
Abstract
Although bone marrow–derived mesenchymal stromal cells (BMSCs) are a main cell source for tissue-engineered bone (TEB), the clinical use of BMSCs is restricted due to the invasive bone marrow aspiration procedure and the decline in available number of mesenchymal stromal cells (MSCs) and differentiation potential with increasing age. Umbilical cord–derived MSCs (UCMSCs) are likely to be a promising alternative cell source for TEB due to their higher availability and potential to proliferate and differentiate. To assess this possibility, we studied bone morphogenetic protein 2 (BMP2)-induced osteogenic differentiation and activation of signaling pathways in UCMSCs and BMSCs. UCMSCs showed a phenotype and differentiation potential similar to that of BMSCs. After 14 days of BMP2 treatment, the overall expression of several osteogenic-specific phenotypes (type I collagen, osteopontin, and osteocalcin) was similar for UCMSCs and BMSCs. The signaling pathway by which BMP2 induced differentiation of both cell types involved the membrane receptor–initiated signals including SMADs, P38, and extracellular regulated kinase. The similar characteristics of BMP2-induced osteogenic differentiation of UCMSCs and BMSCs in vitro would support the use of UCMSCs in TEB.
Introduction
Human umbilical cord, which is routinely discarded as clinical waste, has been shown to be a promising source for MSCs, which can be isolated from the cord blood, 5 subendothelial layer of cord vein, 6 the connective tissue (Wharton's Jelly),7,8 and perivasculature. 9 Baksh et al. demonstrated that the proliferation and differentiation capacity of MSCs derived from human umbilical cord and human bone marrow is similar. 10 Interestingly, human umbilical cord-derived MSCs (hUCMSCs) showed immunosuppressive effect in vitro and an immunologic phenotype similar to that of BMSCs. 11 Moreover, their major histocompatibility complex class I expression levels can be manipulated. 9 All these features point to the clinical potential of hUCMSCs for use as allogeneic grafts.
Weiss et al. and Medicetty et al. have successfully demonstrated the xenotransplantation of porcine umbilical cord-derived MSCs (UCMSCs) into rat brain. There was neither immune rejection nor teratoma formation for 2–8 weeks following transplantation, and the undifferentiated UCMSCs could transform into neurons or neuron precursors in vivo.12,13 They also showed that undifferentiated and differentiated UCMSCs could promote functional recovery after transplantation into Parkinsonian rats,8,14 which suggested that donor UCMSCs could adapt to the recipient and function normally. Therefore, UCMSCs may be a potential MSC source for stem cell therapies.
BMSCs have been widely used for bone repair through tissue-engineered bone (TEB).5,16 Bone morphogenetic protein 2 (BMP2), a superactive osteogenic inducer, markedly promotes osteogenic differentiation of BMSCs and enhances their ossification quality.17–20 Therefore, BMP2 is used in TEB to accelerate bone healing. We investigated whether BMP2 also promotes the expression of osteogenic phenotypes in hUCMSCs as found in hBMSCs, and assessed the feasibility of using UCMSCs for TEB.
This study focused on the isolation and identification of hUCMSCs and their BMP2-mediated osteogenic phenotypic expression in vitro; the “gold standard” human BMSCs (hBMSCs) were used as a control. The phenotype and multilineage differentiation capacity of UCMSCs were analyzed, and the effect of BMP2 on UCMSC osteogenic differentiation was assessed.
Materials and Methods
Isolation and culture of hUCMSCs and hBMSCs
Human UCMSCs were isolated from umbilical cords from three patients (Supplemental Table S1, available online at www.liebertonline.com) who underwent full-term pregnancy (39–40 weeks) with signed consent forms, according to the methods described by Lu et al. 21 In brief, umbilical cord fragments were incubated with 0.075% collagenase type II (Sigma, St. Louis, MO) for 30 min, followed by 0.125% trypsin (Gibco, Grand Island, NY) for 30 min with gentle agitation at 37°C. The arteries and vein were removed under sterile conditions, and cell suspensions were collected using a 100 μm cell filter. Human BMSCs were isolated by bone marrow aspiration from the iliac crest of three healthy volunteers with signed consent forms according to the approved board protocol (Tissue Engineering Center of Southwest Hospital). Both types of MSCs were cultured in Dulbecco's modified Eagle's medium/Hamm's nutrient mixture F12 (1:1) (DMEM/F12; Hyclone, Logan, UT) with 10% fetal bovine serum (FBS; Hyclone), 100 U/mL of penicillin (Sigma), and 100 μg/mL of streptomycin (Sigma) (Supplemental Fig. S1, available online at www.liebertonline.com). The cells from passages 3–5 were used for the subsequent assays.
Phenotypic analysis
The phenotypes of hUCMSCs and hBMSCs were analyzed by standard flow cytometry techniques. The following antihuman antibodies were used to label the cell surface: CD34-fluorescein isothiocyanate (FITC), CD105-FITC (Abcam, Cambridge, UK), CD44-FITC, CD90-FITC, CD29-phycoerythrin (PE), CD31-PE (BioLegend, San Diego, CA), CD45-FITC, SSEA-4-PE, HLA-DR-PE (eBioscience, San Diego, CA), and CD73-PE (Becton Dickinson Pharmingen, San Diego, CA). The labeled cells were analyzed using a FACSCalibur flow cytometer with CellQuest software (Becton Dickinson). Positive staining was defined as the emission of a fluorescence signal that exceeded levels obtained by more than 98% of cells from the control population stained with matched isotype control antibodies (FITC-conjugated and PE-conjugated monoclonal isotype controls; Becton Dickinson).
Multilineage differentiation assays
Osteogenesis
Cells were cultured in osteogenic medium (DMEM/F12 with 10% FBS, 100 nM dexamethasone, 50 μM ascorbic acid (AsA) 2-phosphate, and 10 mM β-glycerophosphate) (Sigma). On day 21, the cultures were stained for mineralization with alizarin red.
Chondrogenesis
Cells were centrifuged and cultured as pellets in DMEM/F12 with 0.1 μM dexamethasone, 50 μg/mL AsA, 10 ng/mL human recombinant transforming growth factor-beta1 (PeproTech, Rocky Hill, NJ), and 50 mg/mL ITS+ Premix (Becton Dickinson). On day 21, the pellets were prepared for histopathology to detect proteoglycan matrix with safranin O staining.
Adipogenesis
Cells were cultured in DMEM/F12 with 1% FBS, 100 nM dexamethasone, and 1 nM insulin (Sigma). On day 21, cells were stained with oil red O.
RT-PCR
Total RNA was extracted from cells cultured in osteogenic, chondrogenic, and adipogenic media and used for analysis of osteogenic, chondrogenic, and adipogenic gene expression. The reverse transcription (RT) reaction was performed with 1 μg of total RNA, 1 μg of Oligo (dT) primer, and 200 units M-MLV RNase H Minus reverse transcriptase (Promega, Madison, WI) in a total volume of 25 μL. One μL of cDNA was used in the polymerase chain reaction (PCR) with Taq PCR MasterMix (Tiangen, Beijing, China). The primers used are listed in Table 1.
ALP, alkaline phosphatase; RUNX2, runt-related transcription factor 2; ON, osteonectin; COL II, type II collagen; COL X, type X collagen; Agg, aggrecan; LPL, lipoprotein lipase; PPARγ, peroxisome proliferator-activated receptor γ.
Cells cultured with BMP2
Both types of MSCs were plated at a density of 5 × 104/cm2 and cultured in osteogenic medium for following studies. BMP2 blocking antibodies (BMP2 Ab, 5 μg/mL; R&D, Minneapolis, MN), Noggin (50 μg/mL; ProSpec-Tany TechnoGene, Rehevot, Israel), or recombinant human BMP2 (rhBMP2, 50 ng/mL; ProSpec-Tany TechnoGene) were added to the culture. Cells in osteogenic medium without any treatment were used as the control. The agents were mixed into the medium, and the medium was changed everyday.
Assay for ALP activity
The alkaline phosphatase (ALP) activity was assayed by measuring the release of p-nitrophenol from p-nitrophenylphosphate (pNPP; Sigma) according to the methods described by Weber et al. 22 Treated cells were washed twice with phosphate-buffered saline (PBS) and detached with trypsin. The cells were divided into two equal parts. A cell count and total protein assay were conducted on the first part, and the second part was centrifuged and resuspended in 500 μL of pNPP glycine buffer (1 mg/mL; Sigma) for 1 hour at 37°C. The reaction was stopped with the addition of 500 μL of 50 mM sodium hydroxide. The absorbance at 405 nm was measured in a volume of 200 μL using a microplate reader (Bio-Rad, Hercules, CA). The ALP activity was quantified as pg/min/cell/mg protein after calibration with a p-nitrophenol standard curve.
Real-time RT-PCR
Total RNA was extracted from treated cells with Trizol (Invitrogen, Carlsbad, CA). The separation and precipitation of RNA was accomplished with chloroform and isopropyl alcohol. Total RNA was isolated and purified by an RNeasy minikit (Qiagen, Valencia, CA) with the addition of RNase-free DNase I (Qiagen). Purified RNA was quantified with a spectrophotometer, aliquoted, and stored at −20°C.
The two-step SYBR® ExScript™ RT-PCR kit (Perfect Real Time; TaKaRa, Dalian, China) for real-time RT-PCR was used to analyze the expression of the following osteogenic genes: runt-related transcription factor 2 (RUNX2), ALP, osteopontin (OPN), α1 chain of type I collagen (COL I), and osteocalcin (OC). 18S RNA was used as the housekeeping gene. Quantification was achieved using standard curves derived from gene expression relative to the level of 18S RNA gene expression. The quantification of the BMP2 gene in treated cells used the same method. The primers used are listed in Table 2 as described by Hou et al. 23
OPN, osteopontin; COL I, α1 chain of type I collagen; OC, osteocalcin; BMP2, bone morphogenetic protein 2.
Western blot analysis
The cells were lysed by incubation at 4°C for 30 min in buffer containing RIPA (0.1 M PBS = 0.1 mol/L PBS. PBS, 1% NP40, 0.1% sodium dodecyl sulfate, 5 mM EDTA, 0.5% sodium deoxycholate, 1 mM sodium orthovanadate, 1% PMSF). The samples were fractioned on an 8%–15% SDS-PAGE gel (8% for COL I; 12% for all cell signaling pathway proteins, RUNX2, BMP2, and β-actin; and 15% for OC) and transferred to a nitrocellulose membrane. The primary antibodies for all cell signaling pathway proteins (anti-phospho-SMAD1/5/8, anti-phospho-p38, anti-p38, anti-phospho-SAPK/c-JUN N-terminal kinase [JNK], anti-SAPK/JNK, anti-phospho-p42/44 extracellular regulated kinase (ERK), and anti-p42/44 ERK from Cell Signaling Technologies, Danvers, MA; anti-SMAD1/5 from Upstate Biotechnology, Charlottesville, NC) were used at a 1:500 dilution; β-actin antibody (Abcam) was used at a 1:5000 dilution; and RUNX2, COL I (Santa Cruz, Santa Cruz, CA), BMP2, and OC (Abcam) antibodies were used at a 1:1000 dilution. Horseradish peroxidase-conjugated secondary antibody (SouthernBiotech, Birmingham, AL) was used at a 1:2000 dilution. Immunoreactivity was detected by enhanced chemiluminescence (KPL, Gaithersburg, MD).
Immunofluorescent analysis
The cells were fixed in 4% paraformaldehyde and permeabilized in PBS containing 0.1% Triton X-100 and 5% bovine serum albumin. The cells were subsequently incubated overnight at 4°C with primary antibodies against phosphorylated proteins and osteogenic proteins. The cells were then incubated with Alexa Fluor 594 (Invitrogen) or FITC (SouthernBiotech)-labeled secondary antibodies and were counterstained with DAPI (Sigma) or phalloidine-FITC (Sigma).
Statistical analysis
All experiments were repeated in triplicate and results reported as the mean ± SD based on independent samples. For statistical analysis, real-time RT-PCR data and western blot data were transformed to natural log values to obtain normally distributed data. The paired t-test was used to compare data between UCMSCs and BMSCs with the SPSS version 10.0 software package. p-Values less than 0.05 were considered to indicate significance.
Results
Phenotypic profile
Flow cytometric analyses of passage 3 hUCMSCs showed a nonhematopoietic CD45−and CD34−cell population similar to hBMSCs. The cells derived from both sources were also negative for matrix receptor CD31 (PECAM-1), HLA-DR, and SSEA-4. However, the percentage of positive expression in hUCMSCs was lower than in hBMSCs. Both cell types were labeled positive for CD29, CD44, CD73, CD90, and CD105, and exhibited a fibroblast-like morphology (Supplemental Fig. S2, available online at www.liebertonline.com).
Multilineage differentiation capacity
The differentiation potential of hUCMSCs and hBMSCs was tested through culture in multilineage differentiation conditions. Alizarin red staining showed the formation of mineralization in osteogenic cultures from both sources on day 21. Chondrogenesis and adipogenesis were identified by safranin O staining and oil red O staining, respectively. From both cell sources, a number of proteoglycans were formed in chondrogenic cultures, and lipid droplets were also found in adipogenic cultures (Fig. 1A).
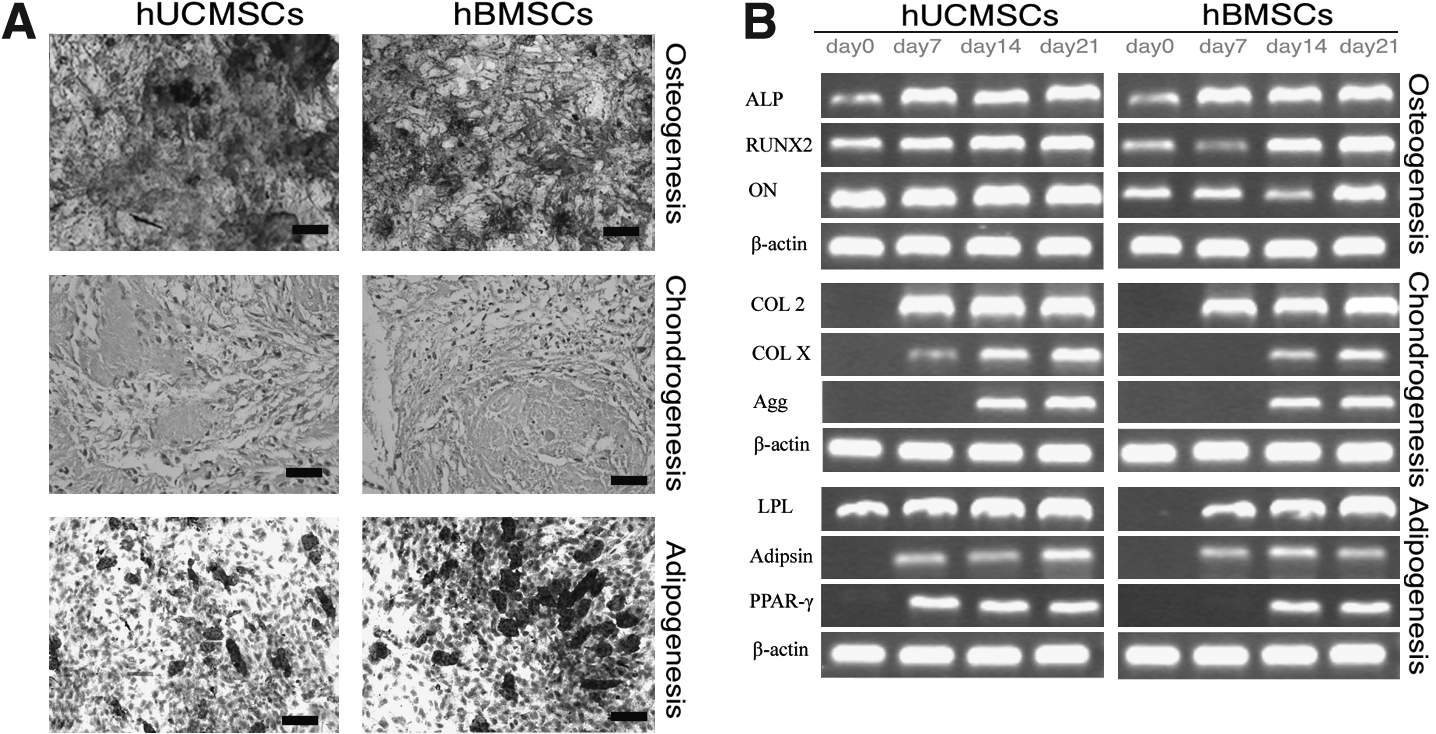
Multilineage differentiation capacity of hUCMSCs and hBMSCs was confirmed by histochemical staining (
The osteogenic phenotype was also shown by the expression of the osteogenic gene osteonectin mRNA, and by the gradual increase in ALP and RUNX2 mRNA expression (Fig. 1B). The mRNA of chondrocytic markers, including type II and type X collagen (COL II and COL X) and aggrecan, were detected by RT-PCR after 7 and 14 days of chondrogenic induction. The mRNA of adipocytic markers, including adipsin, peroxisome proliferator-activated receptor γ, and lipoprotein lipase, were found by RT-PCR after 7 and 14 days of adipogenic induction (Fig. 1B).
Increase of ALP activity under induction of BMP2
The level of ALP activity in cultured hUCMSCs and hBMSCs remained constant (Fig. 2A). However, addition of rhBMP2 significantly increased the ALP activity from day 8 (p < 0.05). The increase of ALP activity in hUCMSCs was similar to that in hBMSCs (8- and 7-fold, respectively). In contrast, the addition of BMP2 Ab and Noggin slightly decreased the ALP activity to about 60% of baseline.

Expression of osteogenic differentiation markers in bone morphogenic protein 2 (BMP2)-stimulated hUCMSCs and hBMSCs. The cells were incubated with BMP2 (50 ng/mL), BMP2 Ab (5 μg/mL), and Noggin (50 μg/mL). The cells in osteogenic medium without any treatment were used as control. After 14-day incubation, the treated cells were assayed for osteogenic differentiation markers by enzymatic assay (
Expression of osteogenic phenotypes under induction of BMP2
The expression of osteogenic phenotypes of differentiated hUCMSCs and hBMSCs was detected by real-time RT-PCR, western blot, and immunofluorescence. The quantification of gene expression by real-time RT-PCR showed that in control group the expression of all osteogenic genes in hUCMSCs was lower than in hBMSCs, and these differences reached significance in ALP gene and COL I gene (Fig. 2B, D). However, there was no significant difference in other groups (BMP2 Ab, Noggin, and BMP2). As for each type of cell (hUCMSCs or hBMSCs), the addition of rhBMP2 markedly increased the mRNA levels of several osteogenic genes including ALP, RUNX2, COL I, OPN, and OC compared to the controls (Fig. 2). In contrast, their expression levels were slightly decreased by treatment with BMP2 Ab and Noggin. After 14-day BMP2 treatment, the mRNA level of ALP gene in differentiated hUCMSCs and hBMSCs increased approximately 5-fold and 3-fold, respectively, compared with the control (Fig. 2B). Similarly, the mRNA levels of COL1, OPN, RUNX2, and OC in differentiated hUCMSCs increased 16.8-fold, 11.5-fold, 7.4-fold, and 30.8-fold, respectively, in response to BMP2 treatment. In differentiated hBMSCs, the values were 6.8-fold, 5.1-fold, 4.8-fold, and 15.5-fold, respectively (Fig. 2C–F). Thus, the expression of osteogenic phenotypes in hUCMSCs and hBMSCs was dramatically increased by BMP2 treatment.
The results of western blot (Fig. 3A–D) and immunofluorescence (Fig. 3E) were similar to real-time RT-PCR. The expression levels of RUNX2, COL I, and OC markedly increased compared with the control after 14-day BMP2 treatment. In contrast, their expression levels were slightly decreased by BMP2 Ab and Noggin. The difference in osteogenic protein expressions between hUCMSCs and hBMSCs in all four groups was not significant except for RUNX2 protein in the control, BMP2 Ab, and BMP2 groups, and for OC protein in Noggin group (Fig. 3B, D, p < 0.05).
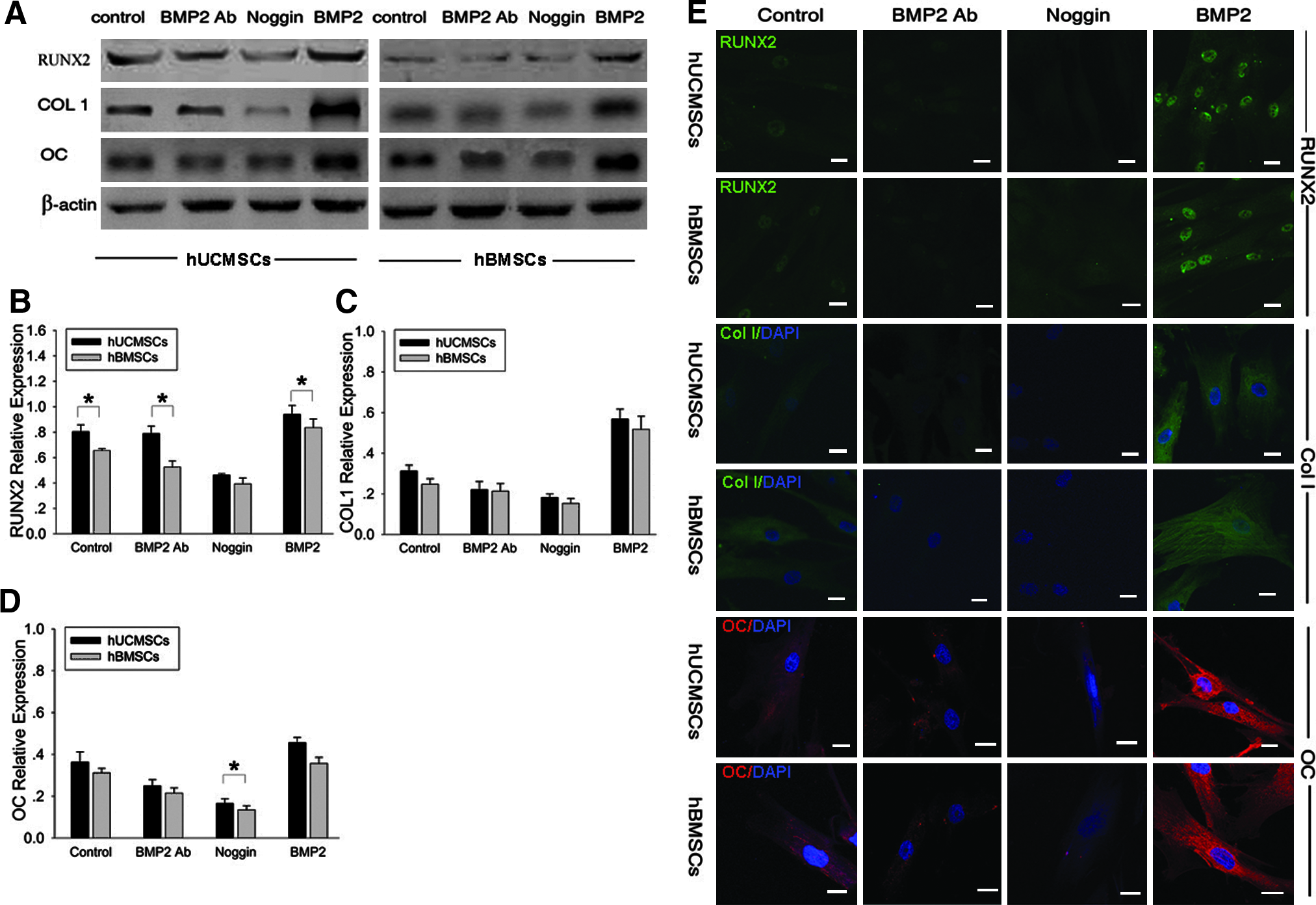
Expression of osteogenic differentiation markers in hUCMSCs and hBMSCs under induction of BMP2 was detected by western blot (
Activation of BMP signal transduction pathways
To investigate the signal transduction pathway by which BMP2 stimulated the expression of osteogenic phenotypes in hUCMSCs and hBMSCs, we examined the activation of downstream factors, SMADs, P38, JNK, and ERK, by western blot. Both types of MSCs were pretreated with or without BMP2 Ab or Noggin for 30 min followed by incubation with rhBMP2 for 1 hour. The cells were lysed and subjected to western blot. The amount of phosphorylated SMAD1/5/8 in both types of MSCs was clearly increased by rhBMP2 treatment compared to the control, while the pretreatment with Noggin dramatically decreased the rhBMP2 effect in both MSCs. The expression of total SMAD was not affected by treatment with rhBMP2, BMP2 Ab, or Noggin. Similar to SMAD, the expression of total and phospho-P38 was changed by treatment with rhBMP2, BMP2 Ab, and Noggin, except that Noggin pretreatment reduced the level of total P38 in hUCMSCs. The total and phospho-JNK levels were not affected by treatment with rhBMP2, BMP2 Ab, or Noggin in either of the MSCs, except that phospho-JNK was inhibited by BMP2 Ab and Noggin in hUCMSCs. However, the phospho-ERK level in hUCMSCs was markedly increased by rhBMP2 induction. The phospho-ERK level was inhibited by BMP2 Ab as well as Noggin in both MSCs, and especially in hUCMSCs through Noggin pretreatment. On the other hand, rhBMP2 failed to activate ERK in hBMSCs (Fig. 4).
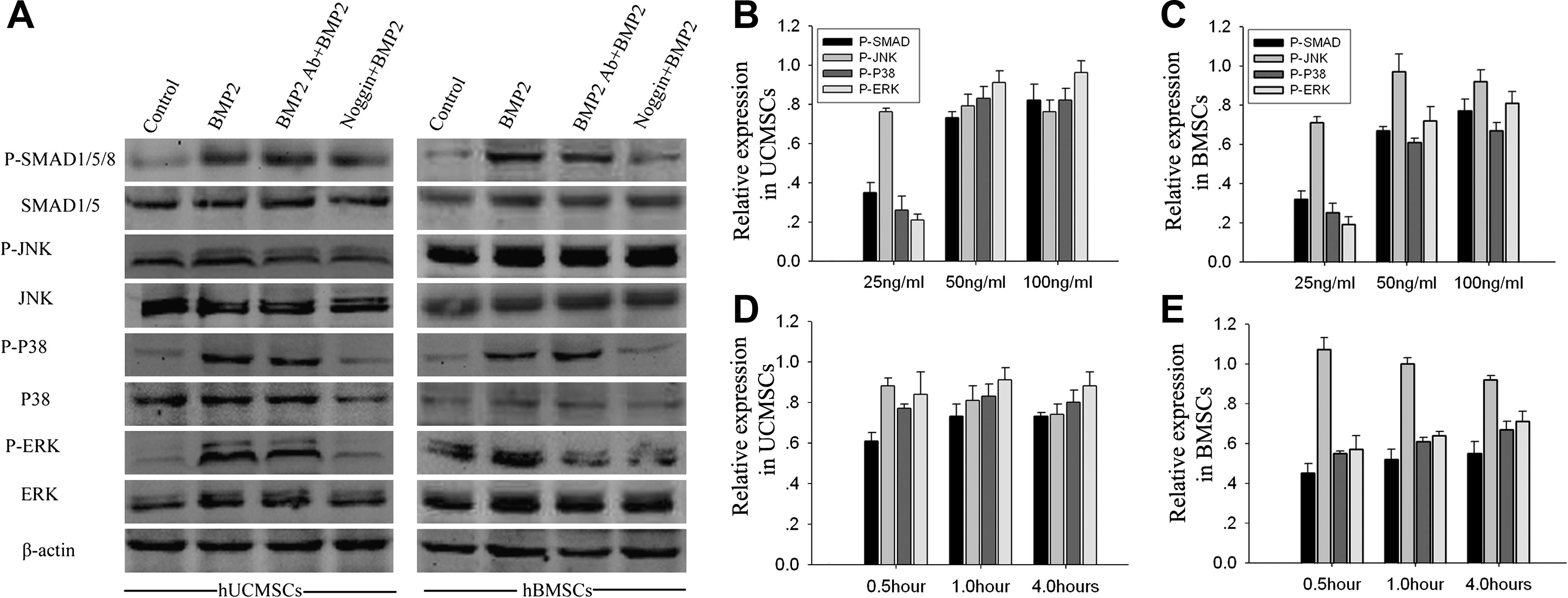
BMP2-responsive signal transduction pathways in hUCMSCs and hBMSCs were assayed by western blot. The cells were pretreated with BMP2 Ab (5 μg/mL) and Noggin (50 μg/mL) for 30 min, followed by treatment with rhBMP2 (50 ng/mL) for 1 hour before harvesting. Control was defined as cells in osteogenic medium without any treatment (
The time-dependent and BMP2-dose-dependent changes of signaling pathway (phosphorylated proteins) were also studied. The profile of signaling pathway activated by different dosages of BMP2 (25, 50, and 100 ng/mL) in both types of MSCs was similar. With increased concentrations of BMP2, relative expression of phospho-JNK remained unchanged. However, the relative expression of phospho-SMAD1/5/8, phospho-P38, and phospho-ERK increased. Incubation with BMP2 for 0.5 hour and 1 hour slightly increased phospho-SMAD1/5/8, phospho-P38, and phospho-ERK in both UCMSCs and BMSCs. However, no significant differences were found by extended incubation (from 1 hour to 4 hours).
The immunofluorescence of phosphorylated proteins showed changes similar to those of western blot following treatment with rhBMP2, BMP2 Ab, or Noggin (Fig. 5). The phospho-SMAD and phospho-P38 in both MSCs appeared aggregated in cell nucleus following rhBMP2 treatment. Although phospho-ERK mostly localized in cell nucleus, a small portion was also found in the cytoplasm. In contrast, phospho-JNK was distributed only in the cytoplasm.
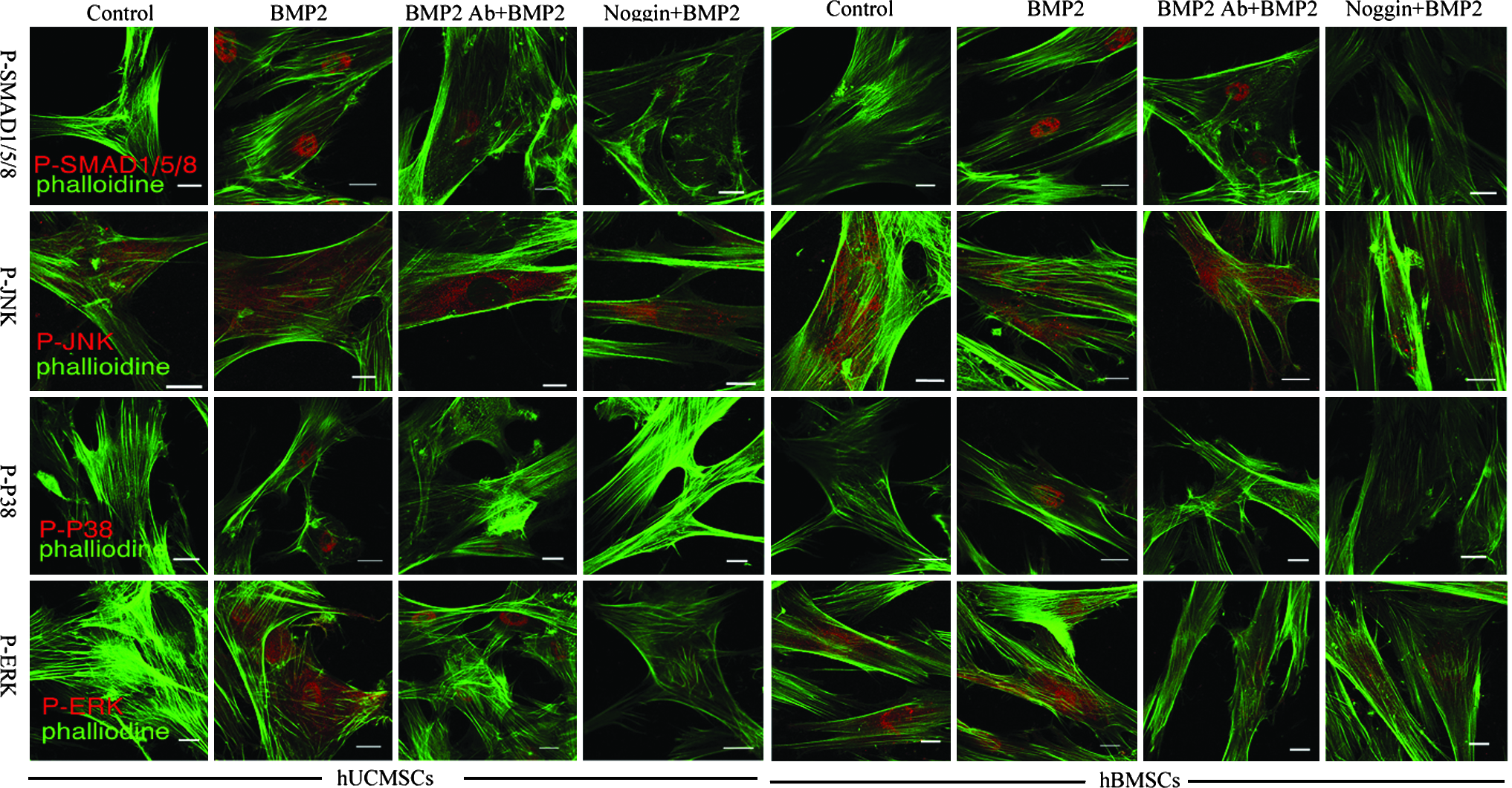
Immunofluorescent staining showed the local expression of BMP2-responsive signal transduction proteins in hUCMSCs and hBMSCs. The cells were pretreated with BMP2 Ab (5 μg/mL) and Noggin (50 μg/mL) for 30 min, followed by treatment with rhBMP2 (50 ng/mL) for 1 hour before harvesting. Control was defined as cells in osteogenic medium without any treatment. The cells were fixed and used for further staining. Scale bar: 20 μm.
Endogenous BMP2 synthesis
MSCs are known to produce BMP2, which stimulates their differentiation in an autocrine manner. The effect of additional stimulators (rhBMP2, BMP2 Ab, and Noggin) on the endogenous synthesis of BMP2 was analyzed by real-time RT-PCR and western blot. After 6-day treatment, the difference in BMP2 gene expression between hUCMSCs and hBMSCs in all four groups was not significant. The results also showed that rhBMP2 increased the mRNA level of BMP2 by 23% in hUCMSCs and by 40% in hBMSCs; treatment with BMP2 Ab decreased the mRNA level of BMP2 to 35% and 39% of baseline, respectively. Similarly, Noggin decreased the mRNA level of BMP2 in hUCMSCs and hBMSCs to 27% and 25% of baseline, respectively (Fig. 6A). Similar results were obtained by western blot (Fig. 6B).
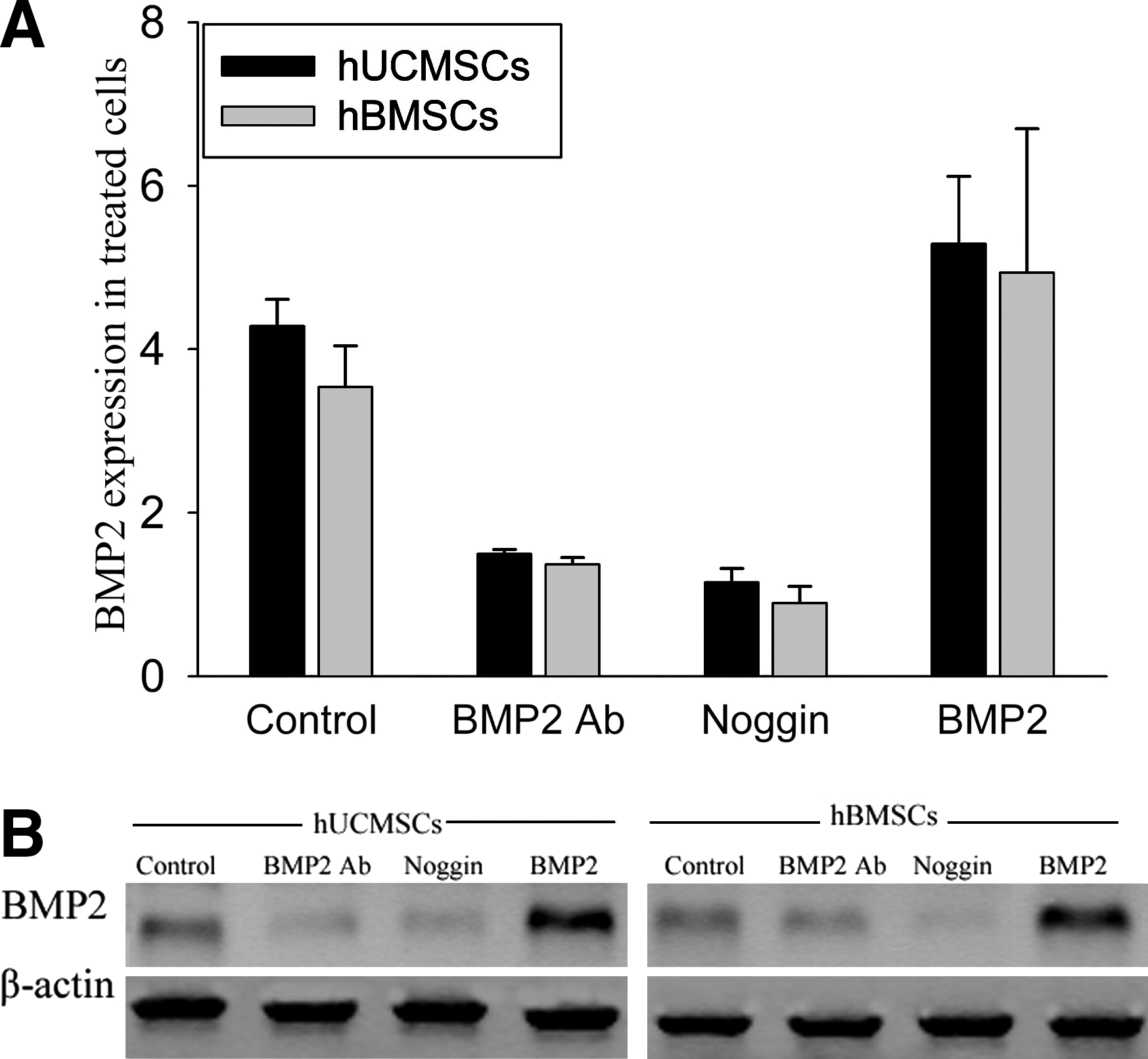
The endogenous synthesis of BMP2 in hUCMSCs and hBMSCs. The cells were incubated with rhBMP2 (50ng/mL), BMP2 Ab (5 μg/mL), and Noggin (50 μg/mL). Control was defined as cells in osteogenic medium without any treatment. After 6-day incubation, the cells were collected to assay BMP2 mRNA by real-time RT-PCR (
Discussion
Although BMSCs have been established as a cell source for bone repair and reconstruction, their clinical application is restricted by the invasive bone marrow harvest procedures as well as the decrease in proliferation and differentiation capabilities with increasing age. UCMSCs isolated from discarded umbilical cords are an inexhaustible alternative cell source for bone repair and reconstruction because of the convenient and noninvasive harvesting procedure. UCMSCs also possess higher proliferation and osteogenic capabilities than BMSCs. 10 BMP2 is a superactive osteogenic inducer that can promote osteogenic differentiation in many types of cells such as BMSCs, fibroblasts, and myoblasts.17,24,25 During bone repair and reconstruction, administration of BMP2 promotes bone healing and is beneficial to bone reconstruction.18–20 Based on these observations, we examined whether BMP2 could induce expression of osteogenic phenotypes in hUCMSCs, since BMP2-treated hUCMSCs could be an alternative source for bone repair and reconstruction.
We first characterized MSCs from bone marrow and umbilical cord by assessing the fibroblast-like morphology, the expression of a classical set of surface proteins, and the multipotential differentiation capability according to the criteria described by the International Society for Cellular Therapy. 26 The results showed that both hBMSCs and hUCMSCs in this study possessed the above three characteristics of MSCs, which was consistent with previous research. 10
The researches on hUCMSCs have significantly increased in recent years. The methods of isolation and culture of hUCMSCs used by the different investigators are similar, which involve removal of vessels and isolation-digestion with collagenase. Many different studies have demonstrated that hUCMSCs possess osteogenic potential.7,9,10,27,28,29 Further, Baksh et al. 10 showed that perivascular cells generate a greater extent of mineralization than BMSCs.
During osteogenic differentiation under the effect of BMP2, some osteogenic markers would express one after another. ALP is an early marker of osteogenic differentiation and mineralization, and the increase in ALP level determines potential osteogenic differentiation. 30 RUNX2, the osteoblast-specific product of the Cbfa1 gene, is a runt domain-containing transcription factor that is essential for osteogenic differentiation and bone formation. Like ALP, expression of RUNX2 is increased in the early stage of osteogenic differentiation. RUNX2 overexpression in BMSCs can induce osteoblast-specific gene expression and mineralization.31,32 The activated RUNX2 triggers downstream molecules including OPN, COL1, and OC, which are also osteoblast-specific markers. OPN, one of the major noncollagenous proteins in extracellular bone matrix, is synthesized by osteoblasts and osteocytes and its expression is initiated in the later differentiation stage of the cell.33,34 COL1 is the abundant form of collagen present in bone matrix and is also synthesized by osteoblasts. 30 OC, a late marker for osteogenesis, plays a regulatory role in balancing the maintenance (resorption) of mineralized bone. 35 In the present study, these relative osteogenic differentiation markers were detected when analyzing the effect of BMP2 on hUCMSCs and hBMSCs by ALP activity, real-time RT-PCR, western blot, and immunofluorescent staining. The antagonist of BMP2, BMP2 Ab, and Noggin36,37 could dramatically decrease the expressive levels of relative osteogenic differentiation markers compared with the control in both cell types. These results confirmed that BMP2 initiated the osteogenic differentiation of hUCMSCs and hBMSCs, which was consistent with previous studies.22,37 Real-time RT-PCR and western blot analyses showed that expression of these osteoblast-specific markers in BMP2-stimulated hUCMSCs was comparable to that in hBMSCs, suggesting that BMP2 was as effective an osteogenic inducer for hUCMSCs as for hBMSCs.
The endogenous BMP2 also played a role in the process of osteogenic differentiation mediated by extracellular rhBMP2. 17 Further, BMP2 Ab and Noggin decreased ALP activity and expression of other osteogenic-specific genes (ALP, RUNX2, OPN, COL1, and OC) in hUCMSCs and hBMSCs compared to the control. Thus, endogenous BMP2 plays a role during the osteogenic differentiation of hUCMSCs, as with BMSCs. 17 However, the antagonistic effect of Noggin on osteogenesis in hUCMSCs appears more effective than that of BMP2 Ab, probably because Noggin noncompetitively inhibits BMP2, 4 and 7 38 while BMP2 Ab is specific for BMP2.
We then investigated the BMP signal transduction pathway in both hUCMSCs and BMSCs. BMP signals are mediated by type I and type II serine/threonine kinase receptors. When the ligand binds, a heterodimer is formed by the type I and II receptors. The constitutive kinase of the type II receptor activates the type I receptor and initiates the signal transduction cascade by phosphorylating downstream factors (SMADs), which subsequently translocate to the nucleus to regulate the transcription of target genes with the help of RUNX2. 39 BMP2 also activates the mitogen activated protein kinase (MAPK), ERK1/2, JNK, and P38. MAPKs can regulate independent downstream effects and can also phosphorylate SMADs.40–42 We found that BMP2 dramatically increased the level of phosphorylated SMADs1/5/8, P38, and ERK but not JNK in hUCMSCs. In hBMSCs, BMP2 significantly raised the level of phosphorylated SMADs1/5/8 and P38 and slightly increased the level of phosphorylated ERK but not JNK, in agreement with previous studies. 37 Both MSCs showed similar phosphorylation profiles in response to BMP2 stimulation in a dose-dependent and time-dependent manner, suggesting that BMP2 stimulates differentiation of these two types of MSCs via similar pathways.
Based on these results, we concluded that both hUCMSCs and hBMSCs possess the three characteristics of MSCs discussed above. Further, BMP2 could promote expression of some special osteogenic markers for hUCMSCs via a pathway similar to that for hBMSCs. The quantitative analysis through real-time RT-PCR and western blot showed that expression levels of special osteogenic markers in hUCMSCs were close to and even more than that in hBMSCs. This raises the question, how cells from two different sources can display such similar characteristics? Both cell types are also found to possess similar characteristics without BMP2 treatment.10,43 Some authors consider that hUCMSCs are earlier-stage cells than hBMSCs,7,44 and the early embryonic antigen SSEA-4 was reported to identify an adult MSC population. 45 We found that the percentage of SSEA-4-positive cells was lower in hUCMSCs. However, the detail mechanism is unclear yet and needs further investigation.
To summarize, BMP2 successfully mediates the osteogenic differentiation of hUCMSCs through a signal transduction pathway similar to that for hBMSCs. The expression level of osteogenic phenotypes in hUCMSCs under the effect of BMP2 is comparable to that in hBMSCs. In addition, hUCMSCs in Wharton's Jelly of the umbilical cord are more advantageous than hBMSCs, with higher proliferation potential, 10 larger potential donor pool, and noninvasive harvest. Thus, considering the feasibility, MSCs derived from Wharton's Jelly of the umbilical cord are likely to be a promising alternative source for bone tissue engineering. However, the results of this study need to be further verified in vivo.
Footnotes
Acknowledgments
We gratefully acknowledge Dr. Caixia Li's guidance on isolation of cells. This work was supported by the National High Technology Research and Development (863) Program of China (2006AA02A122), Chongqing Key Scientific and Technologic Program (2006AB5008), and the Military Foundation.
Disclosure Statement
No competing financial interests exist.