
Abstract
There is increasing interest in developing new in vitro tissue models using typical tissue engineering approaches. This study was designed to (1) develop a novel three-dimensional (3D) in vitro model of bone by seeding murine primary osteoblasts and osteoclast precursors on a resorbable porous ceramic scaffold based on silicon-stabilized tricalcium phosphate (Skelite®), and (2) investigate bone cell interactions in a 3D environment mimicking an in vivo condition and compare it to traditional two-dimensional (2D) cultures. Murine primary osteoblasts from C57Bl6/J mice and osteoclast precursors from C57Bl/6-Tg(ACTB-EGFP)1Osb/J mice were co-cultured on 3D Skelite scaffolds and on standard plastic culture dishes. The differentiation of these cells in both culture conditions was compared by histology (hematoxylin–eosin staining and polarized light analysis), immunohistochemistry (collagen type I), and gene expression analysis by real-time PCR for Runt-related transcription factor 2, osterix, osteocalcin, cathepsin K, and tartrate resistant acid phosphatase. To analyze and compare bone turnover in 3D and 2D co-cultures, we evaluated the modulation of RANKL and OPG mRNA expression. We observed an enhancement of osteoblast differentiation in the 3D mineralized environment that in turn promoted earlier osteoclast differentiation. In this paper, we also report that the increased osteoblast differentiation in the 3D model led to a deposition of extracellular matrix that faithfully reflected the morphology of bone tissue.
Introduction
In the case of bone, different porous ceramic scaffolds can be adopted to mimic the tissue mineralized ECM in newly developed in vitro models to be employed to study bone physiology and pathophysiology and to test potential drugs for the treatment of bone metabolic diseases. Bone substitutes have to be osteoconductive and osteoinductive. Recent studies demonstrated that Skelite® (Octane Orthobiologics, Kingston, Ontario, Canada), a biomaterial based on silicon-stabilized tricalcium phosphate, has these characteristics and represents a good substrate to mediate bone regeneration. Skelite constructs were implanted in critical-sized defects of sheep tibias, either alone or after seeding with bone marrow stromal cells (BMSC). 1 Only BMSC-seeded ceramics displayed a progressive osteoclast-mediated scaffold resorption, coincident with new bone deposition. To investigate the coupled mechanisms of bone formation and scaffold resorption, BMSC-seeded ceramic cubes were implanted in immunodeficient mice, recovered after 2 or 6 months, and analyzed by X-ray computed microtomography with synchrotron radiation (μCT) and μX-ray diffraction. With increasing implantation time, scaffold thickness significantly decreased while bone thickness increased. μX-ray diffraction revealed resorption of the bioceramic only in the contact areas between deposited bone and scaffold, and that scaffold resorption did not occur at areas away from the bone surface, highlighting coupling and cell dependency of the resorption and matrix deposition mechanisms. 1
Bone turnover is a well-balanced process that smoothly coordinates resorption and formation of skeletal tissue where the crosstalk between osteoblasts and osteoclasts plays a key role. Osteoporosis, a major worldwide health problem affecting 4 million to 6 million women and 1 million to 2 million men in the United States and comparable numbers of patients in Europe, occurs when bone resorption exceeds bone matrix deposition, thus resulting in a negative skeletal balance. 2
This study was designed to develop a novel in vitro 3D model by co-culturing murine primary osteoblasts and osteoclast precursors on Skelite scaffolds to reflect more faithfully a bone physiological microenvironment than in the traditional Petri dish–based counterpart cultures. The final aim was to evaluate the effects of a mineralized 3D environment on osteoblast and osteoclast differentiation. We investigated the gene expression modulation of osteogenesis (Runt-related transcription factor 2 [Runx2], osterix [Osx], and osteocalcin) and osteoclastogenesis (cathepsin K and tartrate resistant acid phosphatase [TRAP]) markers at different times during both 3D and 2D co-cultures. Finally, we analyzed in both culture conditions the modulation of receptor activator of nuclear factor NF-kB ligand (RANKL) and osteoprotegerin (OPG) expression in response to an osteogenic stimulus.
The relevance of RANK/RANKL/OPG signaling in osteoclast maturation and bone remodeling has been reported in different studies, increasing the interest in developing new therapies by targeting this pathway.3–7 RANKL is a cytokine of the tumor necrosis factor family that is expressed by osteoblasts and induces osteoclast maturation through RANK binding.8,9 OPG is a basic glycoprotein of the tumor necrosis factor receptor superfamily that acts as soluble decoy receptor for RANKL and thus inhibits osteoclast formation.8,9 The development of an in vitro model of bone where these genes are physiologically modulated could represent an useful tool not only to investigate molecular mechanisms driving bone turnover but also to study the effects of new drugs, such as anti-osteoporotic drugs, on this complex biological process.
Materials and Methods
Materials
Dulbecco's modified Eagle's medium (DMEM), L-glutamine, trypsin/EDTA, and penicillin/streptomycin were obtained from Euroclone (Euroclone Life Sciences Division, Milan, Italy). Fetal calf serum (FCS), collagenase I, and collagenase II were from Gibco (Invitrogen s.r.l, Carlsbad, CA). Ascorbic acid, dexamethasone, and β-glycerophosphate were purchased from Sigma-Aldrich (St. Louis, MO). Skelite disks (67% Si-TCP and 33% HA/B-TCP) were a gift from Octane Orthobiologics (Kingston, Ontario, Canada). Skelite presents an interconnected, open pore structure similar to cancellous bone and has a porosity level of approximately 60% with a pore size range between 200 and 500 μm.
Animals
Murine primary osteoblasts were obtained from C57Bl/6 mice; osteoclast precursors were from C57Bl/6-Tg(ACTB-EGFP)1Osb/J mice, a strain constitutively expressing the green fluorescence protein (GFP). All mice were purchased from Jackson Laboratory (Ban Harbor, MA).
Cell isolation and culture
All primary cultures used in this work were obtained from at least five mice. All experiments were independently repeated at least two times using different primary cultures.
Osteoblasts isolation
Osteoblasts were enzymatically isolated from calvariae of 2–5-week-old C57Bl/6 mice. Briefly, skin and soft connective tissue were removed. Bone was cut into square pieces of 4 mm2 and washed two times in Ringer Solution (147 mM NaCl, 4 mM KCl, and 1.13 mM CaCl2, pH 7.4). The bone pieces were sequentially digested in a trypsin (25 mg)/collagenase I (30 mg)/collagenase II (30 mg) in 25 mL Ringer Solution at 37°C for 10, 20, 30, 50, and 70 min. Osteoblasts obtained from the last two digestions were pooled and plated together in 90-mm-diameter tissue culture dishes. The cells were cultured until confluence in DMEM supplemented with 10% FCS, 2mM L-Glutamine, and penicillin/streptomycin 50 μg/mL (indicated as “normal medium”). After confluence, isolated cells were trypsinized and subsequently used for 2D and 3D co-cultures.
Osteoclast precursors isolation
Bone marrow cells were isolated flushing the femurs and the tibias of 3–5-week-old C57Bl/6-Tg(ACTB-EGFP)1Osb/J mice with 5 mL of phosphate buffer solution (PBS; 137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, and 2 mM KH2PO4, pH 7.4). Cells were centrifuged for 10 min at 260 g and washed three times with PBS. Cells were then incubated with CD34 MicroBeads, and CD34− cells were selected by collecting the cells not retained on MS magnetic columns by MACS system (Miltenyi Biotec, Auburn, CA). 10
Three-dimensional co-cultures
About 4 × 105 osteoblasts and 4 × 105 osteoclast precursors were resuspended in 50 μL of normal medium and seeded on Skelite disks (9 mm diameter; 1.2 mm height) placed at the bottom of wells of 24-well plate coated with 1% of agarose. After 3 h of incubation at 37°C, 2 mL of medium was carefully added. Disks were maintained in culture at 37°C with 5% CO2. Medium was changed three times a week.
Two-dimensional co-cultures
Osteoblasts and osteoclast precursors were mixed with a ratio of 1:1 (5 × 104 cells for each cell population), plated in six-well plates, and cultured in normal medium at 37°C with 5% CO2. The medium was changed three times a week.
Osteogenic stimulation
Two-dimensional and 3D co-cultures were maintained for 10 days in normal medium followed by either 30 or 60 days (see Results and Discussion sections) in osteogenic medium (DMEM supplemented with 10% FBS, 2 mM L-glutamine, penicillin/streptomycin 50 μg/mL, ascorbic acid 50 μg/mL, β-glycerophosphate 1.5 mg/mL, and 10−7 M dexamethasone).
A complete description of all adopted culture conditions is reported in Table 1.
RNA extraction
Total RNA was extracted from 2D co-cultures using RNeasy Plus Mini Kit (Qiagen, Hilden, Germany) following the manufacturer's instructions. To extract RNA from 3D co-cultures, the disks were mechanically shattered in RNeasy Plus Mini Kit lysis buffer and centrifuged; the supernatant was subjected to RNA extraction by the same kit as described in the manufacturer's instructions.
Real-time quantitative reverse transcriptase—polymerase chain reaction
The gene expression analysis were performed by real-time quantitative RT- PCR using the PE ABI PRISM 7700 sequence detection system (Perkin-Elmer, Waltham, MA) and SYBR Green (Applied Biosystems, Foster City, CA). Forward and reverse primers were designed by the Primer Express Software (Applied Biosystems). The sequences of forward and reverse primers were as follows: Runx2, 5′-GCACTGGCGGTGCAACAAG-3′ and 5′-CCATGACGGTAACCACAGTC-3′; Osx, 5′-ATGGCGTCCTCTCTGCTTG-3′ and 5′-GCAGGCTGCAGTCAGCATG-3′; osteocalcin, 5′-CAGCGGCCCTGAGTCTGA-3′ and 5′-CCTCCTGCTTGGACATGAA-3′; cathepsin K, 5′-GGAGCTATGGAAGAAGACTC-3′ and 5′-AACGCCGAGAGATTTCATCC-3′; TRAP, 5′-CTACCTGTGTGGACATGACCA-3′ and 5′-GCACATAGCCCACACCGTTC-3′; RANKL, 5′-GTGGTCTGCAGCATCGCT-3′ and 5′-CACTCTCCAGAGTCGAGTC-3′; OPG, 5′-ATGCAACACATGACAACGTG-3′ and 5′-CTGAAGAAGGCCTCTTCACA-3′; GAPDH, 5′-TGTGTCCGTCGTGGATCTG-3′ and 5′-GATGCCTGCTTCACCACCTT-3′; GFP, 5′-AAGTTCATCTGCACCACC-3′ and 5′-TCCTTGAAGAAGATGGTGCG-3′. The expression of GAPDH was examined as endogenous control. The GFP expression was examined to monitor the cell-type ratio during the cultures. Relative transcripts levels were calculated from the relative standard curve constructed from stock cDNA dilutions and divided by the target quantity of the calibrator following manufacturer's instructions.
Histology, histochemistry, and immunohistochemistry
Skelite disks were fixed with 4% formaldehyde in PBS for 3 h at 4°C, totally decalcified with Osteodec (Bio Optica, Milan, Italy) and routinely embedded in paraffin. Five-μm sections were stained with hematoxylin–eosin and viewed in transmitted and polarized light microscopy. For immunohistochemical analysis, the sections were immunolabeled with anti-collagen type I Ab (Calbiochem, Darmstadt, Germany) at 25°C for 1 h. System Labelled Polymer-HRP Anti-Rabbit (Dako, Glostrup, Denmark) and Liquid DAB Substrate Chromogen System (Dako) were used for detection.
For TRAP staining, specimens were processed undecalcified. The samples were fixed in 3.7% paraformaldehyde in PBS and embedded at low temperature (4°C) in methylmethacrylate. Seven-μm slices were cut with a Jung model K microtome (Reichert-Jung) and used for TRAP staining (without counterstaining) of osteoclasts, or Hoechst-nucleus staining.
TRAP staining of 2D co-cultures was performed using Leukocyte Acid Phosphatase (TRAP) Kit (Sigma-Aldrich) according to manufacturer's instructions.
Bone morphometry
Skelite disks were fixed, decalcified, and routinely embedded in paraffin as described above. Five-micron sections were stained with hematoxylin–eosin and viewed in transmitted light. Twenty random fields per disk were analyzed using Axiovision release 4.5 software to calculate the bone matrix deposition area (BMA) and the total cellular network area (TCA) in each field.
Statistical analysis
The data were analyzed by two-tailed t-tests. Differences were accepted to be statistically significant at p ≤ 0.01.
Results
Three-dimensional cultures of primary murine osteoblasts and osteoclast precursors on Skelite disks
Initial experiments were done to evaluate if static seeding could ensure a homogenous cell colonization of the scaffolds. Skelite disks containing statically seeded osteoblasts were maintained for 30 days in normal medium (Table 1: III). To analyze the cell viability and their distribution within the disks, at the last day of culture, we stained the cells with Hoechst 33258, a fluorescent vital stain. Positively stained cell nuclei stained positively and were homogeneously distributed throughout the disks (Fig. 1A). We also performed hematoxylin–eosin staining of decalcified paraffin-embedded disk sections (Fig. 1B). Depending on their localization, cells appeared with different morphology; cells grown in tight contact with the biomaterial showed bone lining cell-like morphology, whereas cells grown within pores appeared as a cellular network of fibroblast-like cells.

Osteoblasts cultured for 30 days on Skelite scaffolds. (
Further experiments were designed to verify the capability of osteoblast and osteoclast precursors to proliferate and differentiate on Skelite scaffolds. Primary osteoblasts were obtained from bone chips of wild-type C57Bl/6. CD34− osteoclast precursor cells were sorted by CD34+ immunodepletion from the bone marrow of GFP transgenic mice C57Bl/6-Tg(ACTB-EGFP)1Osb/J. To preliminary test the potential of the sorted GFP+ CD34− cells to become mature osteoclasts, these cells were plated together with primary osteoblasts in standard culture Petri dishes with normal culture medium and stained for TRAP activity staining after 30 days. As expected, GFP+ TRAP+ cells were clearly evident after the 30 days of co-culture (data not shown).
To evaluate cell differentiation, we analyzed the level of mRNA expression of different genes involved in osteoblast (Runx2, Osx, and osteocalcin) and osteoclast differentiation (cathepsin K and TRAP) in both 2D and 3D co-cultures. Runx2, Osx, and osteocalcin are markers of the early, middle, and late phase of osteoblast maturation, respectively, while cathepsin K and TRAP are expressed by fully differentiated osteoclasts. Primary murine osteoblasts and osteoclast precursors were co-seeded on Skelite scaffolds or co-plated in plastic Petri dishes. Half of the cultures were maintained for 40 days in normal medium (Table 1: IV and V) and the other half were maintained for 10 days in normal medium followed by 30 days in osteogenic medium (Table 1: I, II, VI, and VII). RNA samples were collected after 10 and 40 days (Table 1: I, II, IV, V, VI, and VII). Given that the initial ratio between osteoblasts and osteoclast precursors at the time of the cell seeding was known, and on the assumption that osteoblasts and osteoclasts were expressing comparable level of the house keeping enzyme GAPDH, the ratio between GFP and GAPDH expression was used to assess variations in the percentage of GFP+ and GFP− cells at the two time points (Fig. 2). Therefore, the expression of each marker gene was normalized to GAPDH and calibrated to the percentage of GFP+ or GFP− cells if expressed by osteoclasts or osteoblasts, respectively. Each determined value was subsequently calibrated to the value recorded after 10 days in 2D co-cultures.

Analyses of green fluorescense protein (GFP) expression by real-time PCR. All measures were normalized to the housekeeping gene GAPDH. In the table we report the estimated percentages of osteoblasts and osteoclast precursors at all time points analyzed. White bars: 2D co-cultures. Striped bars: 3D co-cultures. Each bar is the mean ± SD of n = 3–6 samples. ap < 0.01.
In the 3D co-cultures performed in normal medium, the estimated percentages of osteoblast at 10 and 40 days were 77 ± 9% and 66 ± 5%, respectively. In the 2D co-cultures maintained in normal medium, we observed a lower estimated percentage of osteoblasts, 50 ± 8% after 10 days and 37 ± 6% after 40 days. On the contrary, after the culture in osteogenic medium, the percentage of osteoblast estimated in the 2D co-cultures was comparable with that calculated in the 3D co-culture system, 78 ± 8% and 77 ± 16% (Fig. 2).
In 2D co-cultures, the maximum level of Runx2 expression occurred after 40 days of culture in normal medium (Fig. 3A). In normal medium, after 40-day cultures, the Runx2 mRNA level was 4.2-fold higher than that observed in the same culture system at day 10 and 5.6-fold higher than in the 3D co-cultures at day 40. On the contrary, in the 3D co-cultures the highest level of Runx2 expression was observed after only 10 days. However, no major variations in the level of expression of this gene were recorded at the different culture times. The maximum level of Osx occurred at day 40 in 2D co-cultures performed in normal medium. At this time point the Osx mRNA level was 8.7-fold higher than that recorded in the same culture system at day 10 and 1.5-fold higher than in 3D co-cultures at day 40. In the 3D co-cultures, at day 10, we observed the maximum level of expression for Osx (Fig. 3B). In this culture conditions, at this time, the Osx mRNA level was 6.4-fold higher than in 2D conditions. The level of both Osx and Runx2 mRNA decreased after 30 days of culture in osteogenic medium (Fig. 3A, B).
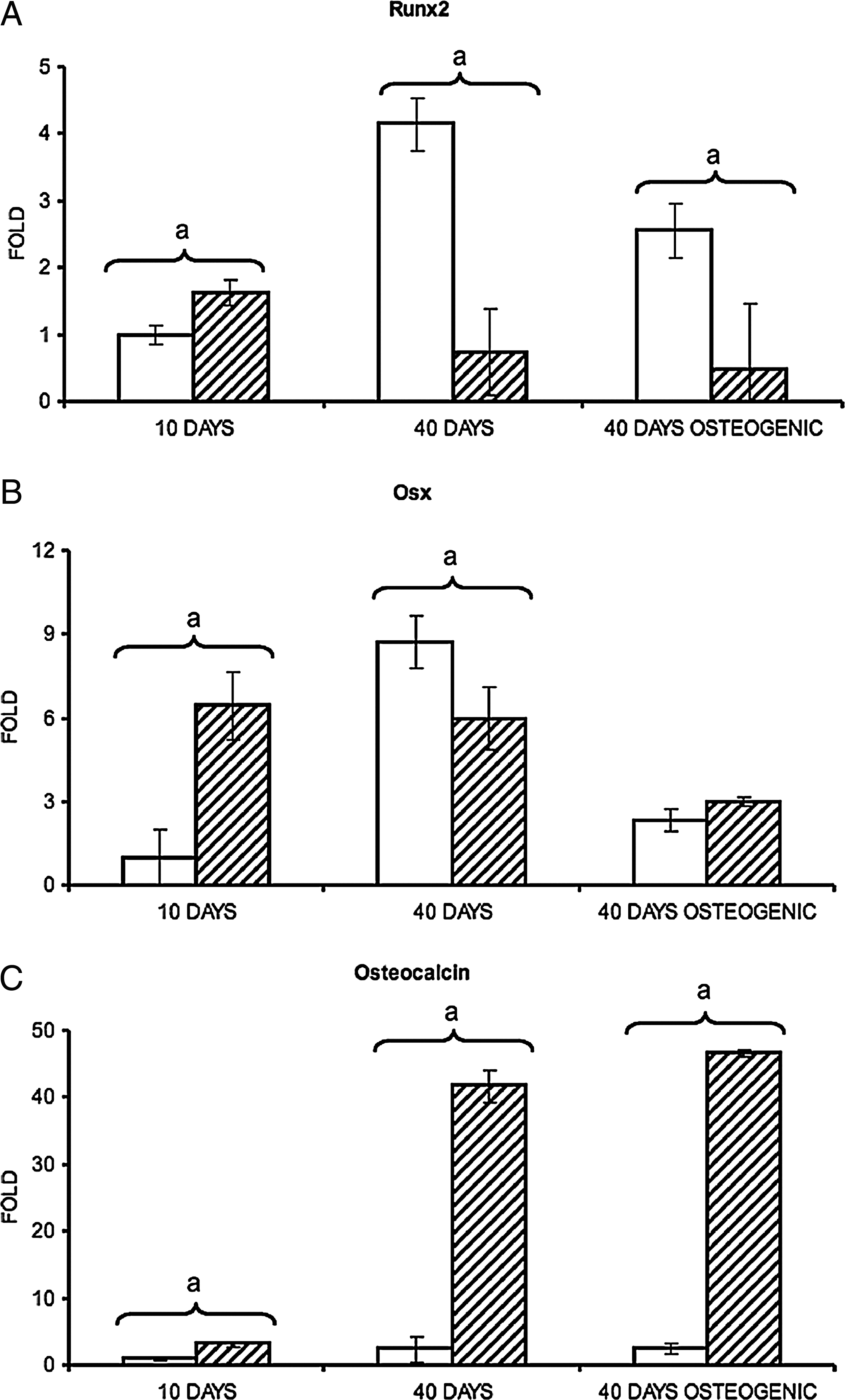
Effect of the 3D environment provided by Skelite scaffolds on the mRNA expression of osteoblast differentiation markers. Analyses of mRNA expression were performed by real-time PCR. All measures were normalized to housekeeping gene expression (GAPDH) and referred to the percentage of GFP− cells (estimated by evaluating GFP expression modulation during the time of the experiment). All results were subsequently calibrated to the expression recorded in samples derived by co-cultures in 2D condition at days 10. (
Afterward, we analyzed the effect of the 3D environment on the osteocalcin (a fully differentiated osteoblast marker) expression (Fig. 3C). At all culture times, the osteocalcin mRNA level was higher in cells co-cultured on Skelite scaffolds compared to those grown in 2D condition independently of the used culture medium. After 10 and 40 days in 3D conditions and normal medium, osteocalcin expression was, respectively, 3.2- and 16.9-fold higher than in 2D co-cultures with the same medium. After 40 days of culture in osteogenic medium, the mRNA level in the 3D co-culture was 19.0-fold higher than the mRNA level in 2D co-culture equally treated.
When we investigated osteoclast maturation by evaluating the expression of cathepsin K and TRAP, we observed the expression of both genes with a trend to a progressive decrease in cells grown on scaffolds at day 10 and 40 (Fig. 4). On the contrary, in 2D co-cultures, cathepsin K mRNA level was 2.6-fold higher at day 40 than at day 10 (Fig. 4A), while the expression of TRAP was barely detectable (Fig. 4B). In 2D conditions the cell stimulation by the addition of the osteogenic medium resulted in an enhanced expression of both osteoclast markers, while the addition of the same medium to the cells grown on Skelite scaffolds did not dramatically affect cathepsin K and TRAP expression (Fig. 4).

Effect of the 3D environment provided by Skelite scaffolds on the mRNA expression of osteoclast differentiation markers. Analyses of mRNA expression were performed by real-time PCR. All measures were normalized to GAPDH expression, referred to the percentage of GFP+ cells, and calibrated to the expression recorded in samples derived by co-cultures in 2D conditions at days 10 (see legend of Fig. 4). (
To compare bone turnover regulation in 2D and 3D co-cultures, we investigated the effect of the osteogenic stimulation on the expression of RANKL and OPG, commonly considered as the two most important genes involved in this biological process. In 3D co-cultures, RANKL expression was almost half of the one observed in the 2D co-cultures, while OPG expression increased twofold. In cells grown as 2D culture, RANKL increased sixfold, while the level of OPG decreased to a barely detectable level (Fig. 5).

Effect of osteogenic stimulation on (
In vitro bone matrix deposition
To further evaluate the effect of osteogenic stimulation on primary murine osteoblasts and osteoclast precursors co-cultured on Skelite disks, we performed an histological analysis (hematoxylin–eosin staining) of paraffin-embedded sections of disks cultured for 10 days in normal medium followed by 30 days in osteogenic medium (Table 1: VII). An eosinophilic ECM on the border of cultured disks was found (Fig. 6A). The same well-organized ECM was never observed in 2D co-cultures (not shown). To evaluate whether organized collagen fibers were present in the samples, the same sections were exposed to polarized light. The ECM resulted positive, indicating a high organization of collagen fibers (Fig. 6B). To further confirm the collagenous nature of matrix, we performed an immunohistochemical analysis with anti–collagen type I primary antibody. Specific immunoactivity in the zone of bone matrix deposition was clearly detectable. Moreover, a lower positive signal was also detected on the Skelite biomaterial, suggesting some collagen type I diffusion through the microporosity of the scaffolds (Fig. 6C). Skelite disks without cells were maintained in the same culture medium and used as negative control to ensure the immunolabeling specificity (Fig. 6C, insert). To confirm the presence of mature osteoclasts after 40 days of co-culture, we performed an histological analysis on undecalcified sections. As expected, few mature osteoclasts were still present and appeared as TRAP+, polynucleated cells (Fig. 6D).

ECM deposition in the 3D co-cultures. Paraffin-embedded disk sections were stained with hematoxylin–eosin and viewed in (
To investigate whether the amount of deposited bone matrix increased during the time of culture, histological sections obtained from Skelite disks cultured for 10 days in normal medium followed by 60 days in osteogenic medium were compared with sections obtained from 40 days cultures in osteogenic or in normal medium (Table 1: V, VII, and VIII) (Figs. 7C, 6A, and 7B, respectively). A section of the unseeded biomaterial is shown as control (Fig. 7A). At 70 days, we observed the presence of a more abundant and organized eosinophilic bone-like ECM both on the border and inside the scaffold (Fig. 7C). The ratio between the BMA and the TCA at 40 and 70 days was determined. At 40 days in normal medium, BMA/TCA was 0.04 ± 0.02; at 40 days in osteogenic medium, 0.05 ± 0.03; while at 70 days in osteogenic medium, the same ratio was 0.11 ± 0.03 (Fig. 7D). In our opinion these results indicate that, as expected, in the 3D cultures, the osteogenic stimulation led to a deposition of a more organized bone-like ECM and that an increase in the bone matrix deposition occurred from 40 to 70 days of culture.

ECM deposition in the 3D co-cultures. Paraffin-embedded disk sections were stained with hematoxylin–eosin. (
Discussion
The traditional 2D cultures have been widely used for decades and played a central role in the progress of various biologic research fields. The results obtained with cells grown in 2D cultures are reproducible and easy to interpret and to compare with results obtained in different laboratories. Unfortunately, this approach cannot capture the relevant 3D complexity of tissues and organs. For these reasons, the demand for in vitro 3D models for better understanding human tissue physiology is increasing.11,12
Efficiency of cell seeding onto biocompatible scaffolds plays an important role in the attainment of functional properties of engineered tissues especially when large constructs are used. Indeed, the mode of seeding influences the distribution and the viability of the cells within the scaffold. Various cell-seeding approaches have been recently investigated, and novel dynamic seeding methods have also been developed.13–15 These procedures are particularly useful to improve seeding efficacy and to ensure a high yield and a good uniformity of cell distribution in therapeutic tissue engineering where scaffolds normally have large dimensions. The thinness of Skelite scaffold utilized in this study encouraged us to adopt a more simple static seeding approach. Murine primary osteoblasts that were seeded this way grew homogeneously and colonized all the pores of Skelite scaffold after 30 days of culture. The viability of cells cultured within the scaffold was confirmed by the Hoechst fluorescent stain.
To study the morphology of primary osteoblasts grown on Skelite scaffolds and to investigate the interaction between the cells and the biomaterial, histological analyses were performed. As expected, cells had different morphology depending on their localization; cells grown in tight contact with Skelite assumed a phenotype similar to that of the bone lining cells.
The possibility to culture primary osteoblasts within the Skelite scaffolds and the good integration between the cells and the biomaterial convinced us to investigate the capability of this type of cells in supporting osteoclast precursors' (preOCs) proliferation and maturation in the 3D culture system compared to the routine 2D cultures. Although the more common approach to co-culturing these cells in monolayer condition is to culture osteoblasts for a specific time period and then to introduce osteoclast precursors cells, we decided to co-seed osteoblasts and osteoclast precursors to have a better homogeneous distribution of the two cell populations within the 3D scaffolds. Osteoclast precursors were derived from monocyte–macrophage lineage cells normally present in bone marrow, commonly considered as CD34− cells. 16 We used osteoclast precursors sorted as CD34− cells from the bone marrow of C57Bl/6-Tg(ACTB-EGFP)1Osb/J mice strain. The mice strain expressing the GFP under control of a β-actin promoter was chosen to have a tricky internal specific marker for the sorted cells. PreOCs can proliferate with the support of factors such as M-CSF and RANKL produced by cultured osteoblasts.17–20 To monitor variations of the ratio between osteoblasts and preOCs in the different co-cultures here analyzed, we evaluated the modulation of the GFP expression during the time of the cultures. In the 3D co-cultures maintained in normal medium, we observed a higher osteoblasts/preOCs ratio compared to the 2D counterpart, thus suggesting an higher capability of the osteoblast, respect to the preOCs, to adhere and proliferate in the mineralized environment provided by the scaffold. After 40 days of co-culture performed in osteogenic medium, we observed a decrease of the preOCs percentage in the 2D cultures respect to the normal medium counterpart. On the contrary, we did not recorded a significant modulation of the preOCs in the 3D co-cultures performed in osteogenic medium respect to those maintained in normal medium. This result could be explained by an induction of the preOCs differentiation in the 2D co-cultures maintained in normal medium, where we recorded the highest level of TRAP and cathepsin K expression.
It has been reported that osteoblast differentiation is controlled by several transcription factors.21,22 To investigate the differentiation status of these cells in the two culture systems, we evaluated markers of the early (Runx2), middle (Osx), and late (osteocalcin) stage of osteogenesis. Runx2 and Osx are key transcription factors maximally expressed at initial and intermediate stages of osteoblast maturation, respectively. The time-modulation of Runx2 and Osx expression drives the early and the middle stages of this biologic process,23,24 but Osx plays also a key role for osteoblast to reach a full differentiation stage and, as such, can be considered a marker to monitor the occurrence of fully differentiated osteoblasts.25,26 We evaluated the expression of these transcription factors in both culture conditions. Runx2 and Osx expression reached the maximum level already after 10 days of culture on Skelite scaffolds, while in 2D cultures were maximally expressed after 40 days. These expression patterns suggested that the osteoblast differentiation process started immediately after seeding of the cells on Skelite scaffolds, but only at a later time when the cells were plated in Petri dishes. To test this hypothesis, we analyzed the expression of osteocalcin, a known marker of mature osteoblasts and bone matrix deposition. Osteocalcin expression was strongly increased in cells grown on 3D scaffolds at all times of the culture, thus confirming that the 3D mineralized environment provided by the Skelite scaffold enhanced osteoblast differentiation. Instead the observed Runx2, Osx, and osteocalcin expression patterns in the cultures performed in the Petri dishes suggested that osteogenesis remained at an early stage in 2D co-cultures.
We looked into osteoclast differentiation in both culture conditions by analyzing cathepsin K and TRAP expression. Cathepsin K is a cysteine protease that plays an essential role in the osteoclast mediated degradation of protein components of the bone matrix.27–29 TRAP is an enzyme highly expressed in osteoclasts; although it is expressed also in other cell types, it is considered a good marker to evaluate osteoclast differentiation. 30 For both genes the highest level of expression was observed in osteogenically induced 2D cultures. Expression of both genes was also observed in the osteogenically induced 3D cultures although at a significant lower level. Interestingly, a time-related increase of cathepsin K and a low level of TRAP expression were observed in the 2D co-cultures maintained in normal medium, On the contrary, a higher expression of both genes, with a trend to a progressive decrease, was observed in the cells cultured on Skelite scaffolds. In particular, in 3D co-cultures in normal medium, TRAP mRNA level was significantly higher than in the 2D cultures in the same medium after 10 and 40 days of growth (about 20- and 10-fold, respectively). It is noted that cathepsin K plays a role in the degradation of the organic component of the bone matrix, while TRAP is involved in the resorption of the mineral phase. It is tempting to speculate that the different nature of the matrix in the different culture conditions, that is, presence or absence of mineral in the ECM, could control the expression of the two genes through some feedback mechanism.
Taken together, our data on osteoblast and osteoclast differentiation in 2D and 3D co-cultures performed in normal medium suggest that the maximum induction of the osteoclastogenesis by the osteoblasts occurs at the stage when Runx2 and Osx were maximally expressed, but osteoblasts did not reach a complete differentiation and organization of the bone matrix, as indicated by the low level of osteocalcin expression. In other words, maximum osteoclastogenesis occurs at the stage when osteoblasts are already differentiated but the organization of the bone tissue is not completed yet. In our opinion, this hypothesis is confirmed by the data obtained from the 2D and 3D co-cultures performed in osteogenic medium. In fact, in 3D co-cultures maintained in this medium, where osteocalcin reached the highest level of expression and an organized bone-like ECM was observed, we recorded a decrease of the osteoclast markers respect to the 2D counterpart where the osteocalcin is expressed at low level and an organized bone-like ECM was not detectable.
It has been reported that, in mineralizing cultures of mouse calvaria osteoblasts, the RANKL mRNA expression in response to vitD3 was decreased in late-stage mature osteoblasts. 31 Thus, RANKL was preferentially expressed in immature cell and immature osteoblasts responded better to osteotropic factors in a potentially pro-osteoclastogenic manner.31,32 It has been demonstrated that the ratio between RANKL and OPG plays a key role in the regulation of bone turnover.3–7 To investigate the different cell response to an osteogenic treatment in 2D and 3D co-cultures, we evaluated RANKL and OPG expression modulation. In 3D co-cultures, where the osteoblast differentiation was enhanced, we observed an increase of osteocalcin (marker of mature osteoblasts) and OPG expression together with an inhibition of RANKL transcription. However, the inhibition of RANKL is not statistically significant (p > 0.01); thus, further investigations are necessary to confirm this pattern of expression. On the contrary, in 2D co-cultures OPG expression was inhibited and RANKL expression was enhanced. Moreover, osteocalcin mRNA level did not increase after osteogenic stimulation in this last culture condition. Taken together, these results indicate that, in the 3D model, the osteogenic stimulation drove bone turnover toward an increase of bone deposition and an inhibition of bone resorption although few mature osteoclasts were still present at the end of the culture. A different response in 2D co-cultures was observed: both bone deposition and osteoclast precursor maturation were induced. Our results are in line with the studies cited above and suggest that in bone the function of osteogenesis and osteoclastogenesis induction, in response to osteotropic factors, are performed by the same lineage of osteoblasts, but at different stages of their maturation.
Although how the dexamethasone promotes murine primary osteoblast differentiation and proliferation is still under debate,33–37 this hormone is commonly used to induce osteogenesis. In this paper we reported that a highly organized type I collagen–positive, eosinophilic bone-like ECM was produced and its deposition increased over the period of culture in the 3D Skelite scaffolds cultured in a dexamethasone-containing osteogenic medium. Moreover, the presence of dexamethasone, ascorbic acid, and β-glycerophosphate in the culture medium led, as expected, to the deposition of a more organized ECM compared to that obtained by the cultures maintained in normal medium.
A good in vitro model of bone should faithfully mimic the morphological features of bone tissue. The model here reported, as demonstrated by the histological analysis of the deposited ECM, reproduces a highly organized bone environment. To our knowledge, in most of the 3D in vitro culture models of bone cells that have been reported in the literature, the analyses focused on the expression of specific genes and on calcium deposition, but no clear evidence of an in vitro bone tissue formation has been reported.
In conclusion, the 3D model of bone we developed represents a useful model to investigate cellular and molecular mechanism of bone turnover, including preclinical tests on new drugs. Our data indicate that the 3D environment enhances osteoblast differentiation that in turn promotes earlier osteoclast differentiation. Moreover, we reported the in vitro formation of a well-organized bone matrix that is not possible to obtain with traditional 2D co-cultures.
Footnotes
Acknowledgments
Supported by funds from the Italian (ASI-OSMA) and the European (ESA-ERISTO) Space Agencies. Support was also received from the Centro Studi Termali Veneto, Pietro D'Abano di Abano Terme e Montegrotto Terme.
The technical contribution of Raffaela Arbicò and Roberta Biticchi for the histology analysis and the mice care is acknowledged.
Disclosure Statement
No competing financial interests exist.