
Abstract
Peripheral nerve regeneration within guidance conduits involves a critical association between regenerating axons, Schwann cells (SCs), and neovascularization. However, it is currently unknown if there is a greater association between these factors in nonpermeable versus semipermeable nerve guide conduits. We therefore examined this collaboration in both silicone- and collagen-based nerve conduits in both 5- and 10-mm-injury gaps in rat sciatic nerves. Results indicate that collagen conduits promoted enhanced axonal and SC regeneration and association when compared to silicone conduits in the shorter 5-mm-gap model. In addition, collagen tubes displayed enhanced neovascularization over silicone conduits, suggesting that these three factors are intimately related in successful peripheral nerve regeneration. At later time points (1- and 2-month analysis) in a 10-mm-gap model, collagen tubes displayed enhanced axonal regeneration, myelination, and vascularization when compared to silicone-based conduits. Results from these studies suggest that regenerating cables within collagen-based conduits are revascularized earlier and more completely, which in turn enhances peripheral nerve regeneration through these nerve guides as compared to silicone conduits.
Introduction
Axons and Schwann cells (SCs) display an intimate relationship in both an uninjured environment and after tubulization following peripheral nerve transection, which allows for SCs to have a very direct and specific effect on regenerating axons.19,20 For example, when regenerating axons contact growth supportive SCs, they release neuregulins from their growth cones that bind to erbB receptors on the SCs, mediating additional SC proliferation.21,22 Proliferating SCs, in turn, are a rich source of neurotrophins that further enhance axonal regeneration. 23 Local vascular changes such as angiogenesis or neovascularization (endothelial cell proliferation and new vessel formation) also play an important role in nerve regeneration through providing oxygen, nutrients, and other factors necessary for supporting axonal regeneration.24–26 For example, Weddell (1942) 27 was the first to suggest that axonal regeneration was intrinsically tied to large vessel neovascularization. Despite this suggested interaction, it is currently unknown how the degree of neovascularization influences axon–SC association and regenerative processes in either silicone- or collagen-based nerve guide conduits.
In the present work, we attempted to answer these questions by examining early regenerative axon–SC association in a short-gap (5 mm) model and investigated how these regenerative processes were influenced by early neovascularization in both silicone and collagen conduit–repaired sciatic nerve injuries. More specifically, we asked whether axonal, SC, and blood vessel profiles were increased in collagen-based as opposed to silicone-based conduits, and whether there was a higher degree of association between these three parameters in either of the tubes tested. Quantitative double label immunohistochemistry was employed to investigate early regenerative events. We then further examined differences in both axonal myelination and blood vessel profiles between silicone and collagen conduits over longer periods of time (1- and 2-month evaluations) in a 10-mm-gap nerve injury model through histomorphometric analysis. This length of nerve injury gap was chosen because a regenerating nerve cable will nearly always bridge the gap, given sufficient time, with a distance of 10 mm or less using a silicone-based conduit in rats, whereas there is little to no regeneration successfully across gaps exceeding 10–15 mm in rats without the introduction of exogenous factors such as growth factors or SCs.9,28 Results indicate that collagen nerve guide conduits promoted enhanced axonal regeneration, SC association, and neovascularization as compared to silicone-based conduits in an early model of peripheral nerve regeneration. Further, axonal regeneration, myelination, and vascularization were superior in collagen as opposed to silicone conduits over both 1- and 2-month periods in a 10 mm nerve injury gap paradigm. Overall, our results suggest that collagen conduits provide a more conducive environment for peripheral nerve regeneration in both 5- and 10-mm-gap models.
Materials and Methods
Animals
Adult male Lewis rats (52 in total), weighing 250–300 g (Charles River, QC), were used in this study, and all interventions were carried out under inhalation anesthetic (Isoflurane, 99.9%; Halocarbon Laboratories, River Edge, NJ). Animals were maintained in a temperature- and humidity-controlled environment, and were allowed standard rat chow (Purina, Mississauga, ON) and water ad lib, with a 12:12 h light:dark cycle. All surgical procedures were carried out in an aseptic manner, and standard microsurgical techniques were used with an operating microscope (Wild M651; Wild Leitz, Willowdale, ON). Animals were sacrificed at endpoint, under deep anesthesia, using an overdose of intracardiac Euthanol (Bimeda-MTC, Cambridge, ON). The protocol was approved by the University of Calgary Animal Care Committee and adhered strictly to the Canadian Council on Animal Care guidelines.
Surgical procedure and experimental design
Following induction of anesthesia, surgical sites were shaved and prepared with Betadine (Rougier Pharma, Mirabel, QC, Canada) and 70% surgical alcohol. Following a dorsal thigh incision in all animals, the right sciatic nerve was exposed, dissected, and transected at mid-thigh level with an 11-0 scalpel blade. Proximal and distal nerve stumps were then secured into either a silicone (1.98 mm internal diameter) or a collagen (1.5 mm internal diameter) nerve guide conduit and sutured in place with two 9-0 sutures (Ethicon, Somerville, NJ). For the short-gap model, sciatic nerves were transected, and then proximal and distal nerve stumps were immediately secured 1 mm into 7-mm-long conduits, creating a gap of 5 mm. For the longer-term injury model, sciatic nerves were transected, and a 7-mm segment of the nerve was subsequently excised. Proximal and distal nerve stumps were then secured 1 mm into 12-mm-long conduits, which resulted in a 10-mm gap between the ends of the transected nerve. Collagen nerve guide conduits were fabricated (a gift from Integra Life Sciences, Plainsboro, NJ) from highly purified type I collagen, derived from bovine deep flexor tendon. Previous studies 29 have shown that these collagen nerve guide conduits are freely permeable to macromolecules, with molecular weights as large as bovine serum albumin (MW = 68 kDa). For the short-gap model (5 mm), outcome measures were analyzed at both 1- and 2-week time points. For the 10-mm-gap model, outcome measures were analyzed at both 4- and 8-week time points.
Immunohistochemistry
At both 1- and 2-week time points following initial surgery, the sciatic nerve (approximately 2–3 mm proximal to initial transection) and the regenerating cable within either silicone or collagen conduits were removed for longitudinal sections through both the proximal stump and regenerative bridge. Tissue sections were fixed in modified Zamponi's fixative (2% paraformaldehyde, 0.5% picric acid, and 0.1% phosphate buffer) and stored overnight at 4°C. Nerve samples were then washed three times in PBS, cryoprotected in 20% sucrose/PBS, and then left overnight at 4°C. Tissue samples were then embedded in optimal cutting temperature compound (Sakura Finetechnical, Torrance, CA), and 12-μm-thick sections were placed onto poly-L-lysine–coated slides. For indirect immunofluorescence, slides were incubated for 24 h at 4°C with primary antibodies anti-NF200 (marker for the heavy subunit of neurofilament in axons, 1:800, NO142; Sigma, St. Louis, MO), anti-GFAP (SCs, 1:250; Dako Diagnostics, Mississauga, ON), and RECA-1 (rat endothelial cells, 1:50; Serotec, Kidlington, United Kingdom). 25 Slides were then washed with PBS and incubated with secondary antibodies for 1 h at room temperature. For double labeling, primary antibodies raised in two different species were incubated together at 4°C for 48 h, followed by appropriate combination of secondary antibodies conjugated to either Alexa Fluor 488 (Invitrogen, Carlsbad, CA) or Cy3. After further washing with PBS, cover slips were mounted with Flurosave reagent (Calbiochem, San Diego, CA) and viewed under a fluorescent microscope (Olympus BX51, Center Valley, PA). Negative controls involved omission of either the primary or secondary antibody on parallel sections.
Analysis of early peripheral nerve regeneration
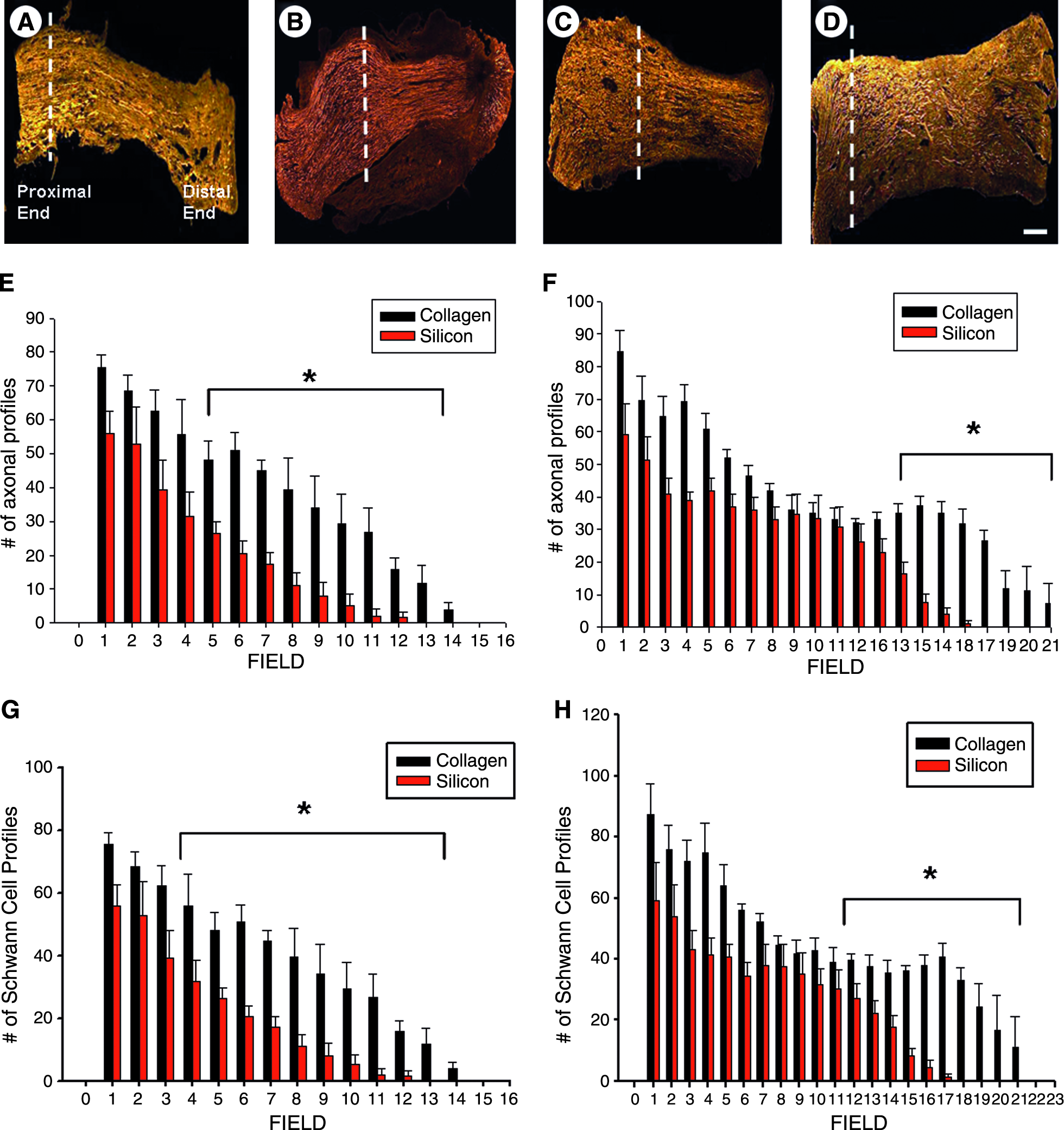
The method employed for assessing early axonal regeneration (1- and 2-week time periods) was similar to the procedure used by Chen et al. (2005). 20 Briefly, axonal profiles stained for neurofilament were easily distinguishable in longitudinal sections containing both the proximal stump and regenerative bridge, and were clearly delineated in all samples observed. Beginning at the distal end of the proximal stump (first separate high power field: 400×; each field was 270 μm in diameter), axonal profiles (NF200) or SC profiles (GFAP) were counted (by an observer blinded to treatment groups) in a line perpendicular to the direction of the bridge in serial, high power fields distally every 270 μm until the field contained no positive neurofilament or GFAP staining. For each individual field, we counted positive NF200 and GFAP profiles that transected the middle of the section. Both NF200- and GFAP-positive profiles were subsequently summed for each field. A representative sample is shown in Figure 1 displaying 12 fields, although this varied between animals and was dependent on the length of the regenerating cable (maximum number of fields containing labeled profiles observed was 21). In another set of samples, RECA-1–positive profiles were counted and expressed per entire nerve cable. Our inclusion criteria for a positive RECA-1 profile consisted of the presence of a fluorescent-lined (denoting RECA-1–positive endothelial cells), cylindrical blood vessel lumen (Fig. 4A), consistent with prior investigators. 25

Harvesting and early peripheral nerve regeneration counting method. (
Vascular morphometry
Vascular morphometry was studied at the 1- and 2-week time points using a modified version of Bray's method. 30 Briefly, at 1 and 2 weeks, five animals in each group were transcardially perfused with 20 mL of a solution of India Ink (25%), gelatin (4%), and mannitol (5%) at 37°C (rate = 2 mL/min). After perfusion, the rats were euthanized and placed in a freezer at −20°C for 1 h before removal of the regenerated nerve cable. This method allows for the outlining of vascular lumina, without noticeable leakage of India Ink into the surrounding tissue. Specimens were then fast frozen (isopentane in dry ice) in optimal cutting temperature compound and sectioned at 12 μm, with sections placed (without fixative) on poly-L-lysine–coated slides. All samples were analyzed at the level of the middle portion of the regenerating cable. Five sections of each nerve were used to calculate a mean value for each rat. Total transverse nerve area, vascular area, and total vessel numbers were measured using Image Pro Plus 5 (Media Cybernetics, Bethesda, MD). The approach and quantification have been previously reported by one of our laboratories.26,31
Histomorphometry
Sciatic nerve regenerative cables were harvested at both 4 and 8 weeks after transection and surgical repair. Samples were then fixed in glutaraldehyde (2.5%), buffered in cacodylate (0.025 M) overnight, washed, and then stored in cacodylate buffer (0.15 M). Samples were subsequently fixed in osmium tetroxide (2%), washed in graded alcohols, and then embedded in Epon. Transverse sections at 1 μm thickness were made through the approximate center of the regenerative bridge and stained with toluidine blue.
With the observer blinded to the identity of the groups, sections of the regenerating bridge, approximately at its center, were photographed at high-power light microscopy (400×). Photographs were taken of the entire nerve cable, with each individual section being used in the final analysis using Image Pro Plus software. The parameters measured included (1) total number of myelinated fibers, (2) fiber diameter, (3) total fiber area, (4) whole nerve area, and (5) blood vessel profiles/nerve cable.
Statistical analysis
Differences between groups were compared using a one-way ANOVA, with post hoc Student's t-tests applied as appropriate. Statistical significance was accepted at the level of p < 0.05, with all results presented as the mean ± SEM.
Results
Collagen nerve guides promote enhanced axonal and SC profiles during early peripheral nerve regeneration
Axonal and SC profiles labeled with both neurofilament and GFAP were easily distinguished from the proximal trunk extending through the region of regenerating cable, and the appearance of samples closely resembled previous findings (Fig. 2A–D).19,20 At the 1-week time point, regeneration in silicone conduits did not proceed past field 12, whereas axonal and SC profiles extended through field 14 in collagen conduits. Results indicated that animals with collagen conduits were significantly different from silicone animals, beginning at field 5 for axonal profiles [t(8) = 3.13, p < 0.05], and SC profiles beginning at field 4 [t(8) = 4.67, p < 0.01; Fig. 2E, F]. At the 2-week time point, regeneration in silicone conduits did not proceed past field 17, whereas axonal and SC profiles extended through field 21 in collagen-based conduits. Results indicated that animals with collagen conduits were significantly different from silicone animals, beginning at field 14 [t(10) = 3.92, p < 0.01], and SC profiles beginning at field 11 [t(10) = 8.10, p < 0.01; Fig. 2G, H]. Therefore, animals repaired with collagen-based conduits displayed a significantly greater number of both axonal and SC profiles at both 1- and 2-week time points. In addition, these profiles extended further through the regenerating cable in animals whose sciatic nerves were repaired with collagen-based as opposed to silicone-based conduits.
Axonal-labeled (NF200) and SC-labeled (GFAP) profiles in a short-gap regeneration model. Neurofilament-stained axonal profiles in (
Collagen nerve guides promote enhanced axonal–SC association during early peripheral nerve regeneration
Consistent with previous research, the majority of NF200 axonal profiles were closely associated with GFAP-labeled SCs in both silicone and collagen conduits (Fig. 3A, B). 20 However, animals whose injuries were repaired with collagen conduits showed a greater number of axon–SC associated profiles in a bimodal fashion. Animals with collagen conduit repair at the 2-week time point showed a statistically significantly higher number of associated profiles from fields 1–6 [t(8) = 5.80, p < 0.01] and from fields 13–21 [t(8) = 2.80, p < 0.01]. Naked axons were defined as axonal processes that were not associated with a corresponding SC profile. Previous research has shown that these types of axonal profiles do not extend throughout the regenerated cable, most often terminating in proximal or mid cable. 20 We therefore analyzed the number of naked axons in regenerating cables of both silicone and collagen conduits at the proximal, mid, and distal cable regions. Because regenerating profiles did not extend beyond field 21 in either silicone or collagen conduit–repaired animals, we designated fields 1–7 as the proximal region of the regenerating cable, fields 8–14 as the mid-cable region, and fields 15–21 as the distal region of the regenerating cable. Regenerating cables through silicone conduits displayed a significantly greater number of naked axons at both proximal [t(11) = 2.56, p < 0.05] and mid cable [t(12) = 2.74, p < 0.05] at the 2-week time point. This was further supported by the fact that collagen conduits displayed a significantly greater percentage of axon–SC association per nerve cable at both proximal [t(12) = 2.56, p < 0.05] and mid cable levels [t(12) = 2.74, p < 0.05; Table 1]. Axon–SC associated profiles did not reach the distal stump in either treatment group.
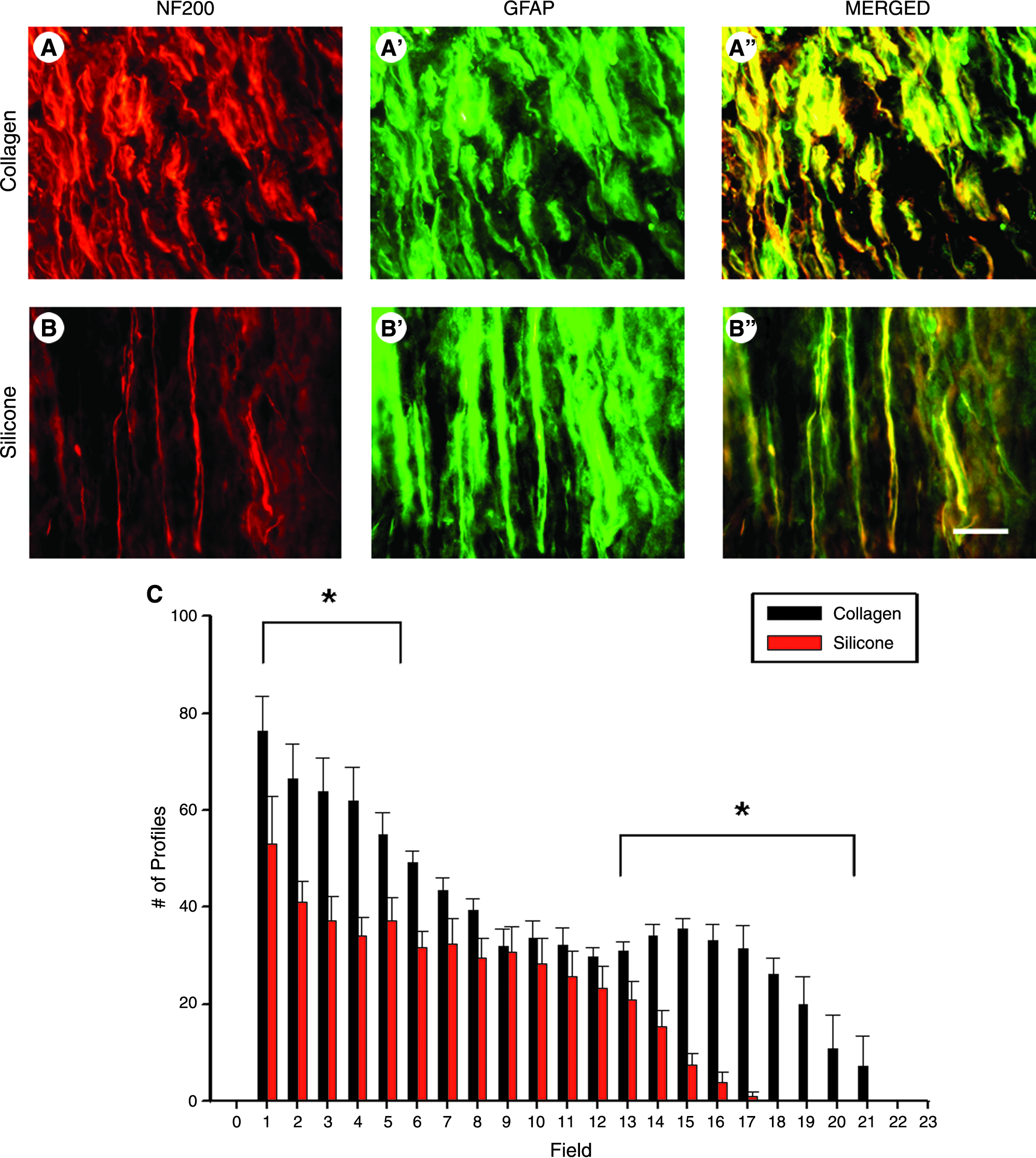
Assessment of neurofilament-stained axonal profiles associated with GFAP-positive SC profiles in collagen (

Collagen nerve guide conduits promote enhanced neovascularization as analyzed through RECA-1 immunofluorescence. (
Tabulated are the numbers of naked axons and percentage of axon–SC association per regenerating nerve cable.
p < 0.05.
Collagen nerve guides display enhanced early neovascularization compared to silicone tubes
RECA-1
At both 1- and 2-week time points, endothelial cells stained with RECA-1 were clearly visible, with blood vessels presenting with well-defined cylindrical lumens (Fig. 4A). At the 1-week time point, both axonal and SC profiles did not extend beyond the area of RECA-1–positive profiles. At the 1-week time point, animals whose injuries were repaired with collagen conduits displayed a significantly greater number of RECA-1 profiles than animals repaired with silicone conduits [t(8) = 3.58, p < 0.01; Fig. 4B]. Although RECA-1–positive profiles were higher in the collagen group as compared to the silicone group at 2 weeks, there was no statistical difference between the groups at this time point.
India Ink
Vascular morphological analysis was performed using a modified India Ink/gelatin perfusion technique. 30 This method allows for visualization of blood vessels within the regenerating cable and nerve morphology, which is not subsequently affected by postmortem fixation (Fig. 5A–D). At 1-week postsurgery, there was a significant increase in blood vessel profiles expressed by the collagen group as compared to the silicone group [t(4) = 2.37, p < 0.05], as analyzed at the level of the middle portion of the regenerating cable. At 2 weeks, collagen conduit groups again displayed a significantly greater number of blood vessel profiles [t(4) = 2.30, p < 0.05; Fig. 5E]. At both 1- and 2-week assessment points, collagen conduit–repaired animals displayed a greater total vascular area percentage per regenerating nerve cable than silicone-repaired animals (8.80 ± 0.57 vs. 5.74 ± 0.54 at 1 week, and 17.34 ± 1.41 vs. 12.6 ± 0.89 at 2 weeks). These differences were statistically significant at both 1-week time point [t(4) = 3.93, p < 0.05] and 2-week time point [t(4) = 2.89, p < 0.05; Fig. 5F]. Therefore, animals whose injuries were repaired with collagen conduits displayed both a significantly greater number of blood vessels and greater total vascular area percentage per regenerating nerve cable than silicone conduit–repaired animals.

Unfixed transverse sections of India Ink–perfused rat sciatic nerves. (
Collagen nerve guides promote enhanced axonal myelination and vascularization in a 10-mm nerve gap injury model
In a short-gap model (5 mm), collagen conduits were shown to promote enhanced axonal regeneration, SC association, and neovascularization at both 1- and 2-week time points. We next assessed whether collagen-based conduits enhanced axonal regeneration and vascularization in a 10-mm nerve gap injury model. For this, histomorphometrical analysis was performed at mid cable at both 4- and 8-week time points (Fig. 6). As shown in Figure 7D, regenerating cables through collagen-based conduits were more robust than silicone-based conduits, displaying a significantly greater nerve area (regenerating cable area) at both 4-week time point [t(6) = 4.87, p < 0.01] and 8-week time point [t(6) = 8.58, p < 0.01]. Animals with collagen conduits displayed a significantly greater mean fiber diameter than silicone conduit animals at both 4-week time point [t(6) = 7.27, p < 0.01] and 8-week time point [t(6) = 8.70, p < 0.01; Fig. 7A]. The overall fiber diameters were greater at 8 weeks than at 4 weeks presumably due to increased maturation of the nerve fiber. These animals also displayed a significantly greater fiber area at 4-week time point [t(6) = 8.92, p < 0.01] and 8-week time point [t(6) = 4.27, p < 0.05; Fig. 7C]. In addition, these animals also displayed an increase in myelin area (data not shown); however, the average g-ratio (ratio of axon diameter to fiber diameter) was not significantly different between groups. Blood vessel profiles were significantly greater in number within collagen conduit animals than within silicone-repaired animals at both 4-week time point [t(8) = 4.01, p < 0.01] and 8-week time point [t(8) = 4.06, p < 0.01; Fig. 7B]. At both 4- and 8-week assessment points, collagen conduit–repaired animals displayed a dramatic increase in the total number of myelinated fibers as compared to silicone-repaired animals (2106 ± 170.74 vs. 939 ± 124.42 at 4 weeks, and 5080.25 ± 178.20 vs. 2486.50 ± 240.67 at 8 weeks). This difference was significant at both 4-week time point [t(6) = 5.53, p < 0.01] and 8-week time point [t(6) = 8.66, p < 0.01; Fig. 7E]. Therefore, data from the 10-mm-gap studies mirror those found in the 5-mm-gap studies, with animals whose sciatic nerve injuries were repaired with collagen-based conduits displaying enhanced axonal myelination, regeneration, and vascularization as compared to silicone-based conduits at both time points evaluated.
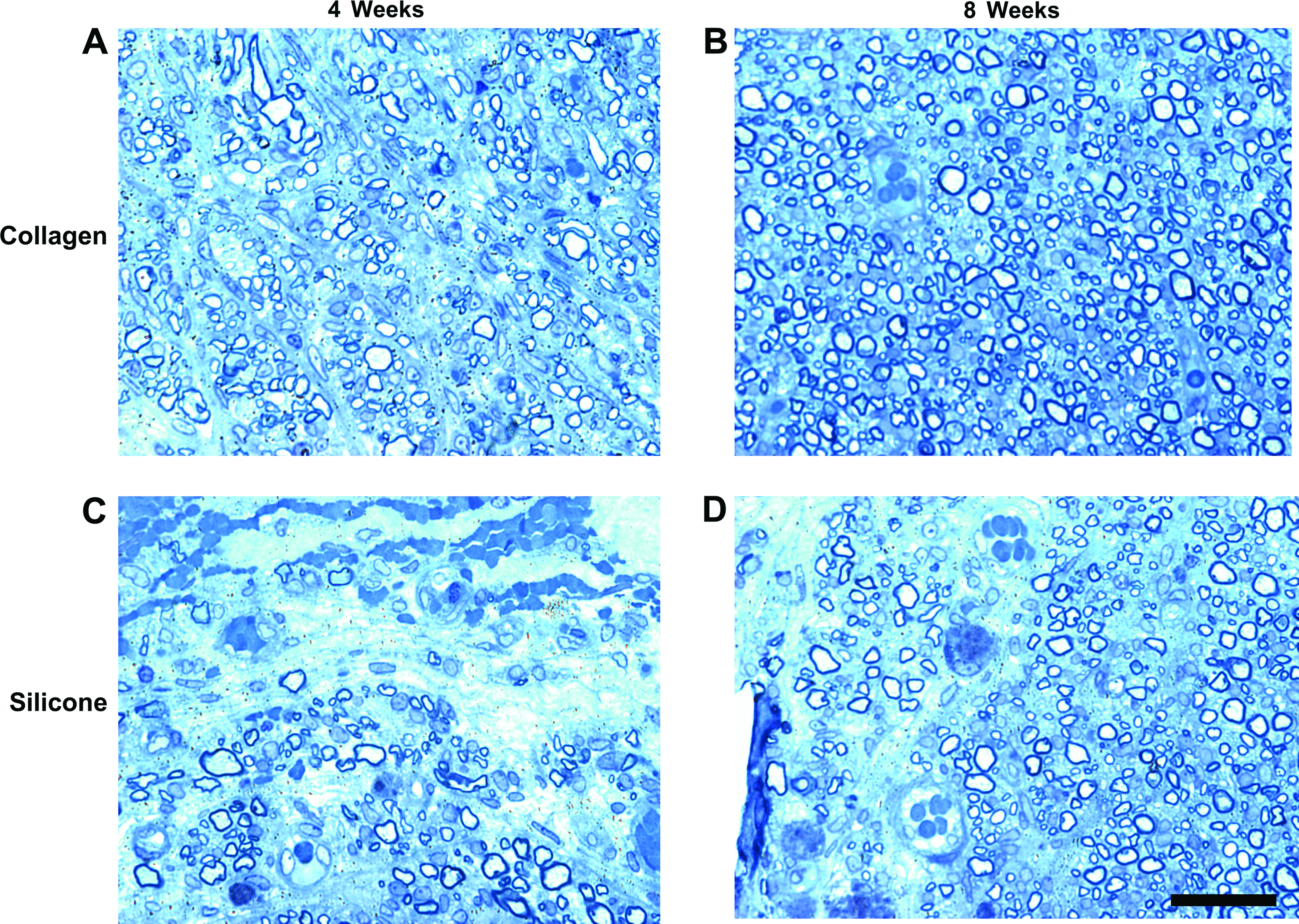
Representative cross section of the mid section of regenerating cables from collagen conduits (
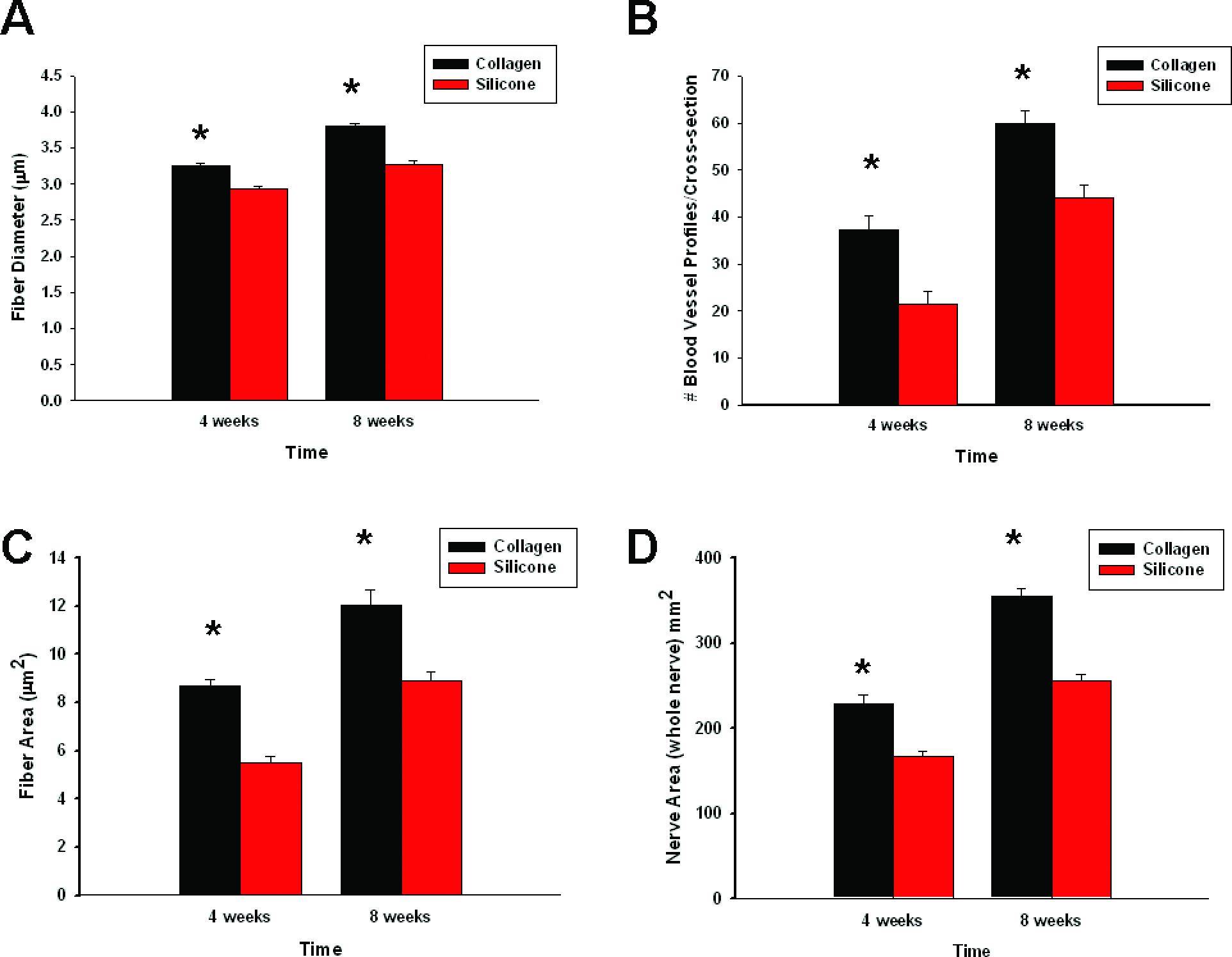
Summary of 4- and 8-week histomorphometry data for both collagen and silicone conduits: (
Discussion
Major findings
The major findings in this study were (1) collagen nerve conduits promoted enhanced axonal regeneration as compared to silicone conduits in a short-gap model at both 1- and 2-week time points; (2) collagen conduits promoted enhanced SC infiltration as compared to silicone conduits in a short-gap model at both 1- and 2-week time points; (3) this, in turn, led to a greater degree of association between axonal–SC profiles and early regeneration, with very few naked axons extending into the distal portions of the nerve conduits; (4) collagen nerve conduits promoted enhanced neovascularization as compared to silicone conduits as measured through both RECA-1 (rat-specific endothelial cell marker), and India Ink (blood vessel lumen) measures; and (5) collagen nerve conduits promoted enhanced axonal regeneration, myelination, and vascularization as compared to silicone conduits in a 10-mm nerve injury gap model at both 4- and 8-week evaluation points. Data from studies of short gap, early peripheral nerve regeneration suggest that axonal–SC–blood vessel association is related to proximal stump regenerative events as opposed to distal processes. Further, data from long-gap studies closely mirror results found in the early regenerative experiments, suggesting that enhancement of early peripheral nerve regeneration is crucial to longer-term regenerative success.
Tube dynamics: silicone- versus collagen-based conduits
Collagen and silicone are biomaterials that possess different intrinsic properties. One important dynamic to consider in nerve conduit design is the permeability of the conduit.16,32,33 Permeability is an important property of the conduit material because as the axons regenerate through the conduits, the metabolic activities of the cells necessitate the exchange of nutrients, growth-promoting factors, and wastes between the inner and outer lumen of the conduit, and has been previously shown to influence the results of axonal regeneration.34–37 The silicone-based conduits utilized in this study are impermeable, while the collagen conduits are freely permeable to macromolecules, with molecular weights as large as bovine serum albumin (MW = 68 kDa). 29 Because neurotrophic cues provided from the distal stump have been previously shown to play a significant role in nerve regeneration through both silicone and collagen nerve guides,10,38,39 conduit permeability could have led to the significant difference and subsequent increase in axon, SC, and blood vessel profiles seen in our studies between collagen and silicone conduits. This corroborates previous work examining short-term nerve regeneration, where den Dunnen et al. 40 have argued that enhanced growth rate and quality of nerve regeneration in permeable poly (DL-lactide-ɛ-caprolactone) conduits as compared to silicone conduits were directly related to the diffusion of nutrients and growth-promoting factors facilitated by the permeability of the conduits.
Recently, the pressure cuff theory has been postulated to interpret differences seen in nerve regenerative parameters between silicone- and collagen-based nerve conduits.41–43 The theory was developed from observations that a capsule of contractile fibroblasts (myofibroblasts) always forms around nerve trunks repaired and regenerated through silicone conduits, but infrequently through collagen-based conduits.42,44 For example, thick myofibroblast capsules (approximately 15–20 cell layer thickness) formed around regenerative cables in silicone-based conduits, whereas the thickness of these capsules surrounding regenerative cables through collagen conduits was dramatically reduced (approximately one to two layers).43,45 The primary premise of this theory is that myofibroblast capsules compress regenerating nerve stumps and therefore prevent a regenerating nerve cable of large diameter to be formed. Although we did not harvest both the connective tissue capsule and regenerating cable for processing, qualitative observations showed thick connective tissue capsules surrounding all silicone-based conduits, whereas a thin connective tissue capsule was seen in only one collagen-based conduit. A thicker myofibroblast capsule and therefore greater contraction of nerve cables regenerated through silicone conduits would explain the dramatic decrease in cable area seen through these conduits when compared to those regenerated through collagen-based conduits. This decrease in both regenerated nerve diameter and area subsequently leads to a corresponding decrease in axonal, SC, and neovascular profiles.
In summary, we believe that the improved permeability of the collagen conduit was the main factor responsible for the differences in regeneration encountered at the relatively early (1–2 month) time points evaluated. The possible contraction of the silicone conduit may have been an additional factor in decreasing the regenerating cable size and thereby regenerative success in the silicone as compared to the collagen conduit. At these early time points, biodegradation would not have been an important factor, although this conduit dynamic must be considered in longer term studies as the rate of degradation may affect swelling properties through increases in osmotic pressure of the conduit. 46 Silicone tubes are nonbiodegradable and have presented long-term problems clinically, including constriction and thereby compression of the nerve within their lumen, resulting in a subsequent surgery for removal of the conduit.13,14 However, the collagen-based conduits used in these studies are biodegradable, with their bioresorption being complete and producing minimal scar tissue or inflammatory responses.15,28 Because of this property, there is less likelihood of entrapment of the anastomotic site due to scarring that can lead to compression neuropathies with nonbiodegradable materials such as silicone.
Axonal–SC association is directly related to the degree of peripheral nerve regeneration
Following transection injury to a peripheral nerve, numerous changes in the behavior of SCs help to model the subsequent regenerative microenvironment such as increased proliferation, changes in gene expression, and a phenotypical change from myelinating to growth supportive.47,48 Following injury, SCs can release factors that aid regeneration in an autocrine (acting on themselves) or paracrine (acting directly on axons) fashion. 23
Following injury, we observed the expression of both GFAP-positive SC profiles and neurofilament (NF200)–positive profiles in regenerating nerve cables harvested from both silicone and collagen-based conduits. However, not only did collagen conduits display greater numbers of both axonal and SC profiles than silicone conduits at both 1 and 2 weeks, but they also extended much further within the regenerating cable (Figs. 2 and 3). Axon–SC profiles appeared to display a very close and intimate relationship, characteristic of what Chen et al. have previously termed “the axon–SC dance.” 20 This association is strikingly evident when we consider both naked axonal profiles (axons that are not accompanied by an SC process) and the percent of axon–SC association per regenerating nerve cable (Table 1). Animals whose injuries were repaired with silicone conduits displayed a greater percentage of naked axons per field, and also displayed a lower percentage of axon–SC association at both the proximal and mid cable portion of the regenerating cable. Because axon growth without an accompanying SC process has been previously shown to limit the distance of axonal regeneration, 20 the high number of naked axons at both the proximal and mid portion of the cable in silicone-repaired animals ultimately led to a lower number of regenerating axons in the distal cable in silicone-treated animals. In contrast, animals treated with collagen conduits displayed a much lower percentage of naked axons per field at both proximal and mid cable, in addition to displaying a significantly greater axon–SC association per nerve cable. These results suggest that an early and intimate relationship between regenerating axonal and SC profiles in collagen conduits enhanced regeneration in these animals.
Neovascularization: role of blood vessel interaction with axon–SC profiles
The peripheral nerve trunk's vascular supply, or vasa nervorum, plays an important role in the regenerating microenvironment of transected peripheral nerves,24,26 although the specific inter-relationship between neovascularization and axonal regeneration is not completely understood. Previous in vitro studies have shown that cultured endothelial cells form capillary-like structures when placed either on or within collagen matrices, suggesting the neoangiogenic potential of this biomaterial. 49 When we consider the in vivo situation, synthetic guidance conduits are different from nerve autografts in that they are initially avascular, which ensures that all vessels seen within the resulting regenerating cable are neoangiogenic in nature and are unhindered by the presence of an existing vascular network. 25 In addition, it has been previously suggested that neovascularization within a silicone conduit environment proceeds in a traveling wave form that increases as it proceeds proximodistally along the regenerating cable. 24
In our studies, we observed a significant increase in RECA-1–positive profiles at the 1-week time point in collagen-repaired animals as opposed to silicone-repaired animals (Fig. 4). Although collagen-treated animals displayed a higher number of RECA-1 profiles at 2 weeks, this number was insignificant (although statistical results approached significance, p = 0.06). However, analysis of India Ink studies demonstrated that collagen-repaired animals displayed a significant increase in positive profiles at both the 1- and 2-week time points, consistent with blood vessel increases previously shown over a 2-week period26,31 (Fig. 5E, F). The benefit of this in vivo perfusion model is that microvessel areas can be outlined without the influence of fixatives that might distort vessel anatomy and caliber, and samples are therefore not affected by postmortem fixation. This fixation difference between the two methods employed provides a possible explanation for why a statistically significant difference was not seen with RECA-1 staining between collagen and silicone conduits at the 2-week evaluation time point. The results of these studies suggest that regenerating nerve cables through collagen-based conduits display increased histological evidence of new microvessel formation as compared to silicone conduits, and are consistent with findings of increased angiogenic profiles of regenerating nerve through both fibronectin mat conduits 25 and direct repair methods. 26 These results are encouraging because we would expect to see enhanced regeneration in both the direct repair and fibronectin mat repair methods as compared to a tubular repair environment due to both the shorter distance regenerating axons must traverse (direct repair) and substrate guidance (fibronectin mats). Although our methods allow for an accurate assessment of the morphological relationship between blood vessel profiles, regenerating axons, and SCs, vascular morphology in and of itself does not necessarily correlate to blood flow as measured through such techniques as microelectrode hydrogen clearance polarography and laser Doppler flowmetry. However, previous work from the Zochodne lab has shown that increases in local endoneurial and extrinsic blood flow or hyperemia occurred in correlation with increases in India Ink–positive profiles at the 2-week time point analysis. 26
The time relationship between increases in angiogenesis, axonal, and SC profiles suggests that these events may share common trophic signals that lead to an enhanced regenerative response within collagen conduits. Two possible shared neurotrophin candidates are nerve growth factor (NGF) and vascular endothelial growth factor (VEGF). NGF, generated locally from infiltration by macrophages that in turn stimulates endogenous SC NGF production, 50 supports both neuron survival and directed growth of regenerating nerve fibers to their target-end organ.51,52 NGF also stimulates angiogenesis, and topical nerve growth factor is currently used clinically to promote healing in human diabetic lower extremity wounds. 53 VEGF is one of the most studied angiogenic factors, and its receptors VEGFR-1 (Flt-1) and VEGFR-2 (KDR/Flk-1) are expressed by both endothelial cells of the vasa nervorum and by SCs.54,55 Exogenous application of VEGF to regenerating peripheral nerves stimulates axonal outgrowth, promotes invasion of SCs, and leads to dramatic increases in neovascularization.25,56,57 In addition, Hoke et al. (2001) 26 have previously shown that upregulation of VEGF mRNA occurs in response to sciatic nerve transection and repair at a 2-week analysis point. More importantly, this data corresponded well to the histological evidence of new microvessel formation as measured through India Ink analysis. This data supports the idea that the mechanism of regeneration through collagen-based conduits may involve an initial upregulation of VEGF, which subsequently leads to a corresponding increase in both axonal and SC regeneration. The increase in neovascularization observed within collagen conduits in our experiments is intimately connected with subsequent increases in axonal–SC profile association, possibly through fostering a more conducive environment through which axonal–SC profiles can regenerate.
Long-gap data: Histomorphological analysis
In all cases studied, regenerating cables were filled with myelinated axons, and between 4 and 8 weeks, fibers within both silicone and collagen conduits continued to mature. The mean fiber diameter of normal, uninjured rat sciatic nerve has been reported to be approximately 7.25 ± 0.39 μm, although this parameter has been shown to be reduced dramatically in regenerating sciatic nerves. 58 For example, in studies where entublation strategies using biomaterials such as silicone, collagen, poly (L-lactide-co-ɛ-caprolactone), and Goretex, regenerated mean axon diameters were never greater than 5 μm.59–61 Our results are consistent with previous findings that axons regenerated through silicone conduits have smaller mean fiber diameters than those regenerated through collagen-based conduits.42,60,62 In addition, the mean fiber diameter displayed by the collagen conduit group was very similar to that of the investigations with other collagen-based conduits, 63 and biomaterials such as coil-reinforced hydrogel conduits, 35 which were shown to display results comparable to autograft procedures.
Regenerating cables through collagen conduits were more robust and displayed a significantly greater nerve (whole nerve) area than silicone conduits at both 4 and 8 weeks (Fig. 7D). The pressure cuff theory is one possible explanation for regeneration being mechanically impeded by circumferential contractile forces within our silicone-based group. This, in turn, may have lead to a dramatic decrease in the area of regenerating cables through silicone conduits as compared to those regenerating through collagen conduits. The increase in regenerated cable area of collagen conduits corresponded to a dramatic increase in the total number of myelinated fibers present in these conduits at both 4- and 8-week evaluation points (Fig. 7E), consistent with previous studies examining regenerative differences between silicone- and collagen-based conduits.41,60,62,64
Conclusions
We have studied early regenerative events in a short-gap transection model that are critical for successful peripheral nerve regeneration. The results extend previous findings concerning axon–SC association and suggest that these two intimate processes may be further enhanced by neovascular processes. These associative processes were found to be greater in collagen as opposed to silicone conduits, leading to a greater regenerative response within these conduits. Results from 10-mm nerve injury gap studies mirrored results from our short (5 mm) gap studies. They suggest that collagen-based conduits promote larger regenerating nerve cables, associated with improved vascularity, which in turn provides a more conducive environment for SC migration and axonal regeneration.
Footnotes
Acknowledgments
The authors are grateful to Dr. Qing-Gui Xu and Joanne Forden for technical expertise and assistance.
This research was supported by grants from the Canadian Institute for Health Research and the Alberta Heritage Foundation for Medical Research to R.M. CIHR Regenerative Medicine and Naromedicine Group Grant, RMF-82496.
Disclosure Statement
No competing financial interests exist.