
Abstract
For the purpose of applying fetal liver cells (FLCs) as a cell source to tissue-engineered bioartificial livers, three-dimensional (3-D) cultures of FLCs using a porous polymer scaffold, as well as monolayer cultures as a control, were simultaneously performed. To achieve efficient growth and differentiation, the FLCs were cultured in the growth medium for the first 3 weeks and then cultured in the differentiation medium for 3 more weeks. In these cultures, stimulating factors (oncostatin M (OSM), epidermal growth factor (EGF), hepatocyte growth factor (HGF), or dimethyl sulfoxide (DMSO)) were added to the media, and their effects were examined. When the growth medium containing OSM and EGF was used, EGF stimulated the growth of FLCs synergistically with OSM. For the differentiation of FLCs into mature hepatocytes, DMSO added to the differentiation medium remarkably enhanced albumin secretion in the 3-D and monolayer cultures, although HGF was effective only in the monolayer culture. Microscopic observation proved that FLCs exhibited hepatocyte-like morphology only in the media containing DMSO. In conclusion, successive supply of the growth medium containing EGF and OSM and the differentiation medium containing DMSO efficiently induced the growth of the 3-D cultured FLCs and their differentiation into mature hepatocytes.
Introduction
Over the past 2 decades, several types of bioartificial livers (BALs), which contain liver cells in BAL modules, have been developed and have succeeded in bridging to liver transplantation.6–10 However, the duration of liver support by BALs has been limited to a short period of a few days, and a superior BAL that is applicable for longer periods until the liver functions of the patients have recovered has not been developed.
To develop a superior BAL, the numbers and metabolic functions of hepatocytes within the BAL module should be maintained over a long period, and thus application of proliferative cells as the cell source of the BAL is essential. However, mature hepatocytes that have widely been used for BALs show little proliferation when cultured. Therefore, in our previous studies, long-term three-dimensional (3-D) cultures of fetal liver cells (FLCs) that are able to proliferate in vitro were performed to develop a superior BAL.11–14 These studies showed that the addition of oncostatin M (OSM), which belongs to interleukin-6 family cytokines, stimulated the growth and liver-specific function of FLCs. However, the levels of albumin secretion activities in these cultures were still lower than those of mature hepatocytes.
In the present study, two kinds of culture media (a growth medium and a differentiation medium) were successively used for FLC cultures under 3-D and monolayer conditions to achieve sufficient cell growth and efficient maturation of FLCs. The effects of the stimulating factors supplemented in these media on the growth and hepatic functions of FLCs were investigated.
Materials and Methods
Cells and culture media
FLCs were isolated from C57BL/6 mice on embryonic day 14, as previously reported.11,12 The University of Tsukuba Animal Experiment Committee guidelines for the care and use of laboratory animals were observed.
As a basal medium (BM), Williams' medium E (ICN Biomedicals Inc., Aurora, OH) supplemented with 10% fetal bovine serum (FBS, Sigma-Aldrich, St. Louis, MO), hormones (dexamethasone, 0.1 μmol/L; insulin, 0.1 μmol/L; aprotinin, 5,000 KIU/L (5.8 TIU/L); Sigma-Aldrich) and antibiotics (penicillin G, 50,000 IU/L; streptomycin, 50 μg/L; amphotericin B, 125 μg/L; Gibco BRL, Grand Island, NY) was used.12,15 To investigate the effects of the stimulating factors on the proliferation and functions of FLCs, human epidermal growth factor (EGF, 10 ng/mL; Chemicon International Inc., Temecula, CA), mouse OSM (10 ng/mL, Sigma-Aldrich), human hepatocyte growth factor (HGF, 10 ng/mL; PeproTech Inc., Rocky Hill, NJ), or dimethyl sulfoxide (DMSO, 1%; Wako Pure Chemical Industries, Ltd., Osaka, Japan) were added to the BM.
Substrate material
As a 3-D scaffold for the culture of FLCs, a highly porous reticulated polyvinyl formal (PVF) resin (Aion, Osaka, Japan) was used, as in our previous studies.11,12,15 Resin with a mean pore size of 100 μm was cut into cubes (2 × 2 × 2 mm), sterilized in an autoclave, and coated with type-I collagen before initiating the culture.11,12
Monolayer and 3-D cultures
Two types of culture experiments (3-D cultures using PVF cubes and conventional monolayer dish cultures as a control) were performed simultaneously. For the 3-D cultures, the FLCs were seeded into the collagen-coated PVF cubes using centrifugation according to the method we developed previously. 16 Briefly, 100 collagen-coated PVF cubes (0.8 cm3) and 5 mL of BM containing 5 × 107 FLCs were added to a centrifuge bottle and centrifuged at 300 × g (1 min) six times. 12 Twenty PVF cubes (0.16 cm3) containing cells were placed in a 35-mm culture dish, followed by the addition of 2.5 mL of fresh medium.
For the monolayer cultures, 1 × 106 FLCs were suspended in 2.5 mL of BM and inoculated onto 35-mm culture dishes coated with type-I collagen. For the 3-D cultures and the monolayer cultures, BM was used for the first day, and then BM with or without the stimulating factors was used. The culture medium was changed twice a week.
Effects of growth and differentiation media on FLCs
To determine the composition of the growth medium suited for FLC proliferation, the effects of OSM and EGF were examined. In these experiments, FLCs were cultured for 4 weeks in one of the growth media (BM, an EGF-containing BM (E-medium), an OSM-containing BM (O-medium), or a BM supplemented with EGF and OSM (EO-medium)).
As a differentiation medium, DMSO, HGF, or both factors were added to an E-medium (ED-medium, EH-medium, or EDH-medium), and an E-medium was also used as a control. In culture experiments to induce differentiation of FLCs, the cells were cultured in the growth medium during the first 3 weeks to increase the numbers of FLCs. Then, one of the differentiation media was supplied for 3 more weeks.
Biochemical assays and morphological examination
In the 3-D and monolayer cultures, the numbers of cells immobilized within PVF cubes or attached to culture dishes were estimated by measuring the DNA amount of these cells using the Hoechst 33342 solution.17,18 Albumin secretion was measured as an index of the liver-specific function of cultured FLCs.11,12
For morphological examination of the 3-D cultured cells, the FLCs immobilized in the PVF resin cubes were examined using scanning electron microscopy (SEM; JSM-6320F, JEOL, Tokyo, Japan). 15 In the case of monolayer cultures, the cultured FLCs were observed using a phase contrast microscope (TE300, Nikon Co., Tokyo, Japan).
Statistical analysis
Data were obtained from two to seven independent runs under each culture condition and are expressed as means ± standard deviations. Statistical analysis was carried out usng the Student t-test, and significance was set at p < 0.05.
Results
Effects of growth medium composition on growth and liver-specific functions of FLCs
EGF and OSM were chosen as the stimulating factors of the growth medium, and the effects of these factors on the growth and liver-specific functions of FLCs were examined. Figure 1 shows the time-course changes in the cell numbers of cultured FLCs. In the monolayer (Fig. 1A) and 3-D (Fig. 1B) cultures, the growth rates of FLCs were greater with the addition of OSM (O-medium) than in BM. However, no stimulating effects of EGF on cell growth were observed (E-medium). When FLCs were cultured in EO-medium, the highest proliferations of FLCs were achieved under all medium conditions tested, and the FLCs proliferated well in the EO-medium during the first 3 weeks. Thus, it was found that EGF stimulates the proliferation of FLCs only in the presence of OSM.
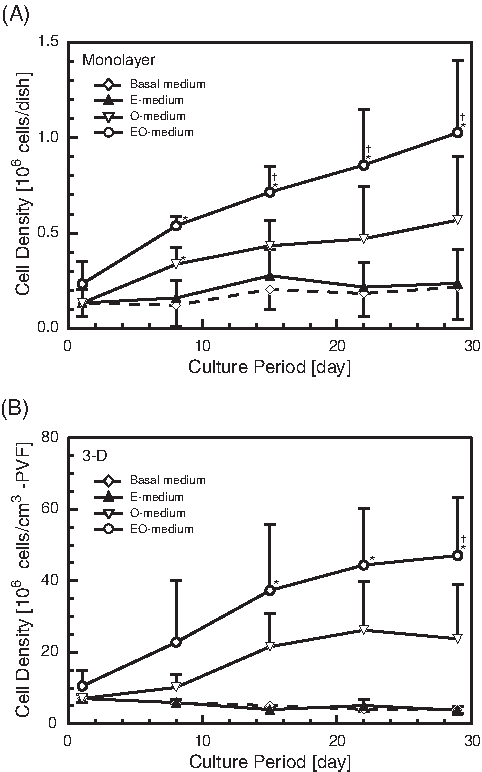
Time-course changes in cell densities of fetal liver cells cultured in growth media. Means ± standard deviations. Open diamonds, basal medium (n = 3); closed triangles, epidermal growth factor (EGF)- containing medium (n = 3); open triangles, oncostatin M (OSM)-containing medium (O-medium) (n = 3); open circles, medium supplemented with EGF and OSM (n = 7). (
With respect to albumin secretion activity, the effects of OSM and EGF were similar to those on cell growth in the monolayer and 3-D cultures (Fig. 2), and the highest activity was obtained when the EO-medium was used. The activities in the 3-D cultures were one order of magnitude greater than with monolayer cultures.
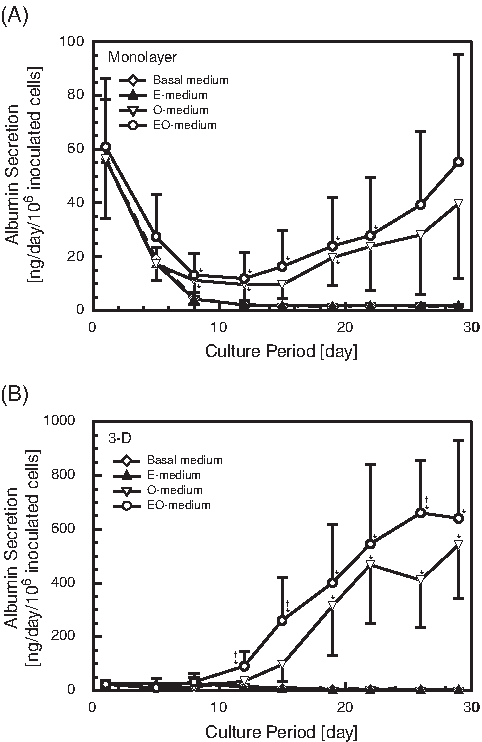
Time-course changes in albumin secretion activities of fetal liver cells cultured in growth media. Means ± standard deviations. Open diamonds, basal medium (n = 3); closed triangles, epidermal growth factor (EGF)–containing medium (n = 3); open triangles (n = 3), oncostatin M (OSM)-containing medium (O-medium); open circles, medium supplemented with EGF and OSM (n = 7). (
From these results, the EO-medium was chosen as the growth medium of FLCs in the following experiments, and this medium was applied for the first 3 weeks in the cultures.
Effects of differentiation media on cell numbers of cultured FLCs
After the FLCs were expanded in the growth medium (EO-medium), the medium was changed into one of the differentiation media, and the FLCs were cultured for 3 more weeks. The time-course changes in the cell numbers of FLCs are shown in Figure 3. When the addition of OSM was stopped (when the culture medium was changed from an EO-medium to an E-medium), the cell numbers decreased in the monolayer and 3-D cultures. In the monolayer cultures (Fig. 3A), the cell numbers also decreased in the ED-medium, but the FLCs were maintained or expanded in the media containing HGF (EH- and EDH-medium). Thus, it was revealed that HGF is essential to promoting the growth of FLCs in monolayer cultures. On the contrary, in the 3-D cultures, the cell numbers of FLCs decreased in all differentiation media, including HGF-containing media (Fig. 3B).
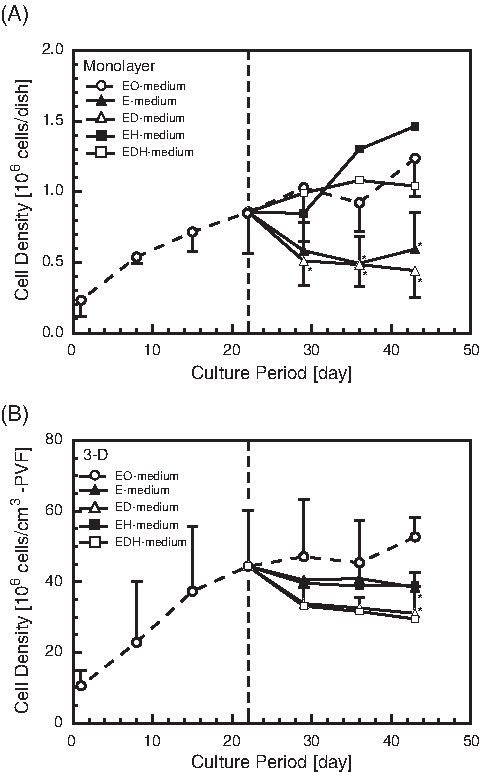
Time-course changes in cell densities of fetal liver cells cultured in differentiation media. Means ± standard deviations. Open circles, medium supplemented with epidermal growth factor (EGF) and oncostatin M (EO-medium) (growth medium, n = 7); closed triangles, EGF-containing medium (n = 3); open triangles, EGF-containing medium with dimethyl sulfoxide (DMSO) (n = 3); closed squares, EGF-containing medium with hepatocyte growth factor (HGF) (n = 2); open squares, EGF-containing medium with DMSO and HGF (n = 2). (
Effects of differentiation media on albumin secretion of cultured FLCs
The time-course changes in albumin secretion activities when the differentiation media were used are summarized in Figure 4. In the monolayer and 3-D cultures, the activities drastically increased after a medium change from a growth medium (EO-medium) to an OSM-free E-medium. However, these increases were temporary, and the elevated activities gradually decreased thereafter. When DMSO or HGF was added to the E-media (ED- and EH-medium), these factors strongly enhanced the albumin secretion of monolayer-cultured FLCs, and the elevated activities were maintained stably (Fig. 4A). Furthermore, when an EDH-medium was used, the highest albumin activity was achieved, and a remarkable increase in activity was detected. In the 3-D cultures (Fig. 4B), after a temporary increase caused by changing the medium to an E-medium, a decrease in albumin secretion was prevented by using media containing DMSO (ED- or EDH-medium). However, no recognizable effects of HGF on albumin secretion were found in these cultures, because no distinctive differences between the ED- and EDH-medium or between the E- and EH-medium were observed.
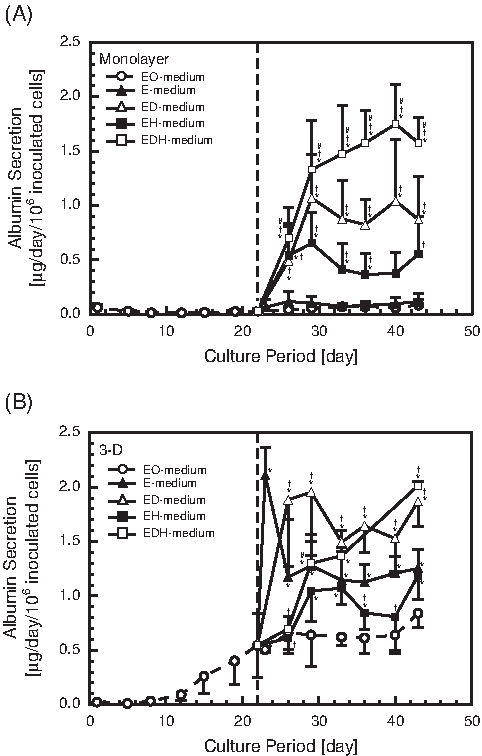
Time-course changes in albumin secretion activities of fetal liver cells cultured in differentiation media. Means ± standard deviations. Open circles, medium supplemented with epidermal growth factor (EGF) and oncostatin M (EO-medium) (growth medium, n = 7); closed triangles, EGF–containing medium (E-medium) (n = 3); open triangles, EGF-containing medium with dimethyl sulfoxide (DMSO) (ED-medium) (n = 3); closed squares, EGF-containing medium with hepatocyte growth factor (HGF) (n = 3); open squares, EGF-containing medium with DMSO and HGF (n = 3). (
In the monolayer and 3-D cultures, the highest albumin secretion activities had a similar value of approximately 2 μg/day per 106 inoculated cells. To obtain the highest activities, HGF was unnecessary in the 3-D cultures, whereas HGF was indispensable in the monolayer cultures.
Albumin secretion activities per FLC
From the data of the cell numbers and albumin secretion activities (Fig. 1–4), albumin secretion activities per FLC were calculated. Table 1 summarizes the activities when FLCs were cultured in growth media. In the monolayer and 3-D cultures, the data of albumin secretion activities per FLC measured on Day 1 seemed to include a remarkable amount of albumin released from unattached FLCs at the cell seeding. Thus, these data might be higher than the actual values on Day 1. After Day 8, the activities increased when FLCs were cultured in the O- or EO-medium under the monolayer and 3-D conditions. The albumin secretion activities per FLC were higher in the O-medium than in the EO-medium, although the activities per inoculated cell number were higher in the EO-medium (Fig. 2). Faster proliferation of FLCs in the EO-medium than in the O-medium might have caused this (Fig. 1).
In all culture experiments, basal medium was used for the first day.
P < 0.05 vs. basal medium.
In the differentiation media (E-, ED-, EH-, and EDH-medium), higher activities were obtained than in the growth medium (EO-medium) under monolayer and 3-D conditions (Table 2). Moreover, the activities were strongly stimulated when cells were cultured in the media containing DMSO (ED- and EDH-medium) under both culture conditions.
In all culture experiments, EO-medium was used for the first 21 days.
SD was not calculated (n = 2).
P < 0.05 vs. EO-medium.
Albumin secretion activities per FLC in the 3-D cultures were considerably higher in the growth and differentiation media than in the monolayer cultures (Table 1 and 2). Maximum value of albumin secretion activity per FLC obtained in the present study was approximately 4.0 μg/day per 106 FLCs (Table 2).
Morphology of FLCs in monolayer cultures
The morphology of monolayer-cultured FLCs under different medium conditions are shown in Figure 5. After cell growth in the growth medium (EO-medium) for 3 weeks, the FLCs exhibited spindle shapes like fibroblasts (Fig. 5A). When the FLCs were differentiated in the EH-medium for 3 more weeks, their morphology changed into round shapes (Fig. 5B). In cases in which DMSO-containing media (ED- and EDH-medium) were used as the differentiation media (Fig. 5C, D), the round cells with clear nuclei and dark cytoplasm constituted the majority. The morphologies of these cells were similar to those of hepatocytes. In these media, the size of each cell was smaller in the EDH-medium than in the ED-medium, corresponding to the results that higher cell densities were obtained in the former than in the latter (Fig. 3A).

Morphology of monolayer cultured fetal liver cells (FLCs) under different medium conditions. (
From these observations, it was clearly shown that the stimulating factors added affected the morphologies of cultured FLCs in the monolayer cultures remarkably. Of these factors, DMSO effectively induced changes of the FLCs into hepatocyte-like morphologies.
SEM observation of 3-D cultured FLCs
Representative SEM images of the 3-D cultured FLCs are indicated in Figure 6. When FLCs were cultured in the growth medium (EO-medium) for 3 weeks, many cells, as well as an abundant amount of extracellular matrix (ECM) secreted by the FLCs, were observed within the small pores of the PVF resin (Fig. 6B). The morphologies of 3-D cultured FLCs in various differentiation media are shown in Figure 6C-E. In these media, the 3-D cultured FLCs had a spherical morphology, although the amount of secreted ECM was different depending on the media used. In the ED-medium (Fig. 6D), the amount of accumulated ECM was less than in the growth medium (Fig. 6B), although much ECM was still observable in the HGF-containing media (Fig. 6C, E). In the EH-medium, in which the highest ECM secretion among three kinds of differentiation media was observed (Fig. 6C), albumin secretion activity was lower than in other differentiation media, suggesting that cells other than hepatocytes had expanded in this medium.
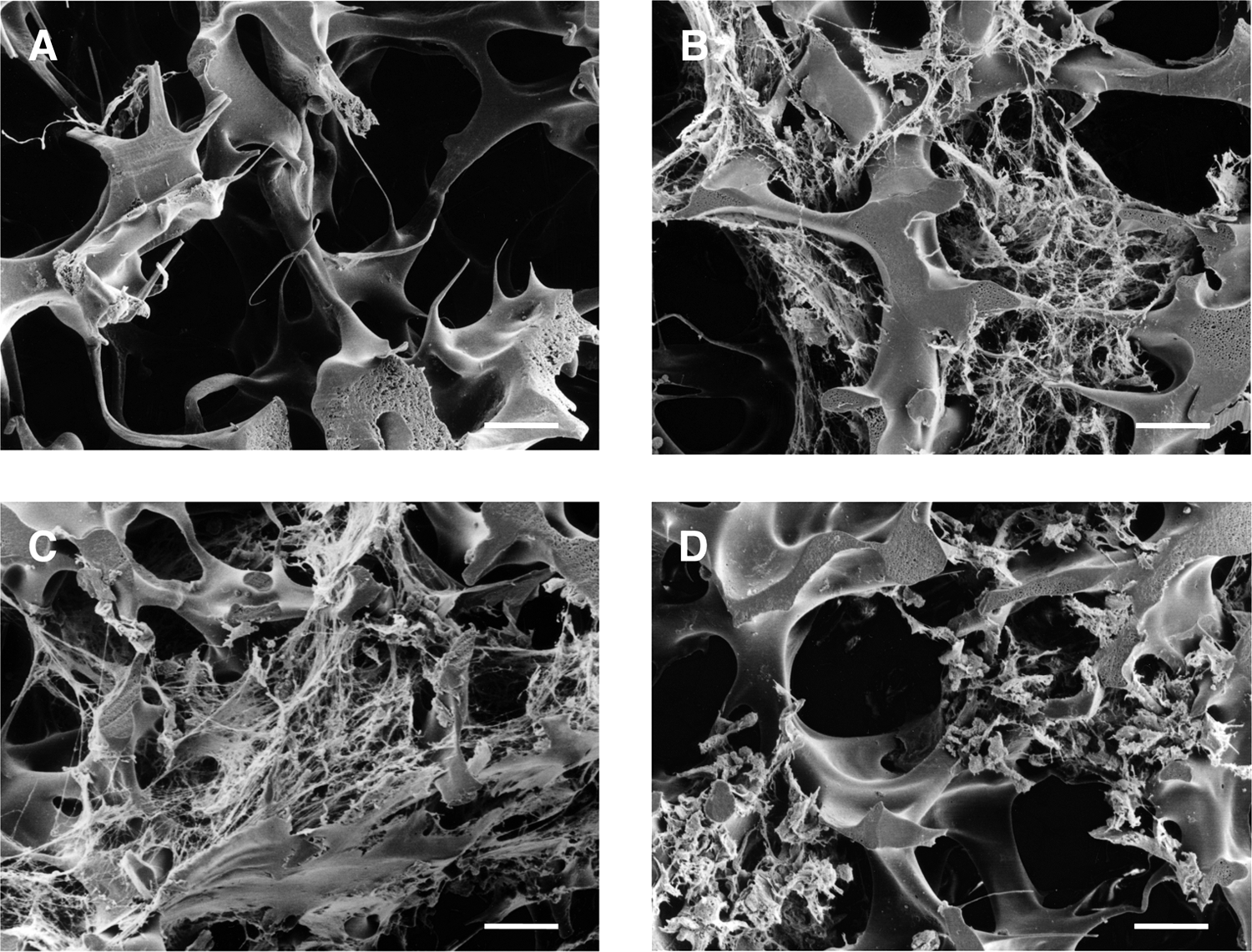
Scanning electron micrographs of fetal liver cells (FLCs) in three-dimensional cultures. (
Discussion
OSM is known as a potent stimulating factor of FLCs and has been reported to improve the cell growth and liver-specific functions of FLCs in monolayer and 3-D cultures.12,19 During mouse development, the messenger RNA (mRNA) of OSM is expressed in the liver after embryonic day (ED) 12. 19 However, OSM is scarcely detected in the liver at the late developmental stage, because hematopoietic cells that migrate from the fetal liver to the bone marrow at mid-late gestation secrete it. 20 Moreover, interleukin-6-family cytokines, including OSM, were reported to be effective at maintaining an immature state of embryonic stem (ES) cells. 21 Based on these facts, it might be possible that OSM suppresses the differentiation of FLCs into a fully differentiated state, although OSM stimulates the growth and functions of FLCs at mid gestation.
In the present study, albumin secretion activity increased dramatically after changing the medium from an OSM-containing growth medium to an OSM-free differentiation medium (Fig. 4). Thus, the suppressive effect of OSM on differentiation into a fully differentiated state was clearly demonstrated.
The EGF is secreted constantly in mice fetal livers from ED 12 to the neonatal stage. 22 The EGF receptor (EGFR) and its mRNA are detectable from ED 13.5 to 15 in developing mice livers,22–24 and the period of expression corresponds to that of gp130, a receptor of OSM. 19 In developing livers, enhanced expression of EGFR during the neonatal stage has also been elucidated.22,23 Thus, it is strongly suggested that EGF is crucial for liver development, similar to OSM, and that EGF and OSM contribute to liver development synergistically. However, few reports have examined the stimulating effects of EGF on the growth and differentiation of cultured FLCs. 25
In the present study, EGF enhanced the cell growth and albumin secretion of FLCs in the presence of OSM in monolayer and 3-D cultures (Fig. 1 and 2), and EGF was also recognized as a potent stimulating factor for FLCs. On the contrary, EGF showed no stimulating effects on FLCs in the absence of OSM. Therefore, the detailed mechanism of stimulation by EGF and OSM should be clarified by investigating the intracellular signaling pathway of these factors.
To facilitate the differentiation of FLCs into a fully differentiated state, HGF and DMSO, which are known to be effective in proliferating and maintaining the liver-specific functions of mature hepatocytes in vitro, were used in this study.26–28 With respect to the effects of HGF on liver development, the expression of HGF in the mouse liver increases with decreasing expression of OSM in the perinatal stage, and HGF has been reported to enhance the proliferation and liver-specific functions of monolayer-cultured FLCs.29,30 In the present study, similar effects of HGF on FLCs were obtained under the monolayer culture condition (Fig. 3A and 4A), although no distinct effects of HGF on FLCs were observed in the 3-D cultures (Fig. 3B and 4B).
Recently, coculture experiments of ES cells and FLCs suggested that FLCs contain cells that secrete some soluble factors inducing differentiation of immature cells into mature hepatocytes. 31 In the present study, the concentration of these soluble factors might be higher in the 3-D cultures than in the monolayer cultures, because the densities of FLCs per milliliter of culture medium were far higher in the 3-D cultures. Thus, advanced differentiation of the 3-D-cultured FLCs induced by the soluble factors was considered to reduce the sensitivity of FLCs to HGF stimulation. Moreover, it seems likely that HGF is one of the soluble factors secreted by FLCs.
The stimulating effects of DMSO on the proliferation and albumin secretion activities of FLCs were confirmed in the monolayer and 3-D cultures (Fig. 3 and 4), similar to those on mature and newborn hepatocytes.27,28 Furthermore, it was clearly shown that DMSO induced morphological changes of FLCs into mature hepatocyte-like cells (Fig. 5).
DMSO has been reported to stimulate the expression of connexin 26 (Cx26) and Cx32, gap junction proteins between mature hepatocytes. 32 The expression of these proteins in rat liver is also known to increase in the late gestation stage or after birth. 33 Therefore, it is speculated that the formation of gap junctions between liver cells is essential for the maturation of FLCs, and enhanced formation of gap junctions might have caused increases in albumin secretion of FLCs by adding DMSO in the present study.
With respect to albumin secretion activity, mature rat hepatocytes cultured within the same PVF scaffold showed albumin secretion of approximately 33 μg/day per cm3 of PVF (10 μg/day per 106 inoculated cells) in our previous study. 34 On the other hand, the maximum albumin secretion of the 3-D cultured FLCs obtained in the present study was 117.5 μg/day per cm3 of PVF (2.0 μg/day per 106 inoculated cells) (Fig. 4). Thus, higher albumin secretion activities per scaffold were obtained in the FLC cultures, whereas the activities per inoculated cells were higher in the cultures of mature hepatocytes. It was also clarified that the maximum value of albumin secretion activity per FLC was approximately 4.0 μg/day per 106 FLCs (Table 2). Therefore, maximum albumin secretion activity per fetal hepatocyte seemed to be higher than this value, because FLCs used in the present study contained fetal hepatocytes and nonparenchymal cells. These results suggest that the FLCs (fetal hepatocytes) secreted albumin on the same order of magnitude as that of mature hepatocytes, although the culture conditions, including densities of the cultured cells and frequencies of the medium exchanges, differed between the 3-D cultures of the FLCs and mature hepatocytes. Further experiments to compare the hepatic functions of the 3-D-cultured FLCs and mature hepatocytes more closely are required.
In conclusion, the proliferation and albumin secretion of FLCs were efficiently enhanced by successive supply of the growth medium containing OSM and EGF and the differentiation medium containing DMSO. Moreover, high albumin secretion activities were achieved in the 3-D cultures even in the differentiation media without HGF. Thus, the 3-D culture methods of FLCs used in this study seem effective to apply immature cells as cell sources to tissue-engineered bioartificial livers.
Footnotes
Acknowledgments
This work was supported by a grant from the Japanese Ministry of Education, Culture, Sports, Science and Technology. The authors also appreciate the advice of Dr. Keiko Ookawa.
Disclosure Statement
No competing financial interests exist.