
Abstract
Despite recent successes in islet transplantation, current immunosuppression protocols required to prevent graft rejection are not suitable for all patients. As a consequence, microencapsulation of islets in alginate has been proposed to protect islets from immune-mediated destruction. Success has been limited, however, due largely to problems with alginate biocompatibility and insufficient immunoprotection by the capsule. The aim of this study was to develop a purified, highly biocompatible, and highly stable alginate from commercially available alginate. We analyzed the chemical properties of the alginate before and after purification and compared in vivo survival and metabolic function of mouse islets encapsulated with either alginate in syngeneic recipients. Recipients of purified alginate capsules exhibited a 105-day graft survival rate of 90.5%, versus 69.2% for recipients of nonpurified alginate, with recipients of purified alginate capsules also showing improved nonfasting blood glucose levels and oral glucose challenges over recipients of nonpurified alginate. On recovery, islets encapsulated in purified alginate capsules demonstrated dramatically reduced capsular overgrowth, and an insulin secretory activity far superior to that of islets in nonpurified alginate capsules. We conclude from this study that alginate purification improves the survival and metabolic function of encapsulated islets. To our knowledge, this is the first paper using pre- and postmodification alginate to demonstrate the direct benefit of purification on transplantation success of islets in simple, open-pore capsules.
Introduction
The objectives of this study were to develop a method to purify and/or modify commercially available alginate to improve the long-term metabolic success of encapsulated islet grafts. Further, as there has been little done in the field to demonstrate the direct effect of alginate purification on transplant success, comparing the outcomes of pre- and postmodification alginate-encapsulated islets in vivo should provide us with greater insight into how alginate could be manipulated to be better suitable for use in transplantation settings. Finally, we had the goal of developing a protocol for alginate purification that could be easily reproduced in any center with readily available materials and minimal technical resources.
Research Design and Methods
Animals
Male BALB/c mice aged 6–10 weeks (HSLAS, Edmonton, Canada) were used as both donors and recipients for transplantation. Diabetes was induced by intraperitoneal injection of streptozotocin (STZ; 275 mg/kg body weight, freshly dissolved in acetate buffer, pH 4.5; Sigma Chemical, St. Louis, MO) 2 days before transplantation. Diabetes was confirmed by the presence of hyperglycemia (≥18.0 mmol/L). Blood was obtained from a snipped tail, and blood glucose levels were monitored with a portable glucose meter (One Touch Ultra; Johnson & Johnson, Milpitas, CA). The animals were kept under conventional conditions in climatized rooms with free access to tap water and standard pelleted food in accordance with the recommendations of the Canadian Council on Animal Care.
Islet isolation
Islets were isolated from male BALB/c mice aged 6–10 weeks according to established protocols. 18 Islets were isolated by collagenase (type V, 1 mg/mL; Sigma) digestion of the pancreas, purified using discontinuous Ficoll (Sigma) density gradients, and hand-picked before culture in Ham's F-10 culture media (Sigma Chemical) in humidified air (5% CO2 and 95% air) at 37°C.
Alginate purification
Before purification, 4% w/v activated charcoal (JT Baker, Phillipsburg, NJ) was bleached in 0.1 mol/L sodium perchlorate (Fisher Scientific, Fair Lawn, NJ) for 30 min, followed with two washes in distilled water and two washes in 95% ethanol. The charcoal was allowed to air-dry overnight. Alginate powder (Pronova UP-MVG; Pronova Biomedical, Oslo, Norway; batch 1 [Pronova 701-256-08] and batch 2 [Pronova FP-301-02]) was dissolved in distilled deionized water (10 g/L). Bleached activated charcoal was added to the solution (20 g/L) and stirred for 3 h. The charcoal/alginate solution was then passed through a succession of filters: first filter paper, and then 1.2, 0.45, 0.22, and finally 0.10 μm filter (Millipore, Mississauga, Canada). The solution was then brought to pH 7.0, after which magnesium chloride (20 g/L) was added and the solution was stirred for 30 min. A solution of 1.7% calcium chloride (26 mL/L) in deionized distilled water was added next and stirred for another 30 min. The precipitated alginate was then recovered and re-dissolved in 0.1 mol/L EDTA + 10 mmol/L HEPES buffer (pH 7) at a concentration 75 mL/g of original alginate dissolved. Once dissolved, 0.125 g NaCl per gram of alginate was added and stirred for 30 min. This solution was then centrifuged at 3000 g using 8000 NMWL Centricon Plus-80 ultra-filtration tubes (Millipore, Bedford, MA), discarding the distillate. Next, 60% ethanol was added to the final solution, refrigerated at 4°C, and allowed to settle for 3 h. The mixture was centrifuged at 2000 rpm at 4°C for 10 min to pellet the alginate. The supernatant was removed, and the precipitate was washed and centrifuged five times more with increasing concentrations of ethanol (60%, 80% × 2, and 100% × 3) as before. To dry the final precipitate, we spread the alginate in a Buchner funnel setup in a biosafety cabinet heated to 35°C, allowing the alginate to dry overnight. Alginate was then prepared as a 1.5% w/w solution (pH 7) in Hank's balanced salt solution (HBSS) without magnesium or calcium (Gibco, Burlington, Canada), filtered through a Gelman “Serum Acrodisc” (Pall Life Sciences, Ann Arbor, MI) 37 mm syringe filter (0.2 μm), and stored at 4°C for later use. Alginates were purified from lots of Pronova UP-MVG to ensure reproducibility of the modification process, though only alginate from the first batch has been used for the islet encapsulation and transplantation studies in this experiment.
Analysis and composition of alginates
The composition of original and purified alginates was determined by measuring the presence of mannuronic (M) and guluronic (G) acid as analyzed by 1 H-nuclear magnetic resonance (NMR) spectroscopy (300 MHz; Bruker DPX, Bruker, Billerica, MA), with fractions of dimers and trimers calculated. 19 Average molecular weights of the alginates were measured by size exclusion chromatography–multiangle laser light scattering (SEC-MALLS). 20 Endotoxin levels in the samples were measured using a Limulus amebocyte lysate (LAL) assay (Chromogenix AB, Mölndal, Sweden). 21
Islet encapsulation
Islets were encapsulated as described previously. 22 Briefly, islets were washed several times with calcium-free and magnesium-free HBSS, centrifuging at 1500 rpm for 1 min between washings. After the final wash, the tissue was centrifuged and the supernatant aspirated. The tissue was then re-suspended into 0.45 mL of HBSS (no Ca++ and Mg++), to which we added 0.45 mL of the 1.5% alginate solution (either Pronova UP-MVG or our purified alginate; both batch 1) and lightly mixed until homogeneous. The alginate–cell suspension mixture (∼10,000 islet equivalents [IE]/mL) was then run through an electrostatic pump generator (Metabolex, Hayward, CA) across a voltage potential into a 120 mmol/L calcium chloride solution causing gelling of the formed alginate beads. These beads were washed several times with HBSS (no Ca++ and Mg++), and then twice with Hams F-10 culture media before plating in Hams F-10 culture media and incubating at 37°C.
Islet transplantation and follow-up
After 24 h culture at 37°C in Hams F-10 culture media, encapsulated islets were washed twice in HBSS, and then suspended in Hams F-10. Before implantation, to standardize the mass of mouse islets transplanted in each recipient, representative aliquots of each preparation were counted and sized to determine total IE. Graft mass varied between 900 and 1200 mouse IE. Encapsulated islets were then implanted into the peritoneal cavity with an 18-gauge cannula via a small incision in the linea alba. The incision was closed using MikRon Autoclip stainless steel wound clips (9 mm; Becton Dickson, Sparks, MD). Mice were followed and monitored for blood glucose levels between 8:00 and 11:00 a.m. When the blood glucose level was ≤ 8.7 mmol/L, the graft was deemed a success. At 105 days posttransplantation, an oral glucose tolerance test (OGTT) was performed on recipients with normalized basal glycemia and in normal controls. After a 2-h fast, D-glucose (3 mg/g body weight) was administered as a 50% solution by oral gavage into nonanesthetized mice. Blood samples were obtained from the tail vein at 0, 15, 30, 60, and 120 min. Total area under the curve (AUC) for glucose clearance was calculated with the trapezoidal rule method for the purposes of comparison of the curves.
At various time points (i.e., >100 days posttransplantation), capsules were recovered by an intraperitoneal lavage with phosphate-buffered saline containing 120 mmol/L CaCl2. From the recovered grafts, an aliquot was taken and stained with dithizone (Sigma) to identify islets. 23 Islets were observed for gross morphology, and capsules were observed for shape, size, and the presence or absence of breakage and/or cellular overgrowth of the capsule on microscopic examination. Hepatic inflammation was categorized as those mice observed to have gross enlargement, discoloration, and hardening of the liver upon capsule recovery. Such observations were recorded as a percentage as an all-or-none phenomenon, with animals showing any hepation inflammation or cellular overgrowth recorded as positive, and animals totally free of the same to be recorded as negative. To confirm the efficacy of the encapsulated islets at correcting diabetes, the pancreas of each recipient was assayed for cellular insulin content as previously described. 24
An aliquot of recovered capsules was also assessed for their glucose-stimulated insulin response using a 2-h static incubation at 37°C. 24 The recovered encapsulated islets were washed twice in HBSS and once in Hams F-10 culture media. The tissue was separated into two samples, each sample incubated in 1.5 mL Hams F-10 supplemented with 2 mmol/L L-glutamine, 0.5% bovine serum albumin, and either 2.8 mmol/L glucose or 20 mmol/L glucose. After the 2-h incubation at 37°C, the supernatant was collected and evaluated for cellular insulin content by radioimmunoassay. The tissue was also collected, and washed twice for 10 min at 37°C with a 0.1% solution of EGTA to dissolve the capsules, after which the tissue was also assayed for total insulin content. The insulin content of the medium was expressed as a percentage of the total content (i.e., tissue plus medium). Stimulation indices were calculated by dividing the amount of insulin release at 20 mmol/L glucose by that released at 2.8 mmol/L glucose.
Statistical analysis
Data are expressed as mean ± SE of n independent observances. Statistical significance of differences was calculated with a two-tailed unpaired Student's t-test, or, in the case of dichotomous data, a Fisher's exact test (chi-squared analysis). Findings were deemed significant at a p-value of less than 0.05.
Results
Alginate purification
Analysis of two lots of alginate by SEC-MALLS demonstrated a reduction in the average molecular weight of the first batch of alginate from 241 to 218 kDa and the second batch from 242 to 220 kDa with purification (Table 1). Endotoxin levels, as measured by an LAL test, were basically unchanged (Table 1). The purification process led to an apparent shift in the composition of the alginate, with the proportion of guluronic acid (FG) in the alginate as measured by 1 H-NMR spectroscopy (Fig. 1) increasing in batch 1 from 68% to 75% and in batch 2 from 68% to 78% (Table 1). Similarly, the fraction of guluronate dimers (FGG) and trimers (FGGG) increased in both instances (Table 1). Overall, the process led to an increase in the average length of guluronic acid blocks from 12 to 15 with the first batch and 14 to 21 with the second batch. The fraction of mannuronic acid (FM) in the alginate conversely decreased with the purification process, along with the proportion of all other blocks analyzed that contained mannuronate.

Composition of batch 2 alginate as determined by 1H-NMR spectroscopy for both unpurified (
FG and FM, fraction of alginate consisting of guluronic and mannuronic acid, respectively; FGG and FGGG, fraction of alginate consisting of guluronic acid in blocks of dimers and trimers, respectively; FMM, the fraction of alginate consisting of mannuronic acid dimers; FMG/FGM, fraction of alginate consisting of mixed sequences of guluronic and mannuronic acid; FMGG/FGGM, fraction of alginate that starts or ends with a block of guluronic acid; FMGM, fraction of alginate consisting of two mannuronic acids interspaced with a guluronic acid; NG>1, average length of guluronic acid blocks; MW, molecular weight.
Transplantation of encapsulated islets
Islets were cultured for 1 day after isolation, encapsulated, and then cultured for 1 day further. Islets were then transplanted into the peritoneal cavity of syngeneic diabetic BALB/c recipients. Animals were followed over time and removed from study when they exhibited two successive blood glucose readings greater than 17 mmol/L. When we plotted our data to observe the metabolic state of our animals over time, we observed that that the average blood glucose concentration of mice receiving islets encapsulated in purified alginate was significantly lower than that of the mice receiving nonpurified alginate capsules (Fig. 2). Further, 90% of mice transplanted with the purified alginate showed graft survival greater than 100 days, with a mean survival time of 139 ± 8 days (Table 2). This is compared to a lower survival rate of 69% and a mean survival times of 118 ± 10 days among recipients transplanted with nonpurified alginate. An OGTT was performed on normoglycemic transplant recipients at approximately 105 days after transplantation (Fig. 3). Animals transplanted with nonpurified alginate–encapsulated islets demonstrated significantly higher blood glucose concentrations throughout the OGTT than those receiving purified alginate (see Fig. 3). Further, the AUC for the OGTT of recipients nonpurified alginate–encapsulated islets was 1994.3 ± 56.1 min·mmol/L, significantly higher than the AUC of 1363.5 ± 52.3 min·mmol/L for those getting purified alginate (p < 0.05).

In vivo function of nonpurified alginate–encapsulated (○), or purified alginate–encapsulated (•) BALB/c mouse islets transplanted into diabetic BALB/c mice. Data are expressed as mean ± SE (p < 0.05). *p < 0.5 versus nonpurified alginate.
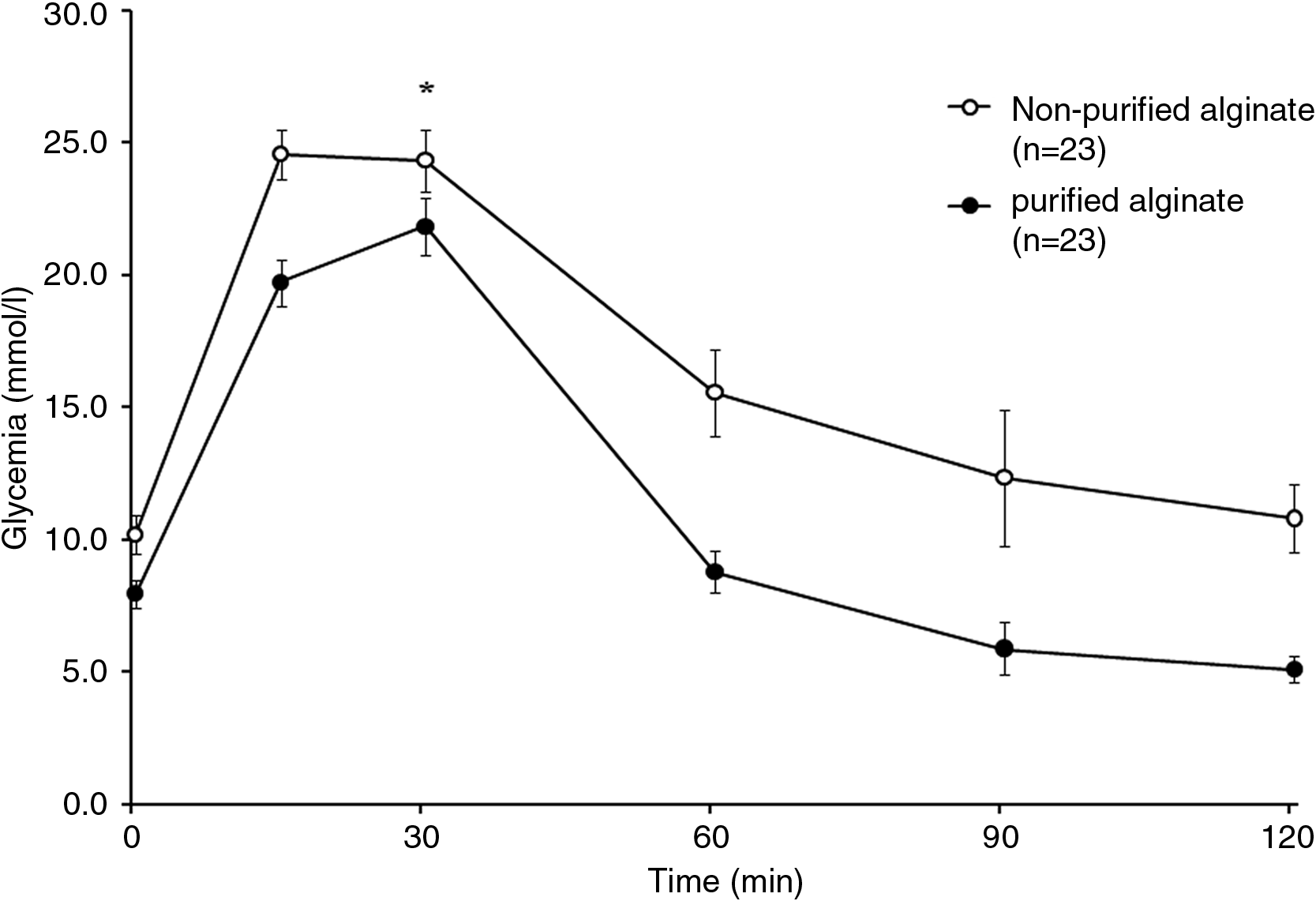
Oral glucose tolerance tests. Blood glucose profile for BALB/c mice with syngeneic transplants of nonpurified alginate–encapsulated (○), or purified alginate–encapsulated (•) islets after 105 days. Data are expressed as mean ± SE (p < 0.05). *p < 0.2 versus nonpurified alginate.
Euglycemia was defined as blood glucose values ≤8.7 mmol/L.
Euthanized while hyperglycemic.
Died while euglycemic.
Euthanized while euglycemic.
Died while hyperglycemic.
p < 0.05 versus nonpurified alginate.
MST, mean survival time.
Recovery of microcapsules
Capsules were recovered at various time points greater than 100 days, at which time the condition of the internal organs of graft recipients was observed. Among mice receiving islets encapsulated in nonpurified alginate, 12 of 27 (44.4%) animals showed signs of severe liver inflammation, compared with 2 of 19 (10.5%) animals receiving islets in purified alginate (Table 3). When we observed the condition of capsules after recovery, animals transplanted with nonpurified alginate–encapsulated islets exhibited cellular overgrowth of the capsule in 20 of 27 cases (74.1%). Such cellular overgrowth was only evident in 3 of 19 cases (15.7%) when purified alginate was used (Table 3). Further, when an aliquot was taken of nonpurified alginate microcapsules covered in cellular overgrowth, capsules contained islets that were dark and necrotic looking and that stained poorly with dithizone (Fig. 4A). On the other hand, samples of nonovergrown, purified alginate microcapsules contained islets that were healthy looking and stained well with dithizone (Fig. 4B). The pancreatic insulin content of all recipients of encapsulated islets was less than 0.9 μg, or less than 3% that of normal control animals, demonstrating that the grafts were responsible for restoration and maintenance of euglycemia in transplanted animals.
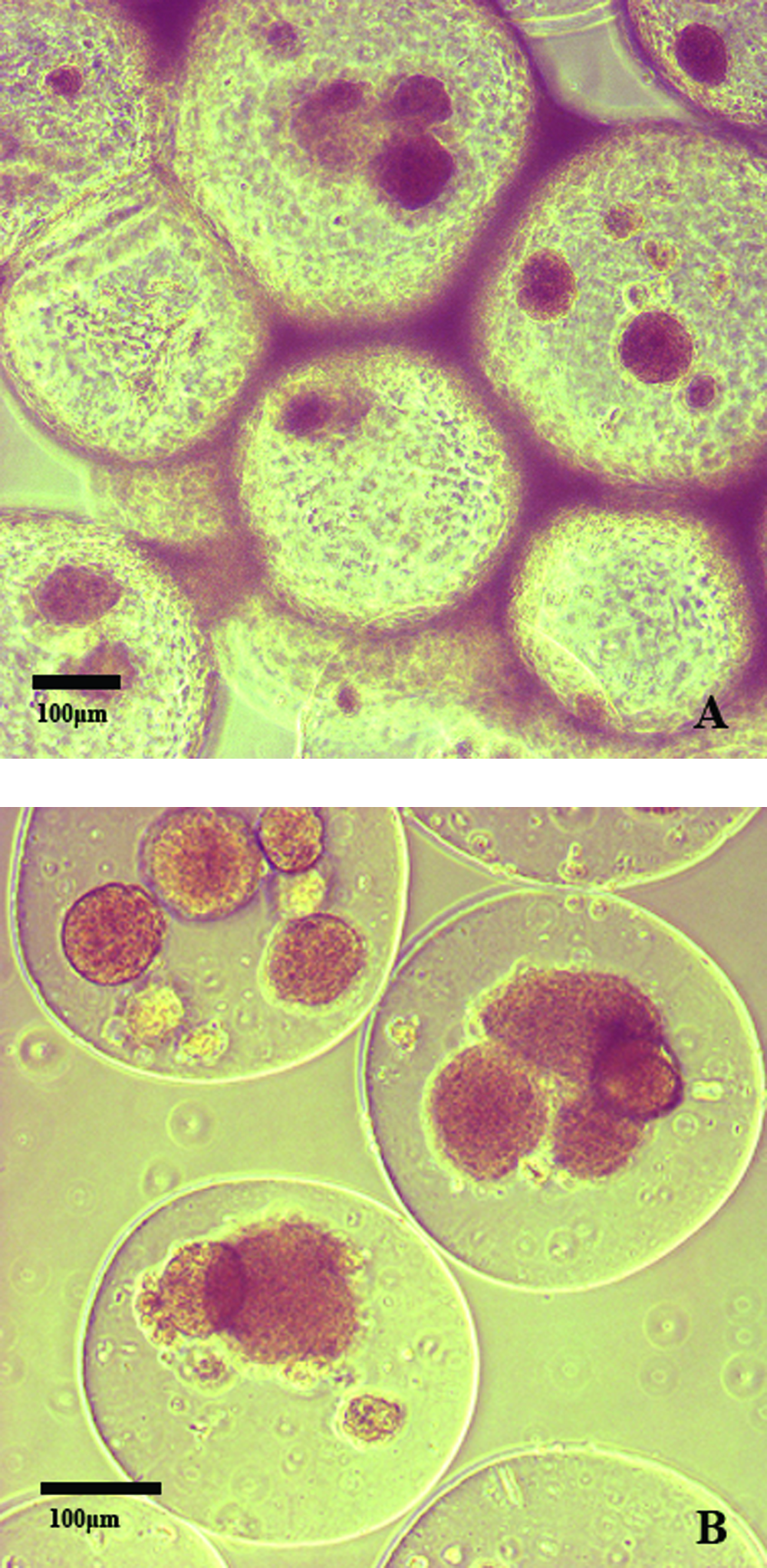
Representative light micrographs of dithizone-stained BALB/c islets encapsulated in nonpurified alginate (
Data are expressed as percentages or as mean ± SE.
p < 0.05.
p < 0.001 versus islets in nonpurified alginate capsule.
Recovered capsules from mice were also assessed for glucose-stimulated insulin release, expressed as a percentage of the total insulin content. Encapsulated islets cultured in the presence of 2.8 mmol/L glucose showed insulin release values of 1.4% and 1.1% of their total cellular content from nonpurified and purified alginate capsules, respectively (Table 3). On the other hand, when cultured in the presence of 20 mmol/L glucose, insulin release values from encapsulate islets were 5.2% and 11.6% from nonpurified and purified alginate capsules, respectively. As a result, nonpurified alginate–encapsulated islets demonstrated an average stimulation index of 2.9 ± 1.1, while purified alginate–encapsulated islets showed a value of 7.9 ± 2.5 (Table 3).
Discussion
The purpose of this study was to determine whether purification of commercially available alginate would lead to an improvement in survival and function of encapsulated syngeneic islets transplanted into an immune-competent mouse model. We have shown previously that encapsulation of islets leads to improved survival and metabolic function in long-term tissue culture compared to nonencapsulated islets, and that these islets demonstrate outstanding function when transplanted into an immune-deficient recipient. 22 With acceptable biocompatibility of the capsules in an immune-compromised recipient, progression to an immune-competent recipient was the next step, not only to assess the capsules for biocompatibility but also for their level of immune protection. 25 Specifically, we wanted to determine if the alginate capsule could sustain islet survival and function in syngeneic recipients as a study of the biocompatibility of the capsule itself. Preliminary work has led us to believe that the commercially available sources of alginate were not ideal (data not shown), and efforts led to an exploration of methods to purify, alter, or optimize the alginate for improved islet survival and function in vivo. Other groups have similarly studied the effects of altering commercially available alginate through enzymatic modification 26 or purification12,27 with varying results, though little has been done to compare pre- and postpurification alginate with respect to its effect on the transplantation success of encapsulated islets.
Alginate modification in our study led to some interesting observations. To begin with, we expected that ultrafiltration against an 8000 NMWL membrane and subsequent removal of very short-chain alginate molecules would lead to an overall increase in the average molecular weight of the final alginate product. Such was not the case. The resultant decrease in the molecular weight of the purified alginate may have been due to degradation of large chains into smaller ones. 28 Further, it is possible that some of the longer alginate chains became trapped in and were not adequately recovered from the Centricon ultrafiltration devices. Although the literature seems to indicate that an alginate of a reduced average molecular size leads to a decrease in stability, 29 our change in molecular weight was relatively small, and led to no apparent loss of stability in the purified capsules. Additionally, it has been recently proposed that removal of monomers and small oligomers of mannuronic and guluronic acid may be beneficial, in that their release from transplanted capsules in vivo may contribute to the immunogenicity of the capsule. 30 We were initially surprised to see that endotoxin levels remained unchanged through the purification process; however, this should not be surprising considering that endotoxin concentrations in commercially available purified alginates are already low (our blank control demonstrated a value of 0.18 IU/mL) and certainly under acceptable transplant limits. 13 Nonetheless, on this point it can be observed that the second lot of alginate demonstrated lower endotoxin levels than the first lot, demonstrating progress made by industry in reducing these levels.
Perhaps most interesting result regarding the alginate purification was the notable increase in the relative guluronic acid (G) content of the postpurification versus prepurification alginate. It has been well documented that alginates of a higher G-block concentration demonstrate greater stability. 29 This is significant because the increase in stability reflects a decrease in the likelihood of capsule breakage, 31 resulting in a reduced incidence of direct exposure of our islets to the immune system. Further, there have been proposed benefits to reducing the M content of alginate capsules, particularly with regard to reducing the immunogenicity of the capsules.32,33 There have been varying opinions over the years as to the preferences of researchers for alginates that are classed as either G rich (higher stability and lower porosity) or M rich (lower stability and higher porosity). Klock et al. indicate that the results of comparisons between alginates are more the result of the processes used to purify the alginate, 34 thus making objective comparison between batches of alginate difficult. Whereas observations seem to suggest that improved graft function and longevity of survival were the result of the increased G content of the purified alginate, it is certainly possible that the improved transplantation success with the purified alginate is the result of the removal of other elements such as proteins or polyphenols14,35 for which we have not tested.
Transplantation of islets into our recipients showed near immediate restoration of normal blood glucose values in most mice, irrespective of whether the islets were encapsulated in purified or nonpurified alginate. Whereas blood glucose values were quite similar for the first week after transplantation, differences between the groups became pronounced and significant from 2 weeks posttransplant and followed consistently through to the end of the study. This was strengthened by our large sample sizes (n = 39 nonpurified alginate capsule recipients; n = 42 purified alginate capsule recipients). Further, blood glucose values of mice transplanted with nonpurified alginate were significantly higher than those of mice transplanted with purified alginate capsules at all points throughout the OGTT. These results could be explained by toxic or other effects of the alginate itself on the islets contained within the microcapsule, though it is more likely that this is a consequence of immune or other forces external to the capsule leading to a loss of viable islet cells within the capsule. Observations of our explanted microcapsules support this thought, as mice receiving islets encapsulated in nonpurified alginate showed a 4.4 times greater incidence of cellular overgrowth on the capsules on recovery than mice receiving purified alginate. We have observed and it has been shown by others that the cells adhering to the surface of the microcapsules are predominantly macrophages.17,36,37 We suspect macrophages to be the predominant cause of the reduced function of the grafts in these recipients by way of a number of possible mechanisms. For example, nutrient or oxygen deficiency within the capsule has been proposed as a potential reason for graft failure with even nonovergrown islets in immune-compromised animals.9,38 As we are using a simple, open-pore capsule of a relatively small size (450 μm diameter), we doubt capsule properties alone to be the lone contributing factor of these potential deficiencies; however, given a “coating” of highly metabolically active macrophages on the surface, it is entirely reasonable that an accumulation of macrophages could lead to a relative deficiency of oxygen and nutrients in the local microenvironment. Further, overgrowth of cells would lead to the local increase in metabolically active waste products of both the macrophages and the islets.
Islet failure may also occur in a more specific manner. Macrophages overgrown on the surface of the capsule may release damaging cytokines, nitric oxide, or reactive oxygen species, causing the death and/or reduced function of the contained islets. Given the simple, open-pore nature of our capsule, these substances, along with any products of islet cell death, could pass freely back and forth across the capsule.39–41 We would expect that this reaction would be primarily mediated via innate immunity because our islets were not transplanted across strain or species, but were syngeneic. The question then arises as to the insult causing such a reaction. More than likely, such a reaction is due to more than one factor, some of which may include the biocompatibility of the alginate, the streptozotocin used to induce diabetes, friction or mechanical forces between the capsule and the liver and peritoneum, or the surgical procedure with subsequent scaring and inflammation. It is interesting to note that in previous studies (data not shown) when empty capsules of both purified and nonpurified alginate were transplanted into immune-competent mice, none of the capsules showed any signs of overgrowth or immune reactivity. This indicates that the alginate itself is not the sole immunogen, and that the presence of the islets, streptozotocin, or other factors was necessary for full immune reactivity. Further, some of the nonspecific causes of islet failure and death could potentially lead to the release of antigens provoking an inflammatory response. It was interesting to note that the predominant organ affected in transplant recipients was the liver, which in 44.4% of mice receiving nonpurified alginate–encapsulated islets capsules and 11.1% of mice getting purified alginate would lose its flat, lobular appearance, and swell up to the appearance of a single spherical mass. In these instances, the capsules were often found clustered around the liver. Combined with the more than fourfold greater frequency of overgrowth in nonpurified versus purified capsules, these results indicate that the composition of the alginate is a key element in this equation. When we compared the insulin responsiveness of encapsulated islets recovered from animals receiving purified versus nonpurified alginate, results were similar to those of the in vivo. Whereas insulin release in the presence of low glucose was similar for both purified and nonpurified alginate–encapsulated islets, culture of purified capsules in a high glucose media yielded more than twice the insulin release of islets in nonpurified capsules. Similar to before, the reduced insulin responsiveness of the islets to glucose can be explained by the loss of functional beta cells over time.
Our findings indicate that the simple protocol for alginate purification demonstrated here reproducibly leads to significantly improved survival and function in encapsulated islets transplanted into immune-competent syngeneic recipients. This leads us to conclude, as others, that alginate composition/purity is essential to the survival and proper function of encapsulated islets in transplantation. Nevertheless, much work yet needs to be done. Graft failure, whether by nonspecific causes, 42 microcapsule material elicitation of a damaging inflammatory response, 43 or other factors, needs to be further analyzed to have a model suitable for human transplantation. Macrophage overgrowth of the capsule is the greatest single factor leading to the failure of our transplants. Along this line of thinking, encapsulation along with macrophage depletion has been reviewed as a possible solution, 17 though the effect of this treatment on the long-term survival of these islets in vivo is uncertain. Whether this mediated failure as a result of nutrient deprivation or specific factors leading to the death of islet cells, or both, is as yet not entirely clear. What is certain is the need to provide continued protection to the islets, particularly as we move toward effective protection in allogeneic and xenogeneic models. This is especially important because macrophages are extremely effective antigen-presenting cells, and could potentially take-up and present shed antigens from the capsule and subsequently activate an even more vigorous innate response. 44 Such a heightened innate response further may act as an adjuvant to a more problematical T-cell–mediated adaptive response present in allogeneic and xenogeneic models. In any case, it is unlikely that encapsulation will provide the entire solution to the problem of graft failure in the absence of immune suppression. Rather, it is more probable that this technology will be one part of a much larger solution.
We feel that the work done here provides a protocol for alginate purification that is readily reproducible and which requires readily available materials and technical equipment. Further, it is evident that the resultant improvement in graft survival and subsequent prolonged and improved restoration of recipients to normoglycemia is worth the time and effort involved in producing this alginate for further use and investigation.
Footnotes
Acknowledgments
Support was provided by the Canadian Diabetes Association, Canadian Institutes of Health Research (Grant #FRN-8030), Alberta Heritage Foundation for Medical Research, and the Juvenile Diabetes Research Foundation. A.G.M. is a recipient of scholarships from the Canadian Institutes of Health Research and the Alberta Heritage Foundation for Medical Research. G.S.K. is a recipient of a Juvenile Diabetes Research Foundation Career Development Award and is an Alberta Heritage Foundation for Medical Research Senior Scholar.
Disclosure Statement
No competing financial interests exist.