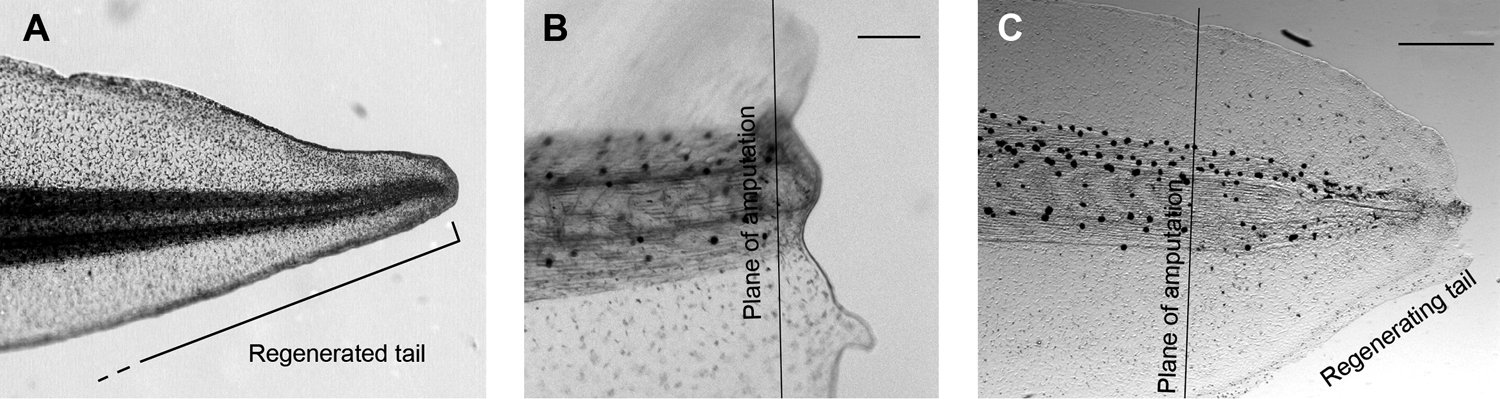
(
By testing a variety of ion transporter inhibitors, it was found 12 that inhibition of the H+-pumping V-ATPase will prevent regeneration of tails, even those cut before the refractory period. This result established V-ATPase as a candidate for an important role in the regeneration process. Subsequent experiments using more-specific molecular genetic inhibitors, such as injection of dominant negative versions of crucial protein components of the V-ATPase, confirmed that the function of V-ATPase is required for normal regeneration to take place. Immunohistochemistry showed that V-ATPase appears in the membranes of cells in the regeneration bud within 6 h of amputation. Thus, this H+ pump is present at the right time and in the right place to play a necessary and direct role early in the regeneration process.
More exciting still are the results of experiments showing that the V-ATPase is not only necessary, but also sufficient to trigger the entire regeneration process. This was done by injecting mRNA for a single subunit, H+ pump from yeast, into the fertilized egg. The tails of the tadpoles that grew from these injected zygotes were then amputated in the presence of the V-ATPase inhibitor concanamycin. Control tadpoles were unable to regenerate because the V-ATPase proton pump was inhibited. In contrast, the yeast proton pump–expressing tadpoles were able to regenerate. This can only be because the activity of the yeast pump allowed the bioelectrical phenomenon—H+ efflux—to take place, and this was sufficient to restore regenerative ability to a majority of treated tadpoles.
Although this result was remarkable, the ability to overcome an applied poison is not the same as the ability to overcome a biologically relevant inhibitory phenomenon that stops a tissue from regenerating. The refractory period is such an inhibitory phenomenon, and it may prove to be a good model of regeneration in humans. This is because many human tissues, such as liver, have regenerative ability; thus, the directions are somewhere in our genes. Even more like the refractory period is the phenomenon of fingertip regeneration. 30 A fingertip, like a tadpole tail, is a complex patterned structure containing different tissues, including skin, skeleton, vasculature, muscle, and nervous tissue. Up to the age of about 7 years, a finger severed distal to the second knuckle can regenerate the lost tissue. However, in older children, as in refractory tadpoles, the regenerative ability is gone. This ability is not often exploited because the wound must be left uncovered for regeneration to take place; however, the standard treatment for a severed finger has been to reduce the risk of infection by covering the wound with skin. Nonetheless, in cases where the wound is kept clean another way, the fingertip can grow back, complete with all its tissues, including a fingernail.*
Coincidentally, one of the contributors to this volume has a regenerated fingertip.
As in the concanamycin experiment described above, mRNA for the yeast proton pump was injected into fertilized eggs, and the embryos were allowed to develop to the refractory stage. Whereas regeneration of untreated controls was inhibited as usual, a significant portion of the yeast proton pump–expressing animals regenerated their tails (Fig. 4C). That dramatic result demonstrated that any proton efflux is sufficient to overcome even a biologically imposed inhibition of regeneration.
These data reveal how state-of-the-art molecular-genetic techniques 11 can be used to exert rational control over bioelectrical signaling in cells. The relevance of these physical control mechanisms for regeneration was demonstrated decades ago,9,31 and this approach is now being extended to modulation of wound healing as well. 32 The implications of the Xenopus tail data are potentially enormous. The process of creating the new tail tissue, with all of its physiological and anatomical complexity, could be left to the cells themselves, without any need for the scientist to carefully manage each step of the growth and differentiation process. The structure itself controlled proliferation, thus preventing tumors; differentiated each tissue, thus correctly signaling the various stem cells; and patterned the structure, thus avoiding teratogenesis. The scientist merely has to hit the correct start button, and the process and all the proper control and regulation of the process are initiated. That the regulatory mechanisms of this regeneration process are stimulated as well leads to the hope that tissues might be engineered without the need for understanding every step. Especially for tissues being engineered in vivo, this could be a tremendous advantage.
Conclusion: One Future for the “Work of Inconceivable Difficulty and Labour”
Proton efflux, or some other bioelectrical phenomenon, may turn out to be the trigger of innate regeneration abilities in humans. Likewise, if bioelectrical activity can regulate large-scale patterning, and gap junctional communication proves to be fundamental to the process of stem cell differentiation, applications abound. Most such applications, however, are far in the future. On the other hand, the data presented here also support the argument that, right now, tissue engineers can gain important insight from considering and experimenting with the bioelectrical environment of the cells used to create tissues. The use of bioelectrical signals gives engineers a new set of tools that influence cell behavior, position, identity, and number; this could lead to an unprecedented level of control over tissue shape and morphology. Already, array analyses of regenerating tissues regularly reveal upregulation of ion transport-relevant genes, and expression profiling has revealed specific bioelectrical signatures for cancer and stem cells. 33 I argue that those targets should be given greater priority for further study.
The illumination of the extremely early role of bioelectricity in these varied developmental processes also supports an argument in favor of seeking control of the mechanisms that will trigger innate developmental mechanisms, rather than mimic later steps in the process. The benefits of such an approach lie in its simplicity and in the possibility that, if tissue development is stimulated at a “natural” initiation point, one may also stimulate innate mechanisms to regulate the process. This was the case with the proton efflux–triggered tadpole tail regeneration; with no a priori knowledge of signaling chemicals, and no effort, time, or money spent to control the myriad biochemical signaling cascades, regeneration of the tail proceeded without error and without side effects. In other words, in response to the surprisingly simple instruction “pump hydrogen ions,” the tissues of the tail were induced to engineer themselves. This paradigm, of creating tissue by triggering rather than mimicking, the mechanisms by which it created itself, could be an exciting addition to the intellectual toolbox available to tissue engineers.
Footnotes
Acknowledgments
The author thanks co-authors and colleagues at the Forsyth Center for Regenerative and Developmental Biology for permission to present their work, especially Michael Levin, who also provided important insights, suggestions, and contributions to the manuscript and to all of the work reviewed. Anonymous reviewers at Tissue Engineering likewise made helpful suggestions. This work was supported by National Institutes of Health Grant 1-K22-DE016633 to DSA.