
Abstract
The capacity of human mesenchymal stem cells (hMSC) for self-renewal and differentiation is a tightly regulated process within their microenvironment—the stem cell niche. For future therapeutic applications of hMSC within the frame of tissue engineering, it is of major importance to understand the factors involved in triggering differentiation cascades of hMSC. Using either osteoblast-conditioned medium or an indirect coculture system, we investigated whether soluble factors from human osteoblasts (hOB) are sufficient to induce early osteogenic markers in hMSC. Thereby, we detected an induction of several osteogenic markers like alkaline phosphatase, bone sialoprotein 2, leptin receptor, decorin, and cathepsin K in hMSC as indicators of the onset of early osteogenesis. Further, because Wnt signaling has been reported to play an important role in osteogenesis, we performed RNAi against the main Wnt mediator β-catenin and the low-density lipoprotein receptor-related protein 5 as a major Wnt co-receptor in hMSC. Whereas alkaline phosphatase was significantly downregulated with this approach, the other osteogenic markers showed a markedly upregulation. These observations suggest that hOB-secreted factors could induce early osteogenic markers in hMSC. Thus, with regard to a therapeutic setting, these findings may pave the way for a more in vivo–related differentiation procedure for the generation of osteoblast-like cells.
Introduction
Osteoblasts are derived from mesenchymal stem cells (MSC) and are located close to the bone marrow, which serves as a major source for MSC and hematopoietic stem cells.7–9 Against this background, it has been demonstrated that human mesenchymal stem cells (hMSC) are pluripotent bone marrow cells that can be expanded ex vivo and differentiated into several mesodermal tissues like cartilage, fat, and bone. 10 The capacity for self-renewal and differentiation represents basic characteristics of stem cells that are under fine-tuned control in the stem cell niche by neighboring cells. 11 In this regard, it was shown that osteoblasts are involved in the regulation of stem cell properties within the hematopoietic stem cells niche. 12 In the case of epithelial stem cells, the hair bulge functions as a niche,13,14 where several inhibitory components of the Wnt signaling pathway, such as Dickkopf (Dkk), secreted frizzled-related proteins, and Wnt inhibitory factor, are expressed, leading to the assumption that this area provides a growth- and differentiation-restricted environment. 15
On the other hand, it has been demonstrated that Wnt signaling is activated during many embryonic developmental stages, 16 in numerous stem cell compartments 17 and in different forms of cancer. 18 Further, Wnt signaling is involved in regulating basic stem cell features such as self-renewal 19 and migration/invasion of hMSC. 20 In general, Wnts are secreted glycoproteins that can initiate the canonical Wnt pathway when binding to a specific heterodimeric receptor implying a member of the Frizzled receptor (Fzd) family and their LRP co-receptors.21,22 In the absence of canonical Wnts, cytoplasmic β-catenin as a main Wnt signaling mediator is phosphorylated by the destruction complex composed of axin, adenomatous polyposis coli protein (APC), and glycogen synthase kinase 3β (GSK-3β). With ubiquitination, β-catenin gets marked for degradation by the proteasome. In the presence of canonical Wnt ligands, the axin/APC/GSK-3β complex is inactivated, resulting in the accumulation of β-catenin in the cytoplasm with subsequent translocation into the nucleus. As a consequence, β-catenin forms a complex with T-cell-specific transcription factor/lymphoid enhancer binding factor (TCF/LEF) family proteins and acts as a bipartite transcription factor that binds to the promoter of Wnt responsive genes, thereby driving their transcription. 23 Despite the importance of this pathway in the regulation of basic stem cell features, detailed knowledge about the involvement of Wnt signaling in triggering osteogenic differentiation processes is lacking. Rather, controversial perspectives have been reported.19,24
Based on our working hypothesis that similar to the setting in the stem cell niche the differentiation niche in the target tissue may participate in the regulation of the hMSC phenotype, we evaluated whether secreted factors of human osteoblasts (hOB) are sufficient to initiate early osteogenic markers in hMSC. Further, the impact of canonical Wnt signaling was analyzed in this context.
Materials and Methods
Cell culture
Cryopreserved hMSC were purchased from Lonza, Basel, Switzerland. Commercially available hOB (PromoCell, Heidelberg, Germany) were used for the generation of conditioned medium and for indirect cocultivation experiments. The cultivation of hMSC was performed in MSC growth medium (Lonza), as recently described, 25 or in osteoblast growth medium with regard to the conditioned medium or indirect coculture approaches. hOB were cultivated in osteoblast growth medium, which had been supplemented by a supplementary mix provided by the manufacturer (PromoCell). In our experiments hMSC and hOB from two different donors were evaluated (hMSC I and hMSC II, and hOB I and hOB II). hMSC were used at the fifth or sixth passage of cultivation, and hOB at the third or fourth passage. For the induction of osteogenic differentiation, 50 μM L-ascorbic acid 2-phosphate, 100 nM dexamethasone, and 10 mM β-glycerophosphate (all from Sigma, St. Louis, MO) were applied as previously described. 25
Alizarin-red staining
To visualize calcium deposition upon osteogenic differentiation, an Alizarin-red-S (Sigma) stock solution of 40 mM was used. Therefore, cells were washed three times with PBS and fixed in 2 mL of ice-cold 70% ethanol for 1 h. After fixation, cells were rinsed with H2O and stained for 10 min in 1 mL Alizarin-red-S stock solution under continuous rotation. To diminish unspecific staining, cells were washed three times in PBS. Stained samples were stored at 4°C covered with PBS.
hOB-Conditioned supernatant
To guarantee standardized conditions for the generation of hOB-conditioned medium, refreshment of osteoblast growth medium purchased from PromoCell was performed after hOB had reached 80% confluence. To achieve an accumulation of hOB-derived secreted factors, hOB cultivation was maintained for 5 days in osteoblast growth medium. This hOB-conditioned supernatant (cOBS) was sterile-filtrated using a 0.2 μm filter (Millipore/Nalgene, Billerica, MA) before further experiments were carried out. cOBS was changed twice a week in each experimental setting.
Indirect coculture assay
For performing indirect coculture assays a transwell system was used, consisting of an upper and a lower compartment. To avoid mixing, 24 h before the start of indirect coculture experiments, 64,000 hMSC were seeded on polycarbonate membrane filters (upper compartment, 24 mm diameter and 0.4 μm pore size) placed in a set of blank tissue culture plates, while hOB were seeded on the bottom of Costar Transwell® chambers (Costar, Corning, NY). To monitor the influence of hOB on the differentiation behavior of hMSC, two different numbers of hOB (64,000 vs. 128,000) were applied to the lower compartment, whereas hMSC numbers were kept constant. Medium was changed twice a week using a total volume of 3.5 mL osteoblast growth medium. On days 4, 7, and 14 hMSC were harvested in 350 μL RLT buffer (Qiagen, Hilden, Germany) for RNA isolation and subsequent quantitative RT-PCR analysis.
RNA isolation and quantitative reverse transcriptase–polymerase chain reaction
DNase-digested total RNA was extracted from hMSC using the RNeasy Mini Kit (Qiagen) according to the manufacturer's protocol. cDNA synthesis was performed from 100 to 500 ng total RNA using iScript cDNA Synthesis Kit (BioRad, Hercules, CA) according to the instructions of the supplier. Quantitative reverse transcriptase–polymerase chain reaction (RT-PCR) was carried out on a LightCycler (Roche, Mannheim, Germany) using LightCycler-FastStart DNA Master SYBR Green I Kit (Roche). For the quantification of alkaline phosphatase (ALP), bone sialoprotein 2 (BSP-2), cathepsin K (CTSK), decorin (DCN), GAPDH, leptin receptor (LR), osteocalcin (OCN), and Runx-2 transcripts, primer sets were designed and purchased from Search LC (Heidelberg, Germany). PCR was performed with 1 μL cDNA as a template, 2 μL primer set (Search LC), 2 μL LC-FastStart DNA Master SYBR Green I mix, and 15 μL H2O. Thermocycling was accomplished as recently described. 25
Microarray
To obtain deeper insights into the osteogenic differentiation process, transcriptional profiles of hOB and hMSC were analyzed using Human Osteogenesis Oligo GEArray® Microarrays (Superarray, Frederick, MD). This microarray contains 128 spots: 114 osteogenic genes, 12 controls (housekeeping genes and artificial sequences), and 2 blank spots (for detailed information: www.superarray.com). Transcriptional changes of hMSC after treatment with either osteogenic differentiation medium (ODM) supplemented with 50 μM L-ascorbic acid 2-phosphate, 100 nM dexamethasone, and 10 mM β-glycerophosphate or cOBS were assayed and compared to the expression levels of hMSC and hOB cultivated in their regular growth medium. After RNA isolation according to the supplier's protocol (Superarray), RNA denaturation was carried out at 70°C, and 3 μg of the respective RNA samples was transcribed into cDNA. Biotin-labeled cRNA was synthesized and amplified during incubation over night. For microarray hybridization, 6 μg of the amplified cRNA was applied to the hybridization solution, and GEArray membranes were hybridized over night at 60°C. Spots were observed using a chemiluminescence reaction (Superarray). Mircroarray data were quantified using ArrayVision 6.0 (Imaging Research, St. Catharines, Canada).
Small interfering RNA transfection of hMSC
Small interfering RNAs (siRNAs) targeted against β-catenin or LRP5 were designed in our laboratory 20 according to the protocol developed by Reynolds et al. 26 One day before siRNA transfection, 48,000 hMSC were transferred into a cavity of a six-well plate by adding 2 mL hMSC medium, resulting in 30% confluence after 24 h incubation. For preparing the transfection mixtures, the respective siRNAs were added to 250 μL serum-free Dulbecco's modified Eagle's medium in a final concentration of 25 nM. In a separate tube, 5 μL of Lipofectamine 2000 (Invitrogen, Carlsbad, CA) was diluted in 250 μL serum-free Dulbecco's modified Eagle's medium. After unifying both solutions, the final transfection mixture was incubated for 20 min at room temperature. This transfection solution was applied to the cells, and after an incubation period of 6 h replaced by 2 mL hMSC medium. To obtain a high cell density that is indispensable for differentiation assays, hMSC were trypsinized in the 6-well plate 1 day after transfection and seeded into a 12-well plate. One day later, medium was changed to either osteoblast growth medium or cOBS, and further cultivation was performed for 7 days.
Statistical analysis
Statistical analysis was performed using SigmaPlot 10.0 (Systat Software, Richmond, CA). Statistical significance was assessed by comparing mean values (±SD) with Student's t-test for independent groups. Significance was assessed for p < 0.05 and p < 0.01.
Results
Alizarin-red staining of hOB
To determine whether hOB used in this study are more preosteoblast-like or osteocyte-like, seven different combinations of supplements were added to the differentiation media. For that purpose, we used 50 μM L-ascorbic acid 2-phosphate (Asc), 100 nM dexamethasone (Dex), and 10 mM β-glycerolphosphate (Gly) as single additives as well as combinations of two or three components. hOB cultivated without any supplements served as a control. After a differentiation period of 14 days, Alizarin-red staining was carried out. For positive staining results reflected by calcium deposition, the addition of dexamethasone and β-glycerol-phosphate was an essential part (Fig. 1A). Additional use of L-ascorbic-2-acid showed the most prominent staining results as multiple and distinctive spots of Alizarin-red stains could be observed (Fig. 1B). All of the remaining combinations resulted in negative stainings (Fig. 1C–H). Thus, our data indicate that hOB used in this study exhibit a preosteoblast phenotype, and for this reason dexamethasone and β-glycerophosphate are essential additives to deposit calcium phosphate complexes in their surrounding matrix.
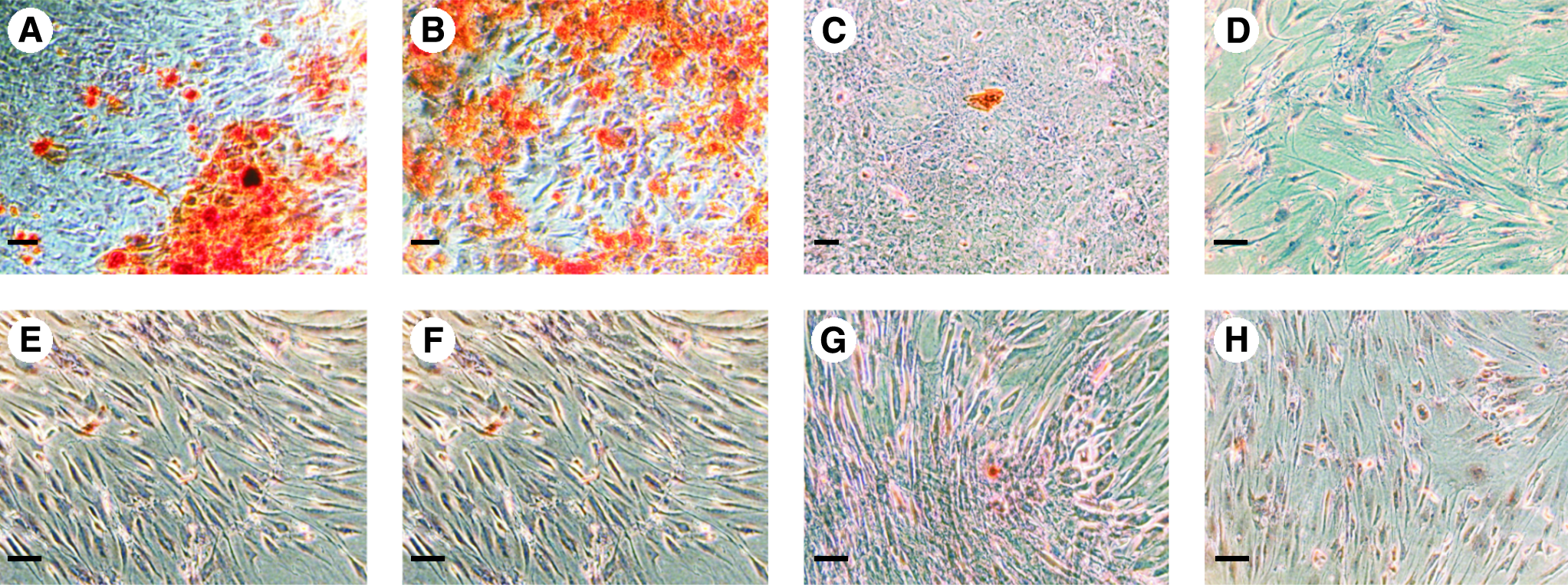
Alizarin-red-S staining of human osteoblasts (hOB) stimulated with different combinations of osteogenic supplements. (
Upregulation of osteogenic markers in hMSC by cOBS
To analyze the effects of cOBS on hMSC, mRNA expression levels of the osteogenic markers ALP, BSP-2, OCN, the receptor for the obesity hormone leptin LR, the cysteine proteinase CTSK, the osteogenic matrix protein DCN, and the osteogenic TCF Runx-2 were quantified by RT-PCR. hMSC cultivated in cOBS were analyzed on days 1, 4, 7, and 14. Normalization on the house keeping gene GAPDH was performed using the respective GAPDH values. Whereas the ALP expression of hMSC stimulated with cOBS was not affected after 24 h incubation, the ALP transcript levels were significantly increased from day 4 to 14 (Fig. 2A). As indicated in Figure 2B, LR expression of cOBS-stimulated hMSC presented increased values on days 4 and 7 (3.1-fold and 2.4-fold) when compared to their respective nonstimulated control samples. Initially, ALP and LR levels were low, but exhibited a slight increase from day 1 to 14 considering control samples only. Likewise, DCN expression levels increased over time. Control cells showed significantly lower DCN copy numbers on day 4 (1.8-fold) and day 7 (1.7-fold) in comparison to cOBS-treated hMSC, whereas on day 14 no significant difference could be detected (Fig. 2C). In contrast, the cysteine proteinase CTSK mRNA expression was significantly increased within the first 4 days, but exhibited a continuous decrease until day 14. cOBS-cultivated stem cells showed significantly higher expression levels on days 4 and 7 (2.7-fold and 2.2-fold increase) compared to hMSC cultivated in regular osteoblast growth medium only (Fig. 2D). As an indicator for a more advanced osteogenic differentiation, we analyzed Runx-2 mRNA expression, as this marker represents an osteogenic TCF. We found no differences in the expression levels of Runx-2 between hMSC treated with cOBS and osteoblast growth medium, respectively, over a time course of 14 days (Fig. 2E). In a similar manner to the expression profiles of ALP and LR, BSP-2 mRNA transcripts revealed an inclined gradient over time. hMSC treated with cOBS showed significantly higher BSP-2 levels than the respective controls (between 1.2-fold and 3.2-fold) (Fig. 2F). The late-stage osteogenic marker OCN was moderately induced at day 4 (1.7-fold), whereas at days 1, 7, and 14 no changes could be observed (Fig. 2G). Taken together, stimulation of hMSC with cOBS resulted in higher expression levels of all examined osteogenic markers except for Runx-2.
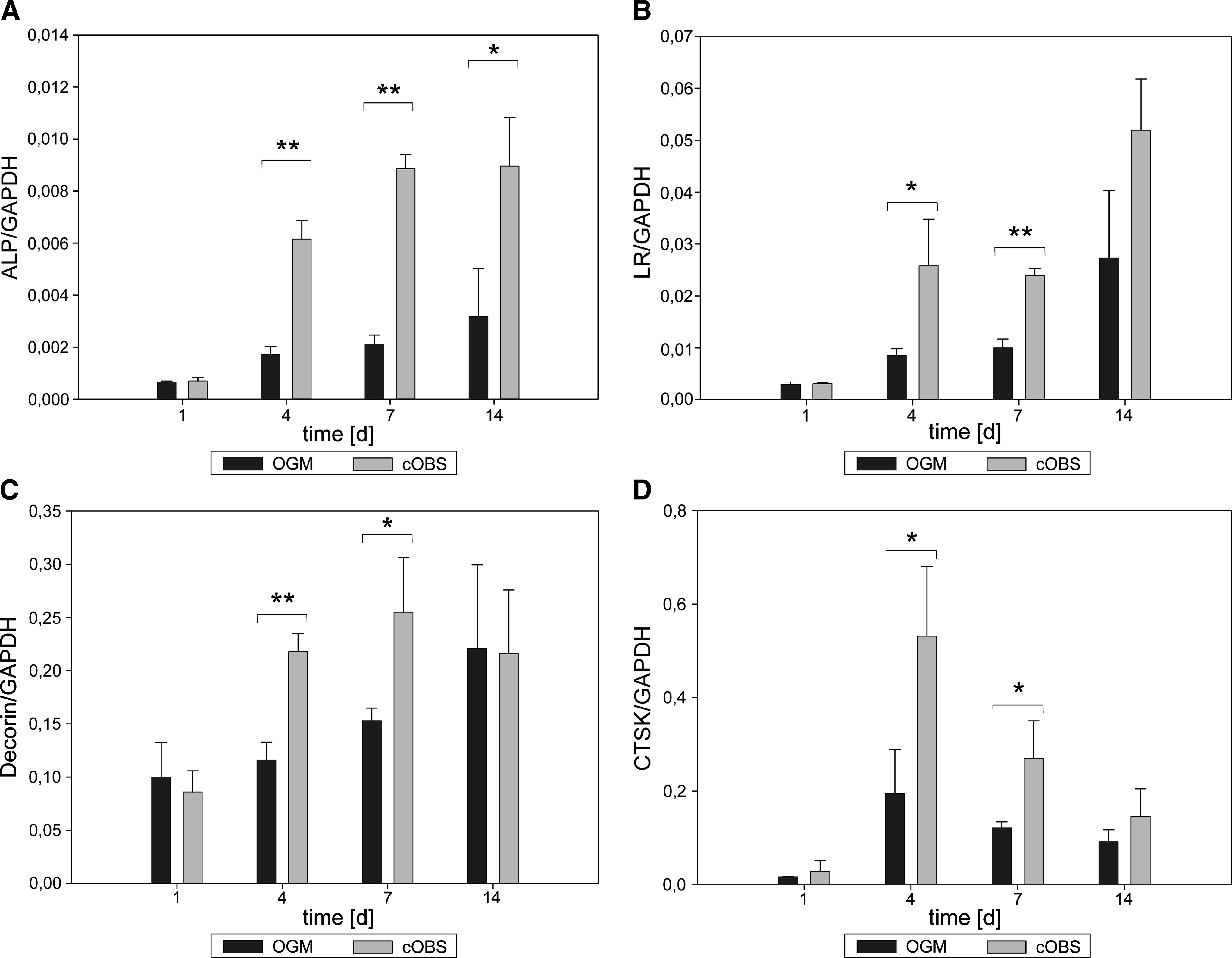
Transcriptional changes of human mesenchymal stem cells (hMSC) after treatment with osteoblast-conditioned supernatant (cOBS) and osteoblast growth medium (OGM) that served as control. (
cOBS experiments with a second hMSC/hOB donor pair
To exclude putative hMSC and hOB donor-related effects, we carried out a set of experiments with a second pair of hMSC and hOB. Applying hOB-conditioned medium under the same settings described above, we analyzed the expression profile of the osteogenic markers (ALP, BSP-2, CTSK, DCN, LR, OCN, and Runx-2). As shown in Table 1, the early osteogenic markers ALP and BSP-2 as well as the late osteogenic marker OCN were induced. Analogically, LR mRNA expression analysis revealed similar results as obtained with the first donor pair. Further, differences in copy numbers of DCN and Runx-2 between control hMSC and cOBS-treated hMSC were only detectable within the standard deviations. Taken together, these experiments indicate that secreted factors from hOB could induce the expression of characteristic osteogenic markers in hMSC, independent of hMSC or hOB donor characteristics.
In parallel, cOBS II was obtained from a hOB II. Induction rates were calculated by the respective copy number ratio of cOBS II–treated hMSC II and control hMSC II.
Statistically significant induction rates were specified with ap <0.05 or bp < 0.01.
hMSC II, Human mesenchymal stem cells of a second donor; cOBS II, human osteoblast–conditioned supernatant II; hOB II, second human osteoblasts donor.
Indirect coculture of hMSC and hOB
To assess whether indirect coculture of hMSC and hOB results in an osteogenic differentiation of hMSC similar to that obtained with cOBS, a transwell assay system was established. To prevent cellular migration on the one hand, but to guarantee the exchange of secreted molecules on the other hand, transwell inserts with a pore diameter of 0.4 μm were used. Two different numbers of hOB were seeded on the bottom of the lower compartment to evaluate whether the hOB number exhibits any influence on the differentiation behavior of hMSC. Expression levels of the osteogenic markers were analyzed by quantitative RT-PCR on days 4, 7, and 14 after starting the indirect coculture experiments. As depicted in Figure 3A, ALP expression showed a significant increase throughout the complete monitored time frame. On day 4, hMSC exhibited a fourfold higher ALP expression level. After 7 days, hMSC indirectly cocultivated with 64,000 hOB exhibited a 3.9-fold higher expression of ALP, whereas the double number of hOB (128,000) resulted in a 7.6-fold increase. At day 14, a 3-fold and 3.2-fold increase of ALP expression was observed when compared to the controls (Fig. 3A), indicating no longer a relation to the hOB number in the lower transwell compartment. Moreover, the expression profile of LR displayed similar characteristics during the indirect coculture experiments. The highest LR expression levels were observed after 14 days. The LR transcript numbers raised independently to the hOB numbers at day 7, whereas at day 14 a slight hOB number-dependent increase of LR expression was observed (Fig. 3B). The analysis of DCN expression revealed a slight increase over time, yet no explicit changes, when hMSC were indirectly cocultured with hOB (Fig. 3C). In a similar fashion to the ALP and LR expression, BSP-2 mRNA levels increased over time. Moreover, at days 7 and 14 high hOB numbers (128,000) resulted in higher induction rates of BSP-2 (2.4-fold and 8.9-fold), whereas low hOB numbers (64,000) still induced BSP-2 copy numbers at days 7 and 14, although in a less pronounced manner (2.0-fold and 3.3-fold) (Fig. 3D). Finally, the expression of the late osteogenic marker OCN was significantly upregulated under indirect coculture conditions in the presence of 128,000 hOB (Fig. 3E). Thus, these results corroborated our findings observed with cOBS. Further, different numbers of hOB may influence—at least in part—the expression levels of some osteogenic markers (e.g., ALP, BSP-2, LR, and OCN).

Differentiation behavior of hMSC kept in a Transwell coculture system. About 64,000 hMSC were seeded on the transwell membrane in the upper compartment, whereas 64,000 and 128,000 hOB were plated at the bottom of the lower compartment resulting in 40% and 80% confluence, respectively. (
Microarray analysis
To obtain more comprehensive insights into transcriptional profile changes during differentiation, microarray analysis was performed using Human Osteogenesis Oligo GEArrays. For this purpose, three different groups consisting of native hOB and hMSC treated with ODM or with cOBS were analyzed over 14 days. All observed mean values were normalized to the respective values of nondifferentiated hMSC that were grown in hMSC growth medium without any additional supplement. On the basis of these normalized expression data, we further analyzed the expression values of genes that displayed ratios greater than 2 or lower than 0.5. As given in Table 1, a total of 15 genes were induced in comparison to untreated hMSC. Of these, five accounted for hMSC treated with ODM, seven for hOB, and three for hMSC stimulated with cOBS. Forty-two genes displayed ratios lower than 0.5 after normalization on control hMSC: 19 exhibited a lower expression level in hOB, and 5 or 18 in hMSC treated with ODM and cOBS, respectively.
These data revealed on the first glance that in differentiated cell entities more genes displayed lower expression levels in comparison to untreated hMSC. Secondly, in hOB and cOBS-treated hMSC a higher number of genes exhibited a more similar expression behavior than ODM-treated hMSC. Thus, more similarities were found between the expression profile of hOB and cOBS-treated hMSC than between hOB and ODM-treated hMSC, suggesting again that hOB-derived factors could induce early osteogenesis in hMSC. In particular, some important genes of the table should be noted. In agreement with previous results, genes of bone homeostasis like ALP or calcitonin receptor (CALCR) were exclusively upregulated in hOB as well as in hMSC stimulated with cOBS. The cysteine peptidase CTSK (dot patterned in Table 2) was slightly increased in these samples as well, but its ratio did not overcome the limitation of 2.0. Eventually, SMAD9, a TCF and regulator of the BMP signaling pathway, showed significantly enhanced ratios in hOB and hMSC stimulated with cOBS, whereas in hMSC treated with ODM no signal could be detected (Table 1).
The expression levels of all detected genes were normalized using the expression values obtained from untreated hMSC. Ratios higher than 2 (dark gray) or lower than 0.5 (light gray) were enrolled for further evaluation. In addition, CTSK ratios (dot patterned) were included as this marker was also evaluated by qRT-PCR. Microarray data were quantified using ArrayVision 6.0 software (Imaging Research). ODM, osteogenic differentiation medium; hOB, human osteoblasts.
Wnt signaling and osteogenesis
Based on the fact that several components of the Wnt pathway have been associated with bone metabolism, we analyzed whether the canonical Wnt pathway might be essential for the early phase of osteogenic differentiation of hMSC. Because loss-of-function mutations in LRP5 result in severe osteoporosis 27 and gain-of-function mutations cause an increase in bone mass, 28 we knocked down LRP5 in hMSC by transfecting LRP5-specific siRNA. Moreover, because β-catenin is a downstream mediator of LRP5 and the central key player of the canonical Wnt pathway, we carried out RNA interference against human β-catenin as well, according to our recently established RNAi protocol. 20 Negative control siRNA-transfected hMSC (nc-hMSC) exhibited the highest ALP expression levels when compared to the LRP5 and β-catenin knockdown approaches. After knockdown of the Wnt coreceptor LRP5, we observed a dramatic drop (about 16-fold) in ALP expression. Similarly, RNAi against β-catenin was associated with a decrease of ALP expression (fivefold) (Fig. 4A). In contrast, the expression levels of LR were upregulated upon knockdown of either LRP5 or β-catenin. In the case of RNAi against LRP5, we observed a slight increase in LR expression, whereas knockdown of β-catenin entailed a substantial upregulation of LR (Fig. 4B). Moreover, RNAi against LRP5 or β-catenin resulted in a slight increase of DCN expression (2.2-fold and 1.5-fold) (Fig. 4C). Similarly, CTSK expression levels raised significantly after knockdown of LRP5 and β-catenin (3-fold and 3.3-fold), respectively (Fig. 4D). RNAi-mediated inhibition of Wnt signaling also came along with an increase of Runx-2 expression (Fig. 4E). Knockdown approaches of β-catenin resulted in higher BSP-2 copy numbers (2.3-fold). However, RNAi against LRP5 showed no changes of BSP-2 expression levels in comparison to control hMSC (Fig. 4F). Manipulation of the Wnt pathway did not influence OCN expression patterns (Fig. 4G). Taken together, the siRNA approaches revealed that under these conditions only ALP seems to be positively regulated by the Wnt pathway in cOBS-treated hMSC, whereas the other osteogenic markers are negatively affected by canonical Wnt signaling.

Inhibition of the Wnt signaling pathway and consequences on osteogenic marker expression. hMSC transfected with siRNA against β-catenin and LRP5 were cultivated for 1 week in osteoblast growth medium or cOBS until RNA was isolated for qRT-PCR. hMSC transfected with negative control siRNA served as controls. (
Discussion
Stem cells can be characterized by two major attributes—namely, the capacity of self-renewal and differentiation. 29 Differentiation processes take place in niches where other cell types of the microenvironment are involved in the regulation of these attributes. 12 However, little is known about the interactions between hMSC and other cell types that reside in the environmental niche of the bone. According to our working hypothesis that cells of the differentiation niche need to be located in the direct vicinity of hMSC in vivo, we analyzed the functional role of hOB in the control of hMSC differentiation. As cells rarely respond to a single stimulus, we investigated whether a cocktail of soluble factors produced by hOB that reside in the immediate environment of hMSC is sufficient to induce early osteogenic markers in hMSC.
Basically, cells of the osteoblastic lineage traverse different stages starting with MSC until they undergo terminal differentiation into osteocytes. In early stages they represent osteoprogenitor cells; later, preosteoblasts; and in a more mature stadium, they are termed “osteoblasts.” 30 To determine the differentiation status of the cells used in our study, Alizarin-red-S staining of hOB was performed, and it revealed that stimulation of hOB with complete differentiation medium or cultivation medium supplemented with dexamethasone and β-glycerol-phosphate was indispensable for matrix mineralization. According to the before-mentioned classification, osteoblasts should be able to mineralize their surrounding matrix. 31 Based on this criterion, our data suggest that the hOB used in the present study can most probably be attributed as preosteoblastic cells.
hMSC possess the capacity to differentiate along the mesodermal lineage into chondrocytes, adipocytes, or osteoblasts.10,32 Moreover, some reports suggest that hMSC could differentiate even across the germ layer, giving rise to cell types with visceral mesoderm, neuroectoderm, or endoderm characteristics. 33 Our own work demonstrated that the mesodermal differentiation capability could be maintained after genetic modifications 25 as well as after stimulation of hMSC with recombinant Wnt3a. 20
Following our working hypothesis that environmental factors are substantially involved in the regulation of stem cell characteristics, we verified the differentiation capacity of commercially purchased hMSC within the mesodermal germ layer (data not shown) and confronted the cells with soluble factors delivered by hOB. hMSC stimulated with cOBS showed higher expression levels of osteogenic markers (ALP, BSP-2, OCN, LR, DCN, and CTSK) except for that of the TCF Runx-2. ALP is a well-established early osteogenic marker 34 as well as DCN that was originally isolated from bone and cartilage tissue. 35 The hormone leptin is produced by adipocytes and regulates osteogenic differentiation and the inhibition of adipocyte formation via the LR. 36 Further, osteoclasts being involved in bone destruction produce and secrete collagenolytic proteinases like CTSK. 37 Nevertheless, osteoblasts can also acquire that function to recycle inadequately processed collagen type I. 38 BSP-2 represents a protein mainly found in the surrounding matrix of newly formed osteoblasts. Adapted from the finding that BSP-2 is also involved in mineral crystal formation, it reflects a reliable marker for osteogenic differentiation. 39 Based on our observations that early osteogenic markers were upregulated with cOBS, secreted factors of hOB seem to be responsible for the initiation of early osteogenesis in hMSC. This finding is corroborated by the fact that the late-stage osteogenic TCF Runx-2 24 was not upregulated in this early differentiation process. In line with this observation, OCN was only slightly induced. As a late-stage osteogenic differentiation marker, this noncollagenous calcium-binding bone protein is normally secreted from osteoblasts at advanced stages of osteogenesis. 40 To evaluate putative donor-related variations, additional experiments with a second hMSC/hOB donor pair were accomplished. In this setting, the induction of the osteogenic markers in hMSC by cOBS appeared earlier (already at day 1, see Table 1) when compared with the first donor pair. However, these type of time-related differences in triggering early osteogenic markers in hMSC are not unexpected, because primary cell populations display heterogenous cellular phenotypes.
In addition, indirect coculture experiments were often used to mimic the interplay of soluble factors between different cell types for monitoring changes in the differentiation status. In this regard, it has been recently demonstrated that hMSC build up more organized filamentous structures, when being indirectly cocultered with endothelial cells, whereas direct coculture of both cell populations lead to a disorganization of α-actin filaments in hMSC. 41 To analyze whether hOB could influence the differentiation activities of hMSC in a similar way as cOBS, we established an indirect coculture approach. Four (ALP, BSP-2, LR, and OCN) out of five analyzed markers exhibited higher expression levels when hMSC were cultivated in the presence of hOB. This indicates that—similar to our results obtained with cOBS—the niche-like environmental conditions of the coculture assay are sufficient for triggering early osteogenesis of hMSC. However, the expression of CTSK did not change regardless of the applied hOB number in the lower compartment, indicating some differences between treatment of hMSC with cOBS and the indirect coculture approach.
To monitor a broader spectrum of osteogenic markers, an osteoblast-specific microarray was performed on hOB and hMSC treated either with cOBS or ODM, respectively, or stem cell growth medium as a control. The validity of the microarray results was evaluated by qRT-PCR on selected genes (e.g., ALP) of hMSC after stimulation with cOBS and ODM (data not shown). In general, two major conclusions can be drawn from the microarray experiments: on the one hand, more genes were downregulated in hMSC that were stimulated with cOBS when compared to untreated hMSC; on the other hand, cOBS drives differentiation of hMSC into a more hOB-like phenotype than ODM. Some genes that exhibited a high expression in hOB and cOBS-treated hMSC belong to the group of early osteogenic markers like ALP 42 and proteases that are in charge of bone remodeling like CTSK. 38 Moreover, we found CALCR and SMAD9 to be upregulated in hOB and cOBS-treated hMSC. Whereas CALCR plays a major role in maintenance of calcium homeostasis, 43 SMAD9 belongs to the receptor-regulated SMAD proteins that are in involved in the TGF-β/BMP signaling pathway. 44 As this pathway is of pivotal importance in osteogenic differentiation, 45 this finding provides further evidence that soluble factors that had been produced by osteoblasts could induce early osteogenic markers in hMSC.
With regard to the molecular mechanisms of osteogenesis, there is growing evidence that canonical Wnt signaling is largely involved in the regulation of this differentiation process.46,47 Consistent with this, it has been reported that loss-of-function mutations in LRP5, an important coreceptor of Wnt signaling, leads to severe osteoporosis, 22 whereas gain-of-function mutations result in an increase of bone mass.27,28 To examine a possible functional interaction between Wnt signaling and preosteoblastic differentiation in hMSC, we performed RNAi against β-catenin and LRP5 as previously described.20,48 Knockdown of either β-catenin or LRP5 resulted in a decrease of ALP and an increase of BSP-2, LR, CTSK, and Runx-2 expression, whereas the expression levels of DCN and OCN were not affected by this approach. Recently, it has been reported that ALP expression could be regulated by canonical Wnt signaling in hMSC; 49 thus, our RNAi results point to an activation of this pathway upon stimulation with cOBS. Because in both knockdown approaches the transcriptional induction of the ALP-expression by cOBS was not evident any more, these observations indicate a participation of the Wnt pathway via its co-receptor LRP5 and the central mediator β-catenin.
In contrast, the analysis of the other examined genes exhibited a transcriptional induction after knockdown of LRP5 and β-catenin when treated with cOBS. These observations suggested a distinct regulation of the expression of these osteogenic differentiation-associated genes. Whereas ALP seems to be positively regulated by the canonical Wnt pathway, the transcript levels of LR, CTSK, and Runx-2 revealed to be negatively influenced under these conditions. Under these experimental settings the ECM proteins DCN and OCN are not affected by canonical Wnt signaling. In our setting the β-catenin knockdown displayed more pronounced effects than that of LRP5. This indicates that apart from the changes induced by Wnt ligands after binding to its co-receptor LRP5, other signaling cascades might be implicated. In line with these findings literature reports refer to the crosstalk between the Wnt- and the BMP2-mediated TGFβ pathway.50–54 In this context, Jian et al. showed in hMSC that the translocation of β-catenin from the cytoplasm into the nucleus is induced by Smad3-mediated TGFβ signaling. 50 Thus, it is conceivable that in our conditioned medium and indirect coculture approaches, beside the canonical Wnt pathway, also other signal transduction cascades may be involved in the regulation of early osteogenic markers in hMSC.
In summary, our data provide substantial evidence that hOB produce and secrete soluble factors that influence the differentiation behavior of hMSC, probably similar to the setting in the natural environmental differentiation niche. In detail, we could show for the first time that cOBS-treated hMSC exhibited a transcriptional profile quite alike to that of hOB, whereas stimulation of hMSC, according to a so far established protocol implying different chemical compounds, 55 initiates a quite different osteogenic differentiation pattern. Thus, with regard to a therapeutic setting, an alternative differentiation approach using cOBS should be considered. This may pave the way for a more in vivo–related differentiation procedure being of fundamental importance for the generation of functional osteoblast-like cells.
Footnotes
Acknowledgments
We thank Lourdes Ruiz-Heinrich for her excellent technical assistance. This work was partially supported by the “Molecular Medicine” grant of the Medical Faculty of the Ludwig-Maximilians-University of Munich to M. Ilmer and P. Neth.
Disclosure Statement
No competing financial interests exist.