
Abstract
Embryonic stem cells (ESCs) can differentiate into all somatic cell types, including cardiomyocytes, which may be used for regenerative cardiac cell therapies. ESCs are commonly differentiated via cell aggregates known as embryoid bodies (EBs), but current cardiomyogenic differentiation methods, such as formation via hanging drops, yield relatively small numbers of EBs and differentiated cells. On the other hand, batch culture methods, like static suspension, yield increased numbers of EBs and cells, but typically exhibit less overall cardiomyogenic differentiation. The objective of this study was to determine if rotary orbital suspension culture, which produces EBs resembling hanging drops, was capable of enhancing cardiomyogenic differentiation compared to static suspension culture. Similar to hanging drops, rotary suspension culture significantly increased the proportion of spontaneously contracting EBs compared to static suspension culture. The gene expression of mesoderm (Brachyury-T) and cardiac transcription factors (Gata4, Nkx2.5, and Mef2c), as well as sarcomeric muscle proteins (α-MHC and MLC-2v) was increased within EBs cultured in rotary suspension conditions. Rotary orbital culture also yielded a greater percentage of EBs that were immunoreactive for α-sarcomeric actin protein compared to static suspension, and augmented the average percentage of α-sarcomeric actin–positive cells detected via flow cytometry. These results demonstrate that rotary orbital suspension culture enhances endogenous cardiomyogenesis of EBs and therefore could benefit the development of regenerative cardiac therapies.
Introduction
Tissue engineering and regenerative cellular therapies capable of replacing cardiomyocytes to injured myocardium offer a promising approach to restore functional tissue; however, an appropriate cell source should have the potential to produce large yields of cardiomyocytes capable of functionally incorporating within the host tissue. Attempts thus far to employ cell therapies to achieve these rigorous criteria have been minimally successful. Skeletal myoblasts and bone marrow–derived cells transplanted within the infarcted heart have elicited small improvements in cardiac function, but have not generated significant amounts of integrated muscle tissue.9–14 The identification and recent characterization of a putative cardiac stem cell population within the heart represents a novel cell source for cardiac regenerative therapies, but inefficient isolation and cultivation methods for these cells currently limit their therapeutic utility.15–18 In contrast to these other cell types, embryonic stem cells (ESCs) are capable of significant expansion in vitro and inherently retain the ability to differentiate into definitive contractile cardiomyocytes that can survive and electromechanically couple with host cells of the heart.19–22 However, before ESC-derived cardiomyocytes can be realistically implemented as a viable cell therapy, efficient methods of differentiating ESCs to cardiomyocytes using scalable techniques must be developed to yield the large numbers of cells (109/heart) expected to be clinically necessary. 23
ESC differentiation is commonly initiated via spontaneous aggregation of single-cell suspensions into clusters of cells termed embryoid bodies (EBs). EBs recapitulate many of the cellular morphogenic events similar to those of preimplantation stage embryos, eventually leading to formation of primitive cell types comprising the three germ lineages (ectoderm, endoderm, and mesoderm) and further differentiation into specialized cells types, including cardiomyocytes.24–29 For experimental studies, hanging drop cultures are typically employed to study mesoderm differentiation because of the high frequency with which cardiogenic differentiation can be readily observed.30,31 However, the number of EBs formed per dish (∼100 per 10 cm plate) is limited by surface area; thus, hanging drops are not readily amenable to scale-up procedures.31–33 In contrast, static suspension cultures yield larger numbers of EBs with less effort than hanging drops, but ESCs in static suspension form a heterogeneous population of EBs that readily agglomerate, thereby further increasing the heterogeneity of differentiation and reducing the overall efficiency of cell differentiation, 34 including the frequency of cardiomyogenic differentiation. Recently, our lab developed a novel EB formation and culture method whereby suspension ESC dishes are placed onto rotary orbital shakers. 35 Compared to hanging drop and static suspension methods, constant rotary orbital motion (40 rpm) applied continuously for 7 days of EB suspension culture enhanced the initial efficiency of cellular incorporation during EB formation and resulted in a significantly greater number of individual EBs and EB-derived cells. Compared to static suspension cultures, rotary orbital shaking yielded more uniformly sized EBs, and agglomeration of individual EBs after formation was inhibited, similar to hanging drop methods. 35
Based on these previous observations between different EB culture methods, the objective of this study was to specifically examine the cardiomyogenic differentiation of EBs cultured in rotary orbital suspension compared to both hanging drop and static suspension cultures. EBs were cultured for 7 days in suspension via rotary, static, or hanging drop conditions, and cardiomyogenic differentiation was assessed based on spontaneous contractile activity, gene expression, and sarcomeric protein expression. The results indicate that rotary orbital suspension culture enhances the endogenous cardiomyogenic differentiation of EBs compared to static batch culture methods. Thus, rotary orbital suspension culture is a simple means of promoting cardiomyocyte differentiation from ESCs for regenerative cardiac therapies.
Materials and Methods
Cell culture
Murine ESCs (line D3) were expanded on 0.1% gelatin-coated (Sigma, St. Louis, MO) tissue culture plates (Corning, Lowell, MA) with Dulbecco's modified Eagle's medium (Mediatech, Herndon, VA) supplemented with 15% fetal bovine serum (Hyclone, Logan, UT), 2 mM L-glutamine (Mediatech), 0.1 mM β-mercaptoethanol (Fisher Chemical, Fairlawn, NJ), 1× nonessential amino acids (Mediatech), penicillin–streptomycin (Mediatech), and 103 U/mL leukemia inhibitory factor (Chemicon International, Temecula, CA). ESCs were re-fed with fresh media every other day, and ESCs were routinely passaged every 2–3 days before reaching 70% confluence.
Embryoid body formation
To form EBs, ESCs were detached from culture dishes using 0.05% trypsin-EDTA solution (Mediatech) before reaching 70% confluence and suspended in differentiation media (same as undifferentiated ESC media but without leukemia inhibitory factor). Hanging drop EBs were formed by suspending 600 cells/15 μL drop from the lid of square Petri dishes (10 × 10 cm) (Nunc, Rochester, NY). To prevent evaporation of the hanging drops, 10 mL of a sterile 1% solution of BSA in PBS was added to the bottom of each dish. Suspension EB cultures (static and rotary) were inoculated with 2–4 × 105 cells/mL in 10 mL differentiation media in 100 × 15 mm nontissue culture–treated polystyrene (i.e., bacteriological grade) Petri dishes (Becton Dickinson Biosciences, San Jose, CA). A thin layer of sterile 2% agar (Sigma) was used to precoat static plates to prevent cell attachment. Static and rotary suspension culture media was refreshed every other day, but hanging drop media was not changed during the 7-day suspension period. Rotary suspension EBs were placed on a rotary orbital shaker (Lab-Line Lab Rotator, Model #2314; Barnstead International, Dubuque, IA) set at ∼40 rotations per minute and calibrated daily for the entire suspension culture period to ensure consistent speed was maintained throughout the studies.
Spontaneous contractile activity assessment
After 7 days of suspension culture, EBs from each formation method were collected and individually plated onto 0.1% gelatin-coated 48-well tissue culture dishes to prevent the spreading of individual EB colonies into one another. The development of contractile foci within EB colonies was monitored every other day (8, 10, 12, and 14) to assess the frequency of cardiomyogenic differentiation. Suspension EBs were collected by diluting 200 μL of EBs into 5 mL fresh media within a new Petri dish, and hanging drop EBs were collected by harvesting one 100 × 100 dish (∼50 EBs) and placing into 5 mL media within a new Petri dish. Individual EBs were selected from the diluted suspension with 1 mL, wide bore pipet tips (100 μL volume; Rainin, Oakland, CA), dispersed into individual gel-coated wells of a 48-well plate, and an additional 400 μL of media was added for a final volume of 500 μL per well. After 24 h, wells were visually inspected to ensure each well contained at least one EB; in cases where 2 or 3 EBs occupied the same well (∼2% of wells; no wells contained >3 EBs), merged EBs were excluded from the contractile analysis. Blinded counts of EB colonies with one or more spontaneously beating foci were scored as “contractile,” and the percentage of contractile EBs relative to the total number of plated EBs was calculated (minimum of 30 EBs per experimental sample; >90 EBs total assessed per condition).
Quantitative reverse-transcriptase polymerase chain reaction
Total RNA was extracted from undifferentiated ESCs and EBs from each of the different formation methods at various time points of differentiation using the RNeasy Mini kit (Qiagen Incorporated, Valencia, CA). Reverse transcription for complementary DNA synthesis was performed using the iScript cDNA synthesis kit (Bio-Rad, Hercules, CA), and quantitative PCR was performed with SYBR green technology on the MyiQ cycler (Bio-Rad). Primers, sequences, and annealing temperatures are listed in Table 1 for Oct-4, Nanog, Brachyury T, Gata4, Nkx2.5, Myocyte enhancer factor-2c (Mef2c), Myosin light chain-2 ventricle (MLC-2v), α-myosin heavy chain (α-MHC), myogenic factor 5 (Myf5), and glyceraldhyde-3-phosphate-dehydrogenase (GAPDH). Each primer set was designed with Beacon Designer software (Invitrogen, Carlsbad, CA), and validated with appropriate cell controls. Relative gene expression levels of Oct-4 and Nanog (pluripotent ESC markers) were internally normalized to GAPDH (housekeeping gene) and quantified relative to undifferentiated ESC expression levels using the Pfaffl method of quantification. 36 For differentiation cell markers, absolute gene expression concentrations were calculated against standard curves and internally normalized versus levels of GAPDH expression.
Histology and immunohistochemistry
Suspension EBs were sampled from each group at day 7 of differentiation, and at day 10 (after EB plating), EBs were scraped from gelatin-coated dishes for immunohistochemical (IHC) analysis. The samples were fixed for 30 min in formalin (4% formaldehyde), embedded in Histogel® (Richard Allen Scientific, Kalamazoo, MI), and polymerized for 2 h at 4°C in cryomolds (10 × 10 × 5 mm; Sakura Finetek, Torrance, CA). After polymerization, the EBs entrapped within Histogel were dehydrated through a graded series of alcohol solutions (70–100%) and xylene, and embedded in paraffin. Five-micron sections were obtained using a rotary microtome (Microm HM 355S) with 80 μm intervals between sections to ensure that unique EBs were analyzed within the different sections, and histological slides were de-paraffinized before staining. For morphological assessment, slides were stained with hematoxylin and eosin using a Lieca AutoStainer XL.
For IHC analysis, de-paraffinized samples were incubated for 1 h in a 0.05% (weight/volume) Triton X solution of 2% horse serum in PBS at room temperature to permeabilize and block the sections. A primary anti-α-sarcomeric actin antibody (1:5000, 5c5; Sigma) was incubated overnight at 4°C, followed by three PBS rinses. A secondary biotinylated horse anti-mouse antibody (1:400; Vector Laboratories, Burlingame, CA) was incubated at room temperature for 30–60 min, followed by three rinses in PBS. The avidin–biotinylated alkaline phosphatase complex was incubated in solution for 30 min at room temperature before application, per vendor's instructions (Vectastain ABC-AP kit; Vector Laboratories), applied to histological sections for 30 min at room temperature, and rinsed twice with PBS and once in 100 mM Tris, pH 8.2, before developing with the chromagen, Vector Red SK-5100 (Vector Laboratories). Cell nuclei were counterstained with hematoxylin (Lieca AutoStainer XL), dehydrated with alcohol and xylene, mounted with Cytoseal™ 60 mounting medium (Richard Allen Scientific), cover-slipped, and imaged on a Nikon 80i upright microscope with a SpotFlex camera. Four to six independent fields were imaged per slide, and images were used to quantify the percentage of cardiomyogenic EBs. Sections of mouse heart and intestine were included with each batch of immunostaining as positive and negative controls, respectively.
Image analysis
For quantitative analysis, only distinct cell clusters >2500 μm2 were considered to be individual EBs. Smaller groups of cells (<2500 μm2) representing partial EBs or debris from scraping were excluded from the quantitative analysis. Blinded counts were performed of the total number of EBs and the number of EBs with positive regions of α-sarcomeric actin expression to report the percentage of cardiomyogenic EBs. IHC EB count data from independent experiments were combined (n = 4 for hanging drop and rotary EBs; n = 3 for static EBs), and a minimum of 40 EBs per condition were assessed. Within NIH ImageJ, a threshold for colorimetric staining was determined according to positive and negative control slide staining intensity and used to define the areas of positive α-sarcomeric actin staining within EBs. A macro program was written and employed within NIH ImageJ to measure the relative cross-sectional area of positively stained cells within individual EBs (as defined by the threshold), as well as the total EB cross-sectional area, and the relative area of α-sarcomeric actin immunoreactivity was calculated for individual EBs.
Flow cytometry
EBs plated at day 7 of differentiation were trypsinized (0.25% trypsin–EDTA) 3 days later (day 10 of differentiation) and triturating every 10 min for a total of 30 min to attain single-cell suspensions for flow cytometric analysis. Cell suspensions were centrifuged at 200 rcf for 5 min to collect the cells, resuspended in 1 mL PBS, transferred to 1.5 mL centrifuge tubes, centrifuged again at 3500 rcf for 8 min, and resuspended in 1 mL formalin for 5 min with rotation at room temperature. Cells were collected by centrifugation (8 min, 3500 rcf) and rinsed three times with PBS; after the final rinse and centrifugation, the cell pellet was resuspended in 1 mL of working buffer (0.3% BSA, 0.001% Tween-20 in 1 × PBS) supplemented with 0.05% Triton X-100 and 10% goat serum, and rotated for 30 min at room temperature. Cell suspensions were divided into equal quantities of 5 × 105 cells for further antibody incubations and analysis, and cells were rinsed and centrifuged three times with working buffer. The resulting cell pellets were resuspended in 300 μL of antibody solution in working buffer (1:500 dilution of 5c5 antibody or 1:500 dilution of isotype control IgM) and rotated for 30 min at room temperature, followed by successive rinse and centrifugation steps. The cell pellet was resuspended in 300 μL of secondary antibody solution in working buffer (1:10 dilution FITC-conjugated polyclonal goat-anti-mouse immunoglobulins; Dako North America, Carpinteria, CA) with rotation in the dark for 30 min at room temperature followed by a series of three rinses in working buffer and centrifugation. The final cell pellet was resuspended in 500 μL of PBS, filtered through the 35 μm cell-strainer cap of a BD Flacon™ 5 mL polystyrene round-bottom tube (#352253; BD Falcon, Franklin Lake, NJ), and run on a digital flow cytometer to capture a minimum of 10,000 events (BD dLSR; BD Biosciences). Analysis was performed using FlowJo software (Tree Star, Ashland, OR), and positive gates were set at 1% of the IgM isotype control population. The 5c5 anti-α-sarcomeric actin was validated for flow cytometry settings using the immortalized cardiomyocyte-like cell-line HL-1 (kindly donated by Dr. William Claycomb). 37
Statistical analysis
Experimental conditions were examined with triplicate samples in independent experiments, and data presented reflect the mean value ± standard deviation. Analysis of variance was performed to determine statistical significance (p < 0.05) between experimental groups, and, where significant, was followed by post hoc Tukey analysis to define statistical differences (p < 0.05) between specific experimental variables.
Results
Morphological comparison
The morphology of EBs formed by hanging drop (Fig. 1A, D, G), static (Fig. 1B, E, H), or rotary (Fig. 1C, F, I) suspension culture methods was compared after 7 days of suspension culture. Consistent with previous results, 35 individual rotary EBs were similar in shape and size to those formed by hanging drops (Fig. 1A, C). Rotary culture prevented agglomeration of EBs compared to static suspension, which exhibited a large variance in EB size and amorphous shapes resulting from EB agglomeration (Fig. 1B, E). Rotary suspension culture also yielded a larger number of EBs than static suspension with an equivalent number of initial cells. Histological analysis indicated that internal EB organization was also similar between rotary and hanging drop EBs (Fig. 1G, I), whereas static EBs contained increased regions of cell necrosis within the center of large agglomerates of EBs (Fig. 1H). Altogether, rotary orbital culture produced relatively large batches of homogenous EBs in suspension that closely resembled hanging drop EB morphology. Because hanging drop culture has been previously shown to enhance mesoderm and cardiac differentiation of ESCs,30,38 the effects of rotary orbital suspension culture on cardiomyogenic differentiation of EBs were further assessed.

EB morphology. At 7 days of differentiation, rotary EBs (
Development of contractile foci
The development of spontaneously contractile, rhythmically beating foci within EBs is a commonly observed indication of cardiomyogenic differentiation. The onset of spontaneous beating was detected within EBs from hanging drop, static, and rotary culture typically 1 day after plating onto gelatin-coated dishes (Fig. 2). Hanging drop culture consistently yielded the highest frequency of contractile EBs compared to either static or rotary culture methods, with > 60% of hanging drop EBs beating at 8 days of differentiation and reaching 89.4 ± 6.7% by day 10 of differentiation. EBs generated by rotary orbital suspension culture demonstrated a significantly greater occurrence of spontaneous beating activity compared to EBs cultured in static suspension at each time point observed (p < 0.01). At 8 days of differentiation, 37.4 ± 23.7% of rotary EBs exhibited contractile foci, while only 5.9 ± 4.1% of static EBs contained beating areas. By 10 days of differentiation, 50.7 ± 16.7% of rotary EBs contained contractile activity, whereas static culture only achieved 20.7 ± 11.2% contractile EBs (Fig. 1). About 50% of rotary EBs retained contractile activity through day 12 of differentiation, but a slight decline in the percentage of contractile EBs was observed by 14 days, similar to hanging drop EBs, which also exhibited a slight decline in spontaneous contraction after 12 days of differentiation. The significant differences in spontaneous beating activity of EBs observed between the different culture methods suggested that an increased potential of cardiomyogenesis was induced by rotary orbital culture compared to static suspension culture.
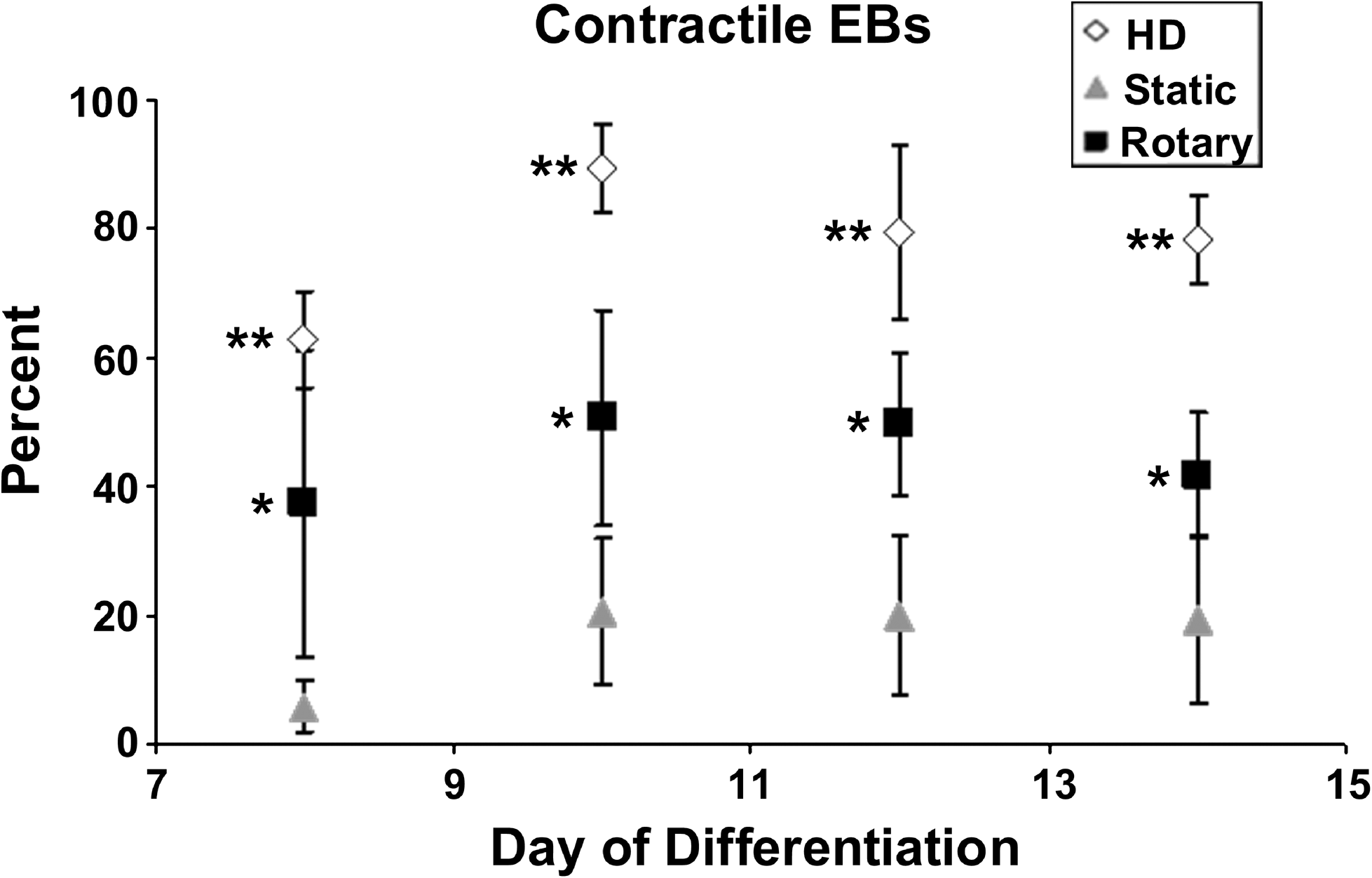
Percent contractile EBs. After day 8 of differentiation, rotary EBs consistently exhibited approximately 50% contractile EBs (days 10 and 12), while static EBs achieved only a maximum of about 20% (days 10, 12, and 14). Hanging drop (HD) EBs consistently exhibited beating foci in >80% of EBs, between days 10 and 14 of differentiation. By day 14, both hanging drop and rotary EBs slightly decreased contractile EB percentages. Data points represent the combined sum of data from three independent experiments. n = 9 (HD and rotary) or 6 (static); p < 0.01, *compared to static and **compared to static and rotary.
Gene expression
Based upon the differences observed in contractile activity of EBs elicited by the different formation methods, quantitative gene expression analysis of cardiac differentiation was performed. The expression of pluripotent genes, Oct-4 and Nanog, progressively decreased over the course of EB differentiation such that by 7 days of suspension culture, negligible levels remained detectable and no significant differences in pluripotent gene expression were observed between EBs from the different formation methods (Fig. 3A, B). Hanging drop and rotary EBs exhibited significantly decreased expression of Nanog by day 4 compared to static EBs (Fig. 3B), and hanging drop EBs expressed significantly less levels of Oct-4 at day 4 of differentiation compared to rotary and static culture methods (Fig. 3A). Brachyury T, a transiently expressed mesoderm transcription factor, was expressed by EBs formed by each of the different methods at day 4 of differentiation, but at significantly greater levels by rotary EBs compared to hanging drop and static EBs (Fig. 3C). The cardiac transcription factors Gata4, Nkx2.5, and Mef2c were expressed by rotary, static, and hanging drop EBs, although the onset and extent of expression differed. Hanging drop and rotary EBs exhibited significantly increased levels of Gata4 at day 4 or 7 (respectively) compared to either of the other culture methods (Fig. 3D). By day 7 of differentiation, rotary and hanging drop EBs exhibited a significant increase in expression of Nkx2.5 compared to static EBs (27 × and 44 ×, respectively) (Fig. 3E). Similar to patterns of Nkx2.5 expression, levels of Mef2c expression were significantly greater in hanging drop EBs at day 4 of differentiation, and expression of Mef2c by rotary EBs was increased by day 7 of differentiation, relative to either hanging drop or static suspension cultures (Fig. 3F). In addition to cardiac transcription factors, rotary and hanging drop EBs expressed transcripts for sarcomeric muscle proteins, α-MHC and MLC-2v, at comparable levels at day 7 of differentiation (∼9-fold and ∼6-fold greater than static EBs) (Fig. 3G, H). Myf5, an early skeletal muscle-specific transcription factor, was not expressed until 14 days of EB differentiation, indicating that expression of sarcomeric muscle markers before 14 days was specific for cardiac muscle, similar to previous reports. 39 Altogether, the expression profiles of genes directing EB cardiomyogenic differentiation indicated that rotary orbital suspension promoted a cardiomyogenic genotype similar to hanging drop culture more robustly than static suspension methods.
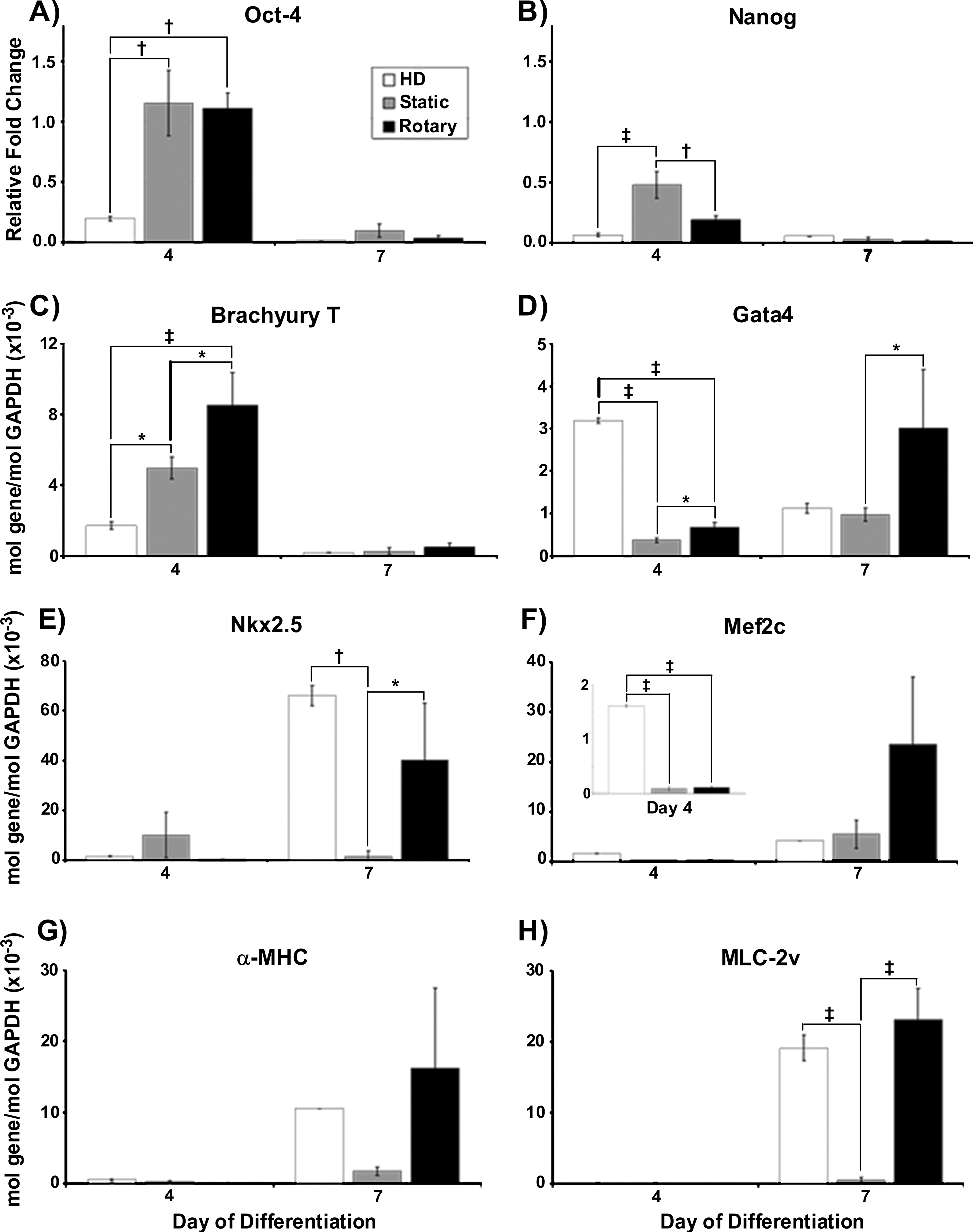
Gene expression. Hanging drop (HD) EBs exhibited a decrease in Oct-4 (
α-Sarcomeric actin immunostaining
IHC staining for α-sarcomeric actin was performed to identify and confirm the presence of developing cardiomyocytes within EBs. At early stages of EB differentiation (before 14 days), immunostaining for α-sarcomeric actin is a reliable phenotypic marker of cardiomyocyte differentiation by ESCs because of the coincident absence of early skeletal muscle transcription factors, as previously noted. 39 By day 10 of differentiation, clusters of α-sarcomeric actin + cells were readily observed in cross sections of many rotary and hanging drop EBs, but few static EBs stained positive for α-sarcomeric actin (Fig. 4, top row). In addition, α-sarcomeric actin + regions appeared to occupy a larger portion of the cross-sectional area of rotary EBs, occasionally comprising a majority of the EB interior (Fig. 4, middle row). The morphology of α-sarcomeric actin + cells within rotary and hanging drop EBs was consistent with that of immature cardiomyocytes before the development of distinct, organized sarcomeres or myofibrils (Fig. 4, bottom row). Quantitative assessment of cardiomyogenic (α-sarcomeric actin+) EBs confirmed that a significantly increased percentage was attained with rotary and hanging drop methods (78 ± 18% and 66 ± 25%, respectively) compared to static suspension culture (7 ± 8%, Fig. 5A) at day 10 of differentiation. Further, rotary EBs contained 36.9 ± 27.8% of sarcomeric actin + area, whereas hanging drop and static EBs contained 13.5 ± 2.1% and 18.8 ± 4.7%, respectively (Fig. 5B). Thus, IHC staining analysis, consistent with spontaneous contractile activity and cardiac gene expression, suggested that rotary orbital culture promoted the differentiation of cardiomyocytes to a greater extent than static suspension culture conditions.
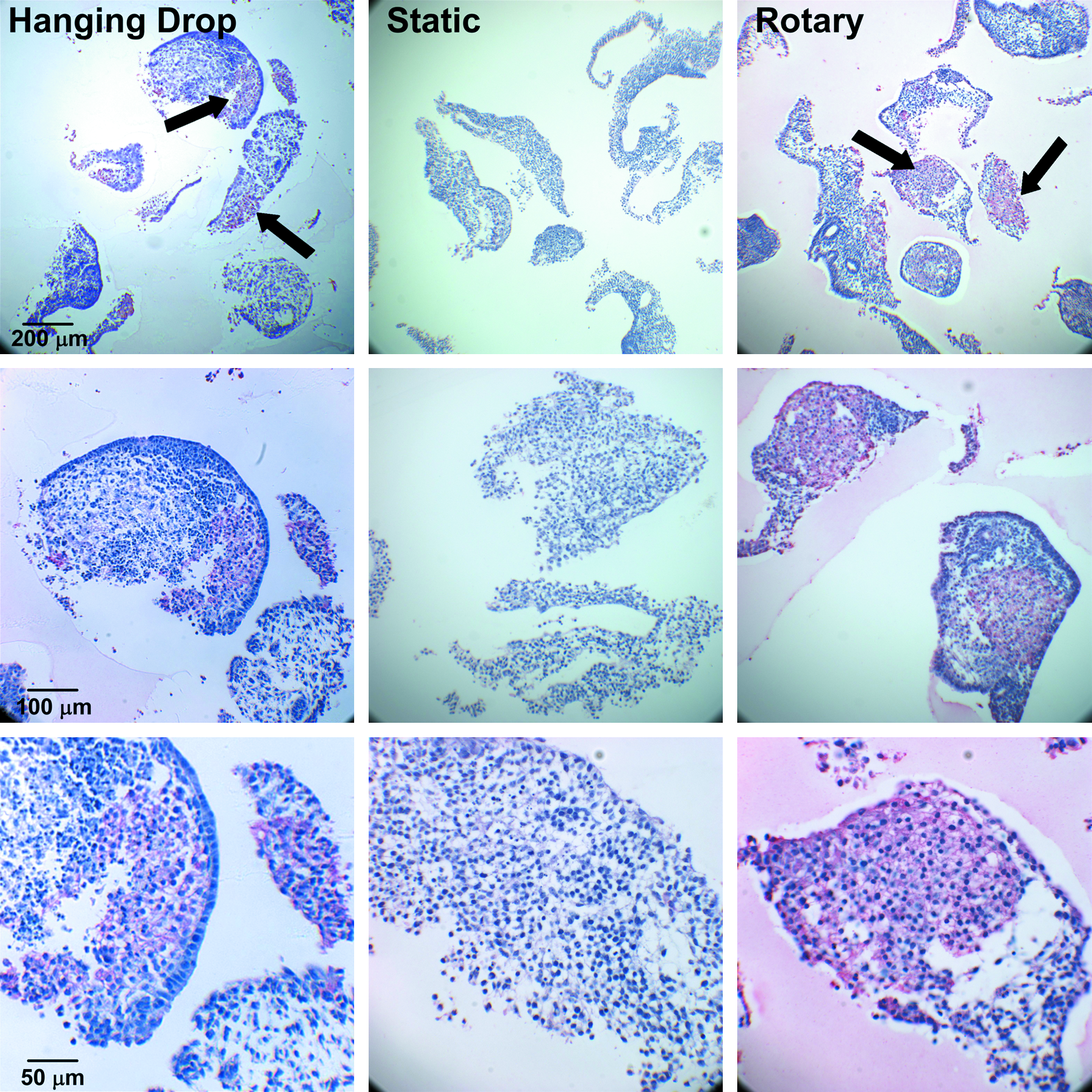
Sarcomeric protein staining. α-Sarcomeric actin IHC staining using 5c5 antibody within hanging drop (left), static (center), and 40 rpm rotary (right) EBs. Pink/red staining indicates 5c5 + clusters of cells within hanging drop and rotary EBs, but no apparent positive areas in static EBs. Top row, low magnification; middle and bottom rows, high magnification. Black arrows indicate positive stain. Color images available online at www.liebertonline.com/ten.
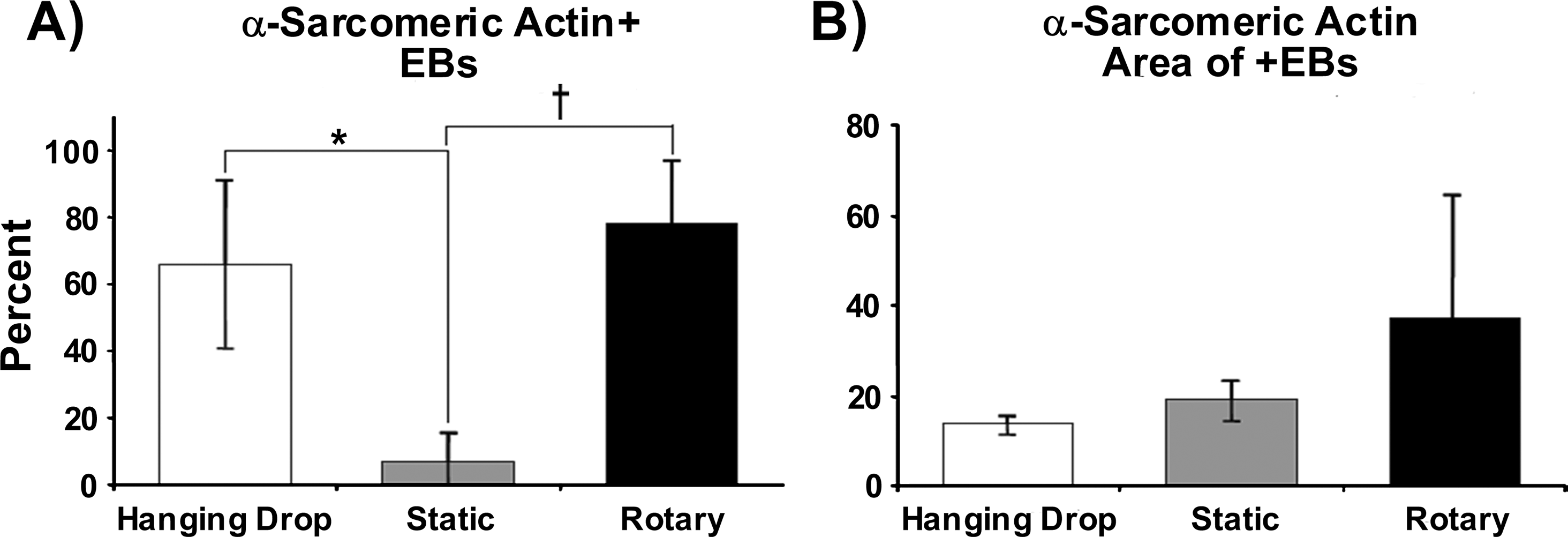
Quantification of sarcomeric protein staining. Percent of EBs stained positive for α-sarcomeric actin at day 10 of differentiation from hanging drop, static, and rotary culture conditions (
α-Sarcomeric actin immunocytometry
Flow cytometry analysis for cardiac myocyte immunoreactivity of dissociated EBs was performed to assess the percent of α-sarcomeric actin + cells yielded by the different EB culture methods. Based on histogram analysis, the population of cells from rotary and hanging drop EBs exhibited a distinct positive shift in fluorescent signal intensity compared to static EBs and immunostaining controls (Fig. 6A). Static EBs contained 3.54 ± 3.07% α-sarcomeric actin + cells, whereas hanging drop EBs contained 6.71 ± 3.63% and rotary EBs possessed 13.08 ± 7.37% α-sarcomeric actin + cells (Fig. 6B). The average value of α-sarcomeric actin + cells was calculated by quantifying the positive number of gated fluorescent events relative to the overall population of cells (Fig. 6C). Thus, consistent with the results of gene expression and IHC staining, flow cytometry studies suggested that rotary orbital culture enriched the number of cardiomyocytes within EBs compared to the other culture techniques examined.
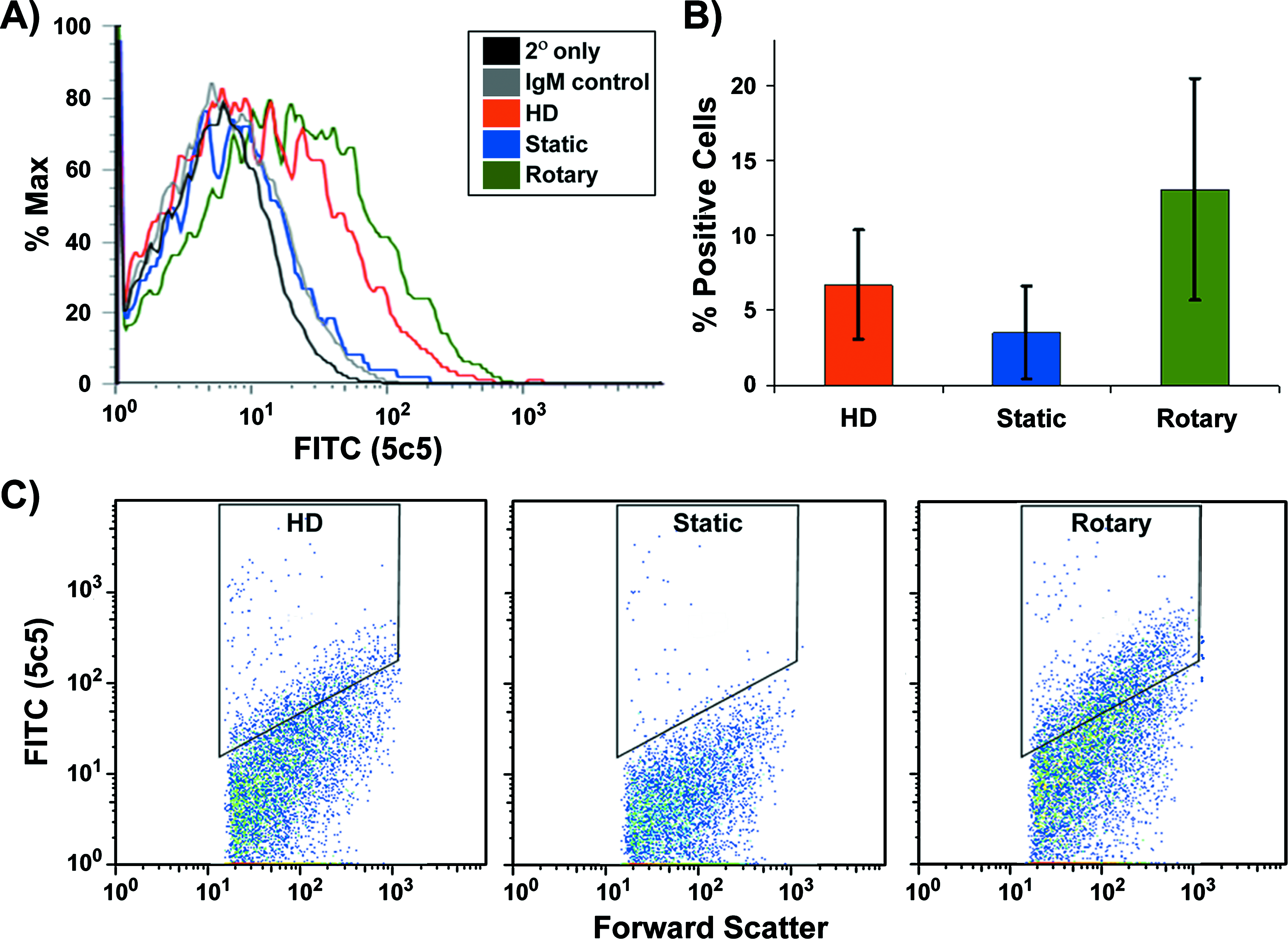
Sarcomeric protein immunocytometry. Histogram representation of associated α-sarcomeric actin expression within dissociated EBs from hanging drop, static, and rotary orbital culture at day 10 of differentiation (
Discussion
In this study, the effects of rotary orbital suspension culture on cardiomyocyte differentiation within EBs were assessed compared to hanging drop and static suspension culture methods. Suspension culture of EBs under rotary orbital conditions for 7 days resulted in a significantly greater frequency of spontaneous contractile EBs than static culture. Differentiation proceeded within EBs from each formation method, but rotary EBs exhibited significant increases in the expression of genes associated with mesoderm and cardiac differentiation, similar to hanging drops. Rotary orbital culture also increased the proportion of α-sarcomeric actin + EBs and augmented the relative size of sarcomeric actin + regions within individual EBs, as well as the percent of α-sarcomeric actin + cells within rotary EBs as determined by immunocytometric analysis. These results indicate that rotary orbital suspension culture promotes enhanced cardiomyogenesis of EBs relative to static suspension, and thus may increase not only the efficiency but also the yield of cardiomyocytes differentiated from EBs because rotary orbital culture increases the overall numbers of EBs and differentiated cells compared to static suspension culture. 35
The endogenous yield of cardiomyocytes from spontaneously differentiating ESCs is typically only 1–5% of the total differentiated cell population.23,40 It is widely accepted that hanging drop culture conventionally increases cardiomyogenic differentiation compared to static suspension,30,31,38 suggesting that the method of EB formation can impact the proportion of cell phenotypes within EBs. Despite these observations, currently there is a lack of direct quantitative comparisons of cardiomyogenesis between different formation techniques. The 3.5% of α-sarcomeric actin + cells within static EBs at day 10 of differentiation reported in this study (Fig. 6) is similar to previous reports of 3.9% sarcomeric myosin + cells (Mab MF20) at day 9 of differentiation 40 and 2% at day 14 of differentiation 41 from statically cultured EBs using the mouse ESC D3 cell line. In this study, cardiomyogenic differentiation was enhanced almost fourfold by hydrodynamic culture conditions created by rotary orbital mixing compared to static suspension in the same culture vessel. The differences in efficiency of cardiomyocyte differentiation between EB formation methods implicate the ability of microenvironmental conditions to dictate endogenous levels of cell differentiation.
Several elements of the microenvironment can simultaneously impact stem cell differentiation, including soluble cytokines and nutrients, cell–matrix and cell–cell adhesions, and mechanical forces.42,43 Coculture of EBs with endoderm explants and visceral endoderm–like cells or treatment with conditioned media from endoderm cells increases cardiomyogenic differentiation, presumably due to secreted factors.44–46 Similarly, other directed differentiation techniques rely on exogenous addition of specific soluble factors capable of enhancing cardiomyogenesis of EBs, such as bone morphogenic protein 2 (BMP-2),44,47 transforming growth factor beta-1 (TGFβ-1), 47 insulin-like growth factor 1 (IGF-1), 48 and retinoic acid,40,49 several of which are secreted from developing endoderm. The fact that rotary culture alone promotes cardiomyocyte differentiation of EBs suggests that endogenous production of cardiomyogenic molecules within EBs may be enhanced. Previous studies indicated that rotary orbital culture increased endoderm differentiation 35 ; thus, rotary orbital suspension culture may promote endoderm-derived cardiomyogenic factors locally within EBs. Rotary suspension may create a pro-cardiomyogenic microenvironment within EBs that could potentially be further enhanced by additional supplementation of cardiac-inductive molecules to the culture media.
Genetic modification and physical separation techniques can be used to increase the purity of cardiomyocytes attained from differentiating EBs. For example, the resulting purity of mouse ESC–derived cardiomyocytes can be increased to 70% or greater (from <5% originally) by either positive or negative selection strategies used in conjunction with genetically engineered cell lines.21,40,50 Physical separation of cells by density centrifugation, such as Percoll gradients, has been used to increase the purity of cardiomyocytes derived from both mouse and human ESCs.26,50,51 However, an increase in cell purity does not actually reflect an increase in cell yield derived from ESCs, but rather, simply an enrichment of a particular cell phenotype (i.e., cardiomyocytes) relative to the overall differentiated cell population. As noted previously, the rotary orbital culture method alone can increase the total yield of differentiating cells from EBs compared to static suspension culture inoculated at equivalent starting densities. 35 Based on the present study, rotary culture also endogenously enhances cardiomyocyte differentiation, and selection techniques could be easily integrated downstream of rotary orbital suspension culture to further increase the purity, as well as the yield of cardiomyocytes obtained from this method.
One novel environmental parameter introduced by rotary suspension culture is the effect of hydrodynamic mixing conditions on ESC differentiation. Although it was previously recognized that rotary orbital shaking promotes initial ESC aggregation and accelerates the kinetics of EB formation compared to static culture,35,52 the present study indicates that cardiomyocyte development from EBs is also enhanced by rotary culture. Although differences in EB size, agglomeration, and necrotic core formation have been observed within EBs from hanging drop, static, and rotary orbital suspension culture, 35 no apparent links between these morphological features and cardiomyocyte differentiation were clearly observed. Hydrodynamic mixing conditions imposed on EBs by rotary culture also introduce fluid shear stresses that may directly impact ESC and, more specifically, cardiovascular differentiation. In contrast to the wealth of studies examining the response of somatic (differentiated) cell types to fluid mechanical forces, very little is known about the characteristics or effects of fluid mechanics that may impact cardiogenesis during early mammalian development. Interestingly, intracardiac flow and the resultant shear forces are required during embryonic development to achieve normal cardiogenesis in zebrafish. 53 Recent work has also demonstrated that differentiating, adherent ESCs exposed to laminar shear stress via a cone-plate viscometer exhibited significant histone modification and subsequent increases in Mef2c and α-sarcomeric actin expression. 54 Other stirred suspension systems, including slow turning lateral vessel and spinner flask bioreactors, have been used to form EBs and differentiate cardiomyocytes; however, only 5.5% of cells were reported to be sarcomeric myosin + within spinner flask conditions before genetic selection, 40 and relatively small increases in human cardiomyocyte yield (1.5–2-fold >static suspension) were achieved after the implementation of selection methods.55–58 Although the mechanism remains unclear, hydrodynamic forces created by rotary orbital culture provide a simple, batch-culture system capable of promoting the endogenous cardiomyogenic differentiation of ESCs. Thus, the findings of the present study demonstrate a novel approach for engineering stem cell fate using hydrodynamic conditioning and provide design criteria to implement in the creation of novel bioreactors used to scale-up the production of differentiated cardiomyocytes from stem cells.
Footnotes
Acknowledgments
The authors would like to thank Gayathri Balasubramanian, Virginia Chu, and Beth Krauth at the Georgia Institute of Technology for technical assistance. This work was supported by funding provided by the American Heart Association (0665265B) and the Georgia Tech/Emory Center for the Engineering of Living Tissues (GTEC, NSF EEC-9731463). C.Y.S. is supported by a National Science Foundation Graduate Fellowship, and G.Y.B. was supported by funding from GTEC for an Undergraduate Research Scholars award.
Disclosure Statement
No competing financial interests exist.