
Abstract
Similar to native cardiovascular tissues, the mechanical properties of engineered cardiovascular constructs depend on the composition and quality of the extracellular matrix, which is a net result of matrix remodeling processes within the tissue. To improve tissue remodeling, and hence tissue mechanical properties, various mechanical conditioning protocols, such as strain-based or flow-based conditioning, have been applied to engineered cardiovascular constructs with promising results. We hypothesize that tissue remodeling is dependent on the mode of straining. Therefore, the effects of two modes of straining, being either static or dynamic, were quantified on several indices of tissue remodeling. Differences in matrix composition (collagen and glycosaminoglycans [GAGs]) and maturity (collagen cross-links) were quantified with time on gene expression and protein levels. In addition, the secretion of specific collagen remodeling markers (matrix metalloproteinase-1), collagen synthesis marker (procollagen type I carboxy-terminal propeptide, PIP), and collagen degradation marker (carboxyterminal telopeptide of type I, ICTP) was investigated with time. Static strain stimulated collagen gene expression and production with time. Dynamic straining resulted in (1) lower collagen gene expression and production, but (2) enhanced collagen cross-link expression and density, and GAG production, and (3) stimulated collagen remodeling processes, as expressed by enhanced production of remodeling markers. Thus, despite a lower collagen production, the quality of the neotissue was enhanced by a dynamic straining component. These straining mode–dependent remodeling responses allow us for the first time to balance collagen and cross-link production and, thus, to fine tune tissue mechanical properties via mechanical conditioning protocols. This is of utmost importance for cardiovascular tissue engineering, where insufficient mechanical properties are currently a main limiting factor for present in vivo application.
Introduction
Mimicking native mechanical behavior represents a major goal in cardiovascular tissue engineering, in particular, when facing high-pressure applications. Similar to native cardiovascular tissues, the mechanical properties of tissue-engineered constructs depend on the composition and quality of the extracellular matrix, which is a net result of matrix remodeling processes in the tissue. The extracellular matrix of tissue-engineered constructs predominantly consists of collagen fibers, embedded in a gel of proteoglycans, including glycosaminoglycans (GAGs). Collagen is the main load-bearing part of these tissues, and collagen cross-links and remodeling define their maturity and quality.
So far, tissue engineering of cardiovascular substitutes based on fast-degrading scaffolds seeded with autologous myofibroblasts has been demonstrated as a promising procedure to produce living heart valve and blood vessel replacements.6,7 In this approach, cells are stimulated to produce de novo extracellular matrix components in vitro. However, engineered tissues often lack sufficient amounts of properly organized matrix components and consequently do not meet in vivo mechanical demands. To optimize tissue quality and hence improve the mechanical properties of engineered tissues, mechanical conditioning strategies are critical.8–10
Various conditioning protocols, such as flow-based and strain-based protocols, have been demonstrated to improve tissue properties. Previous conditioning protocols often intended to mimic physiological circumstances. Using an in vitro pulse duplicator, in which the tissue is exposed to increasing pulsatile flows, tissue-engineered heart valves have been obtained, which were successfully implanted at the pulmonary position in sheep. 7 Novel strain-based conditioning protocols have shown to result in functional tissues that hold the promise to serve as aortic valve 11 and small artery 12 replacements. However, the mechanical properties of these tissues still do not meet those of native cardiovascular structures due to insufficient remodeling. These strain-based conditioning protocols make use of static strain, induced by constraining of the tissue, and additional dynamic straining. The biological remodeling mechanisms underlying the strain-induced phenomena, however, remain unclear. Therefore, the goal of this study is to quantify strain-induced tissue remodeling. It is hypothesized that tissue remodeling depends on the mode of straining. Accordingly, the effects of two modes of straining, being either static or dynamic, were quantified on several indices of tissue remodeling over a period of 10 days. Differences in matrix composition (collagen and GAGs) and maturity (collagen cross-links) were quantified at the gene expression level and at protein levels with time. In addition, the secretion of specific collagen remodeling markers was investigated with time: matrix metalloproteinase-1 (MMP-1), collagen synthesis marker (procollagen type I carboxy-terminal propeptide, PIP), and collagen degradation marker (carboxyterminal telopeptide of type I, ICTP).
The present study is the first to explore such temporal effects in a well-defined model system of three-dimensional cardiovascular tissues. The effect of strain on matrix production has previously been studied in two-dimensional culture systems,13–17 lacking a physiological three-dimensional environment. Strain-stimulated remodeling has only been sparsely studied in three-dimensional cell-seeded constructs. In cell-seeded collagen gels, it has been demonstrated that dynamic strain influenced the gene expression of collagen and elastin. 10 However, corresponding levels of protein content were not determined. Studies investigating the effect of dynamic strain on the formation of neotissue in cell-seeded fast-degrading scaffolds do report matrix protein values, but only at the time of sacrifice after several weeks.18,19 Elucidating strain-induced tissue remodeling on gene and protein levels simultaneously is important to understand and predict strain-induced remodeling processes. This will provide valuable input for tissue engineering protocols to fine tune tissue properties using mechanical conditioning protocols.
Materials and Methods
Cell culture
Myofibroblasts were acquired from the human vena saphena magna according to the Dutch guidelines for secondary use material and expanded using regular cell culture methods as described previously. 20 Culture medium consisted of advanced Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA), supplemented with 10% fetal bovine serum (FBS; Greiner Bio One, Monroe, NC), 1% GlutaMax (Gibco, Carlsbad, CA), and 0.1% gentamycin (Biochrom, Terre Haute, IN).
Engineered cardiovascular tissues
Scaffold preparation and cell seeding procedures were performed as described previously. 18 Briefly, rectangular strips (35 × 5 × 1 mm) of nonwoven polyglycolic acid (PGA) meshes (Cellon, Bereldange, Luxembourg) were coated with a thin layer of poly-4-hydroxybutyrate 7 (TEPHA, Cambridge, MA). The PGA scaffold consists of fibers with a diameter in the order of 10–15 μm and pore sizes that are sufficiently large for complete cell penetration within 3 days. 21 The bottom surface of the scaffolds was reinforced with a nontoxic elastic silicone layer (Silastic MDX4-4210; Dow Corning, Midland, MI; thickness 0.5 mm), enabling precisely controlled cyclic deformation of the constructs up to several weeks. 19 In the longitudinal direction, the scaffolds were attached to the flexible membranes of six-well plates (Flexcell Int., McKeesport, PA) at the outer 5 mm using Silastic MDX4-4210. The scaffolds were vacuum dried for 48 h, followed by exposure to ultraviolet light for 1 h and were subsequently placed in 70% ethanol for 5 h to obtain sterility. Prior to cell seeding, tissue culture medium was added to facilitate cell attachment. The scaffolds were seeded with human venous myofibroblasts (passage 7) at a seeding density of 2 × 106 cells per cm2 using fibrin gel. During the seeding procedure, cells were centrifuged and resuspended in a thrombin solution (10 IU/mL) (Sigma Chemicals, St. Louis, MO), mixed with a fibrinogen solution (10 mg/mL) (Sigma Chemicals), and dripped evenly on the scaffold. The cell–thrombin–fibrinogen solution is absorbed throughout the whole scaffold. Subsequently, polymerization of the fibrin gel starts, serving as a cell carrier during culture. 22 The engineered constructs were cultured in tissue culture medium consisting of advanced DMEM (Gibco), supplemented with 10% FBS, 1% GlutaMax, 0.3% gentamycin, and L-ascorbic acid 2-phosphate (0.25 mg/mL; Sigma Chemicals). Medium was changed every 3 days and stored for medium analysis.
Tissue culture and mechanical conditioning
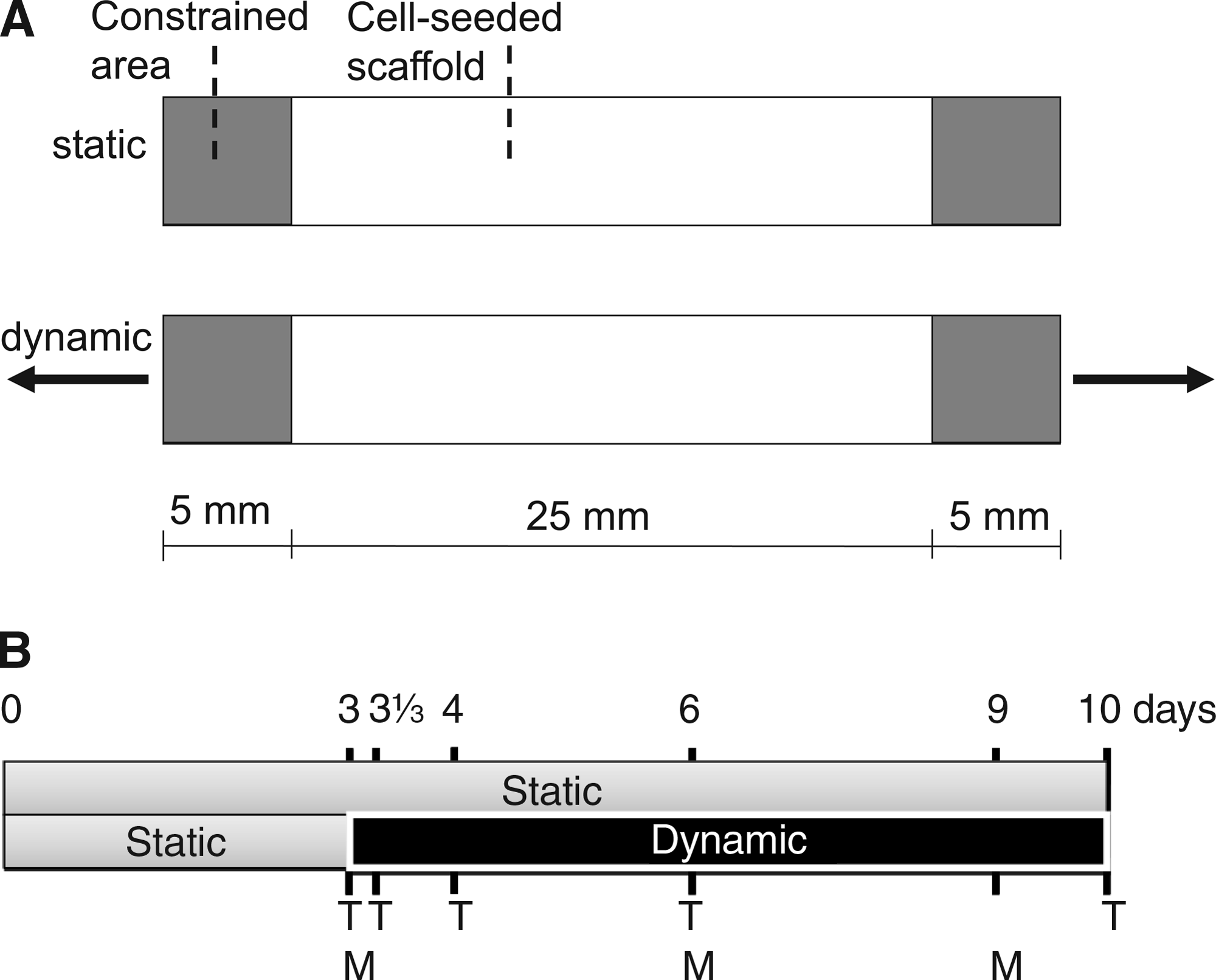
As the tissue-engineered strips were attached at the outer ends, static strain was generated by the cells as a response to constraining (Fig. 1A). After 3 days of culture under static conditions, four samples were sacrificed and used as baseline controls. Then, the engineered tissues were split into two groups. One group (n = 16) served as static control, whereas the other group (n = 16) was subjected to additional uniaxial dynamic straining (4%, 1 Hz) on top of static strain using a modified version of a Flexercell FX-4000T straining device 19 (Flexcell). A straining magnitude of 4% was chosen as a suitable straining condition based on previous studies where it was shown that continuous dynamic straining at 8% and higher deteriorated the mechanical properties compared to static strain. 19 Static and dynamically strained samples (n = 4 for each group per time point) were sacrificed after 3, 31\3 (8 h after start of dynamic straining), 4, 6, and 10 days of culture (Fig. 1B). The silicon layer was gently removed, and the samples were divided into two parts. Half of the sample was used for quantitative PCR (qPCR) analyses and the other half for tissue content analyses.
(
Quantitative PCR analysis
RNA isolation and semiquantitative mRNA analysis using real-time competitive PCR (ABI Prism 7700; Applied Biosystems, Nieuwerkerk aan den IJssel, The Netherlands) were performed as described previously.23,24 In brief, RNA was isolated using an RNAeasy extraction kit (Qiagen, Venlo, The Netherlands) according to the manufacturer's instructions. The concentration and purity of the RNA were determined by measuring the absorptions at 260 and 280 nm. Subsequently, 500 ng RNA was transcribed into cDNA using random primers. Gene expression of collagen I (COL1A2), collagen III (COL3A1), PLOD-2 (encoding for cross-link enzyme), and MMP-1 was analyzed by qPCR using specific forward and backward primers and FAM/TAMRA-labeled probes. All data were normalized to glyceraldehyde-3-phosphate dehydrogenase expression (GAPDH).
Quantification of matrix composition
Lyophilized samples were digested in papain solution (100 mM phosphate buffer, 5 mM L-cystein, 5 mM ethylenediaminetetraacetic acid, and 125–140 μg papain per mL) at 60°C for 16 h. The amount of DNA was determined using the Hoechst dye method, 25 and expressed per mg dry weight (dw). Subsequently, digested tissue samples were hydrolyzed in 6 M hydrochloric acid (Merck, Darmstadt, Germany) and used for amino acid analyses. Hydroxyproline (Hyp) residues, as a measure for collagen content, were measured on the acid hydrolysates using reverse-phase high-performance liquid chromatography after derivatization with 9-fluorenylmethyl chloroformate. 26 The amount of Hyp was expressed per DNA. The GAG content was determined using a modification of the assay described by Farndale et al. 27 The amount of GAG in the samples was determined from a standard curve and expressed per DNA.
The number of mature collagen hydroxylysyl pyridinoline (HP) cross-links, as a measure for tissue maturity, was determined in hydrolyzed samples using high-performance liquid chromatography.28,29 The number of HP cross-links was expressed per collagen triple helix (TH).
Medium analysis
Concentrations of the remodeling enzyme MMP-1 and markers for collagen synthesis (PIP) and collagen degradation (ICTP) were determined in culture medium by ELISAs. MMP-1 concentrations were quantified in culture medium by a nonisotopic immunoassay for human MMP-1 protein (Calbiochem, Merck, Darmstadt, Germany). Collagen synthesis marker PIP was measured using a procollagen type I C-Peptide ELISA kit (Takara Bio, Otsu, Shiga, Japan). Collagen degradation marker ICTP was determined using a quantitative enzyme immunoassay designed for in vitro measurement of carboxyterminal cross-linked telopeptide of human type I collagen (Orion Diagnostica, Espoo, Finland). The ELISAs were performed according to the recommendations from the supplier. The assays were performed on medium samples after 3, 6, and 9 days of culturing (Fig. 1b) and corrected for control medium that had been in contact with scaffolds without cells for 3, 6, and 9 days.
Statistics
Results are expressed as mean value ± standard deviation. Student's t-tests were used to elucidate differences between static and dynamic conditioning at each time point. Dunnett's multiple comparison tests were used to compare values of later time points to the 3-day static baseline control, except for HP cross-links, where values were not detectable at this time point. In this case, values of later time points were compared to the first detectable time point after 4 days of culture. Differences were considered to be significant at a p-value < 0.05.
Results
Effect of straining mode on gene expression
Human venous myofibroblasts expressed both collagen I and collagen III genes, of which collagen I mRNA was most abundantly expressed. Following 3 days of culture, dynamic strain was applied to the samples in the dynamic group. Both collagen I and III mRNA levels were significantly lower in dynamically strained samples from day 4 onward, as compared to static samples (Fig. 2A, B). In contrast, PLOD-2 mRNA expression was upregulated by dynamic straining, as compared to static conditioning (Fig. 2C). Compared to static strain, a significant higher MMP-1 mRNA expression was observed in the dynamically strained group after 6 and 10 days of culture (Fig. 2D). With time, static strain increased collagen I and III mRNA expressions, while no temporal variations compared to the 3-day baseline control were found as a consequence of dynamic straining. Dynamic straining increased PLOD-2 and MMP-1 mRNA expressions with time, as compared to the static baseline control after 3 days of culture. Static strain induced no temporal effects in PLOD-2 and MMP-1 mRNA expressions.

Collagen I (
Effect of straining mode on tissue protein content
The effects of static and dynamic conditioning on the amount of DNA, matrix proteins Hyp and GAG per DNA, as well as of HP cross-links per TH are depicted in Figure 3. There were no significant changes in the amount of DNA per dry weight between static and dynamic straining. In addition, no temporal variations were found as compared to the 3-day baseline control (Fig. 3A). The amount of Hyp per DNA increased with time in both groups, although dynamic straining was less favorable for collagen production, as compared to static strain (Fig. 3B). On the other hand, GAG production was stimulated by dynamic straining and increased with time (Fig. 3C). In contrast, static strain did not show an increasing trend in GAG production with time. HP cross-link densities were not detectable until 4 days of culture. Dynamic straining resulted in significant higher numbers of HP cross-links per TH, as compared to static strain (Fig. 3D). With time, cross-link densities increased in both groups.

Tissue content including DNA (
Effect of straining mode on collagen remodeling markers
The effects of different straining modes on specific collagen remodeling markers for collagen synthesis (PIP) and degradation (ICTP), and remodeling enzyme MMP-1 are shown in Figure 4. All of these remodeling markers showed similar effects. PIP, ICTP, and MMP-1 concentrations in culture medium of dynamically strained samples increased with time and were higher compared to static samples. No temporal variations were observed in the concentrations of these markers in the medium of statically conditioned samples.

Concentrations of PIP (
Discussion
Similar to native cardiovascular tissues, the mechanical properties of engineered cardiovascular constructs depend on the composition and quality of the extracellular matrix, which is a direct result of tissue remodeling. Tissue remodeling, in turn, is strongly influenced by mechanical loads applied to the tissue. During tissue engineering strategies, this is mimicked by conditioning the tissue with mechanical cues for prolonged periods of time. Current mechanical conditioning protocols for cardiovascular tissue engineering are mainly strain based, and have shown to result in functional tissues that hold the promise to serve as aortic valve and small artery replacements.6,11,12 Nevertheless, the mechanical properties of these engineered tissues still do not meet those of adult native cardiovascular structures as a consequence of insufficient remodeling during the in vitro culture periods. Elucidating strain-induced tissue remodeling is thus required to optimize tissue mechanical properties through mechanical conditioning.
While the majority of studies use two-dimensional systems, in this study a well-defined three-dimensional model system was used to explore temporal strain-induced remodeling responses in engineered cardiovascular tissues. It was hypothesized that these responses are influenced by the mode of straining. Therefore, the effects of two modes of straining, being either static or dynamic, were quantified on several indices of tissue remodeling with time. Differences in matrix composition (collagen and GAGs) and maturity (collagen cross-links) were quantified on gene expression and protein levels. In addition, the secretion of specific collagen remodeling markers (MMP-1, PIP, and ICTP) was investigated.
As native heart valves and blood vessels mainly consist of collagen type I and, less abundantly, of collagen type III, the capacity to produce these types of collagen is crucial for cardiovascular tissue engineering. Both collagen I and collagen III mRNA were expressed in the engineered tissues, of which collagen I mRNA was mostly expressed. Static strain increased collagen I and III mRNA expression with time. Apparently, the internal strains generated due to constraining the tissue are sufficient to stimulate cells to express collagen mRNA. This emphasizes the relevance of constraining during tissue culture. Additional dynamic straining resulted in lower collagen I and III expressions. Similar effects were found for the Hyp protein levels. Apparently, dynamic straining might not be required for the production of collagen over the applied culture time. Obviously, cells need some time to produce, remodel, and mature the tissue. Although collagen III expression in the dynamically strained group at day 10 was not significantly different from the 3-day baseline control (p = 0.15), there seems to be an increase in collagen III expression from 6 days onward. Similar to the wound healing process, 30 during in vitro tissue engineering, collagen type III might be earlier formed and deposited than collagen type I. These early present collagen fibers would then be remodeled, leading to a mature tissue, consisting of mainly type I collagen, which is the predominant type in cardiovascular tissues.
Besides collagen content, also collagen cross-links play an important role in tissue maturity and determination of the mechanical properties. 31 The mature HP cross-links are the main type of collagen cross-links present in cardiovascular tissues. These cross-links are formed following hydroxylation of the telopeptides of collagen. PLOD-2 has been identified as a telopeptide lysysl hydroxylase, the enzyme responsible for this hydroxylation process. 24 In the present study, PLOD-2 mRNA expression levels of dynamically strained constructs increased with time and were higher than for statically conditioned constructs. Thus dynamic straining seems crucial to upregulate PLOD-2 to enhance the formation of HP cross-links.
The effects of straining on gene expression were consistent with those at the level of protein expression: dynamic straining resulted in lower collagen amounts, but enhanced cross-link densities, as compared to static strain. This suggests that during continuous dynamic straining, cells switch their production balance from collagen to cross-link synthesis, thus catalyzing a more stable deposition of collagen fibers. Despite a lower collagen amount, the quality and structural stability of the neotissue is enhanced by cross-links, enabling properly organized collagen fibers. In contrast to the effect on collagen production, the production of GAGs was enhanced by dynamic straining. Most likely, GAG production is upregulated to embed, stabilize, and protect the formed matrix components.32–34
Besides quantification of tissue remodeling on gene expressions and protein levels, specific collagen remodeling markers were investigated. Markers for collagen synthesis (PIP) and degradation (ICTP) were measured to provide insight in the straining mode dependency of the collagen turnover balance. Despite a lower collagen mRNA expression compared to static conditioning, dynamic straining enhanced PIP. The formation of collagen from soluble collagen occurs by releasing the procollagen propeptides (PIP). This process seems to be enhanced by dynamic straining. Collagen degradation marker ICTP, which is released after cleavage of collagen by MMPs, was also enhanced by dynamic straining. The balance between synthesis and degradation, and the resulting net collagen content, appeared to be straining mode dependent. Dynamic straining increased both synthesis and degradation, and hence remodeling, but resulted in lower net collagen levels as compared to static strain. In addition, higher MMP-1 mRNA expression and protein levels were found in response to dynamic loading. Changes in remodeling enzymes, such as MMPs, have been reported to play a key role in tissue maintenance, repair, and strain-induced remodeling.35,36 The relevance of MMPs in cardiovascular development and remodeling has been confirmed by highly elevated levels of MMP-1 in fetal developing heart valves compared to very low values in adult valves. 37 These results suggest a direct correlation between MMP-1 levels and remodeling activity. So, in addition to enhanced collagen synthesis and degradation markers, enhanced MMP-1 levels in dynamically strained samples also indicate enhanced remodeling activities due to dynamic straining.
It is likely that the strain-enhanced cross-link densities and remodeling activities will eventually lead to a more mature collagen architecture and improved mechanical properties. Other studies based on similar cell-seeded biodegradable scaffolds demonstrated improved mechanical properties after 4 weeks of culture due to dynamic straining while collagen levels were equal to statically strained samples.11,19 The present study now indicates that the improved mechanical properties are merely due to higher cross-link densities than to collagen levels.
To reach native values in engineered cardiovascular constructs, mechanical conditioning protocols need to be optimized and controlled. Where regular autologous tissue engineering protocols take several weeks, we have shown that already after several days, straining mode–dependent differences are induced and detectable. Interestingly, effects on the level of gene expression corresponded to protein data. This means that accurate and fast qPCR studies in this well-defined model system can be used as valuable tools to unravel the effects of other straining parameters, such as straining period, magnitude, and frequency. Moreover, the analysis of medium samples enables an online nondestructive measurement of collagen remodeling. The results of this study are helpful toward a well-considered optimization of tissue engineering protocols. To combine the favorable effects of collagen production due to static strain and of cross-link production due to dynamic straining, current studies in our lab focus on intermittent straining protocols. In addition, a sequential conditioning protocol, in which sufficient amounts of collagen will be produced before cross-links and remodeling activities are induced by a dynamic component, may further improve tissue properties.
This study focuses on the effect of static and dynamic straining on remodeling of collagen, being the main load-bearing part of cardiovascular tissues. Besides collagen, elastin plays a role in the biomechanical behavior of cardiovascular tissues, providing resilience to the tissue. Disturbances in the elastin homeostasis in heart valves and arteries are believed to represent an underlying cause of valve replacement failure 38 and the formation of aneurysms. 39 Therefore, elastin formation has been acknowledged as a missing link for a proper biomechanical function of tissue-engineered cardiovascular replacements. 40 A limited amount of studies have investigated the effect of mechanical straining on elastin formation by cardiovascular cells. Mechanical straining tends to increase elastin synthesis by vascular smooth muscle cells. Similar to collagen responses, these elastin responses seem to be sensitive to straining magnitude and mode.9,41 Ongoing studies in our lab investigate whether mechanical conditioning leads to a proper elastin network in engineered cardiovascular tissues or that also biochemical cues need to be included.
In conclusion, this study demonstrates that collagen remodeling is straining mode dependent. Static strain stimulated collagen gene expression and production with time. Dynamic straining resulted in (1) lower collagen gene expression and production, but (2) enhanced collagen cross-link expression and density, and GAG production, and (3) stimulated collagen remodeling, as expressed by enhanced production of specific collagen remodeling markers. This indicates that despite a lower collagen amount, the quality and structural integrity of the tissue are improved by dynamic straining. These straining mode–dependent remodeling responses allow us to balance collagen and cross-link production and thus to fine tune tissue properties. This is of utmost importance for cardiovascular tissue engineering, where insufficient mechanical properties are a main limiting factor for present in vivo application.
Footnotes
Acknowledgments
This research is supported by the Dutch Technology Foundation (STW), Applied Science Division of NWO, and the Technology Program of the Dutch Ministry of Economic Affairs. The authors would like to thank Jessica Snabel and Anita van de Loo for performing the biochemical assays. PCR facilities and materials were kindly provided by TNO Leiden, Department of Tissue Repair.
Disclosure Statement
No competing financial interests exist.
The work was performed at the Department of Biomedical Engineering, Eindhoven University of Technology, Eindhoven, The Netherlands.