
Abstract
In medicine, N-methyl pyrrolidone (NMP) has a long track record as a constituent in medical devices approved by the Food and Drug Administration and thus can be considered as a safe and biologically inactive small chemical. In the present study, we report on the newly discovered pharmaceutical property of NMP in enhancing bone regeneration in a rabbit calvarial defect model in vivo. At the cellular level, the pharmaceutical effect of NMP was confirmed, in particular, in combination with bone morphogenetic protein (BMP)-2, because NMP increased early and late markers for maturation of preosteoblasts and human bone marrow–derived stem cells in vitro. When we used the multipotent cell line C2C12 without autologous BMP expression, NMP alone had no effect on alkaline phosphatase activity, a marker for osteogenic transdifferentiation. Nevertheless, in combination with low BMP-2 doses, alkaline phosphatase activity was more than eight times as great. Thus, the pharmaceutical NMP mode of action is that of an enhancer of BMP activity. The dependency of the effects of NMP on BMP was confirmed in preosteoblasts because noggin, an extracellular BMP inhibitor, suppressed NMP-induced increases in early markers for osteoblast maturation in vitro. At the molecular level, NMP was shown to have no effect on the binding of BMP-2 to the ectodomain of the high-affinity BMP receptor IA. However, NMP further increased the phosphorylation of p38 and Smad1,5,8 induced by BMP-2. Thus, the small chemical NMP enhances BMP activity by increasing the kinase activity of the BMP receptor complex for Smad1,5,8 and p38 and could be employed as a potent drug for bone tissue regeneration and engineering.
Introduction
Locally secreted BMP antagonists regulate extracellular BMP binding to its receptor and BMP signaling. One of the best-characterized BMP antagonists is noggin, which was identified as a dorsalizing factor produced by Spemann's organizer during early embryogenesis. Noggin exerts its bioactivity through high-affinity binding to extracellular BMPs and blocking their binding to BMP receptors.7,8
With the cloning, expression, and mass production of recombinant human BMPs (rhBMPs), powerful growth and differentiation factors toward the osteogenic lineage had been developed up to the level of clinical application. However, the first clinical trials showed that milligram doses of BMP-2 were required. 9 Therefore, BMP presentation and release kinetics, mainly determined by the delivery system, have to be optimized for efficient clinical application of BMPs.10–12
Another strategy to decrease the BMP dose in clinical applications is to combine the cytokine with enhancers of BMP activity. Ideally, an enhancer should be a small molecule, easily applicable to the carrier materials and able to increase the activity of the therapeutic protein. In the future, enhancers might become novel tools to tune therapeutic molecules such as BMPs. In conjunction with bone tissue engineering, such enhancers could reduce or substitute for the clinical application of BMPs.
Recently, we reported on the application of BMP-2 in conjunction with guided bone regeneration (GBR). 13 GBR is a clinically well-documented and successful procedure in dental implantology. 14 It is based on the placement of a membrane over the defect to create a compartment where bone can regenerate without competition from other, faster-growing tissues. Here we report on the unexpected finding that a N-methyl pyrrolidone (NMP)-releasing GBR membrane induced an acceleration of bone regeneration in a rabbit calvarial defect model. Because this newly discovered pharmaceutical aspect of NMP to serve as an accelerator of bone regeneration could prove useful for bone repair and bone tissue engineering, we started to characterize the bioactivity of NMP in more detail.
Methods
Reagents, guided bone regeneration membranes, and antibodies
NMP was purchased from Sigma-Aldrich (St. Louis, MO). The biodegradable woven flexible guided bone regeneration membrane OsseoQuest was purchased from Gore (Saint Guenault, France), and the solid rigid InionGTR membrane and NMP were obtained from Inion OY (Tampere, Finland). Polyclonal anti-p38, anti-α-actinin, anti-ERK1/2, anti-pERK1/2, anti-pp38, and anti-pSmad1/5/8 were obtained from Cell Signaling Technology (Danvers, MA).
Animal testing for GBR
We generated a GBR model in rabbits by drilling four 6 mm-diameter defects into the calvarial bones and placed membranes on each side of the defect to mimic a GBR situation. Membranes were used as recommended by the manufacturer. The InionGTR membrane was used as solid rigid membrane without NMP pretreatment or as a solid flexible membrane after NMP treatment. In brief, the solid, rigid InionGTR membranes were submerged in NMP for 30 s, air dried for 15 min, and then washed in phosphate buffered saline (PBS) to remove NMP from the outside of the membrane. During this procedure, the initially rigid membrane became flexible. 15 Four weeks after the procedure the animals were sacrificed, and the samples were embedded for histology and analyzed for bone regeneration, as previously reported. 16 The ethics committees of the Canton of Zurich and the University of Zurich approved all studies.
Cell cultures
C2C12 and MC3T3-E1 clone 4 were purchased from American Type Culture Collection (ATCC, Manassas, VA) through its European distributor European Collection of Animal Cell Cultures (Salisbury, United Kingdom). The osteoblastic cell line MC3T3-E1 was established from a C57BL/6 mouse calvaria and selected on the basis of high alkaline phosphatase (ALP) activity in the resting state. These cells have the capacity to differentiate into osteoblasts and osteocytes and have been shown to form calcified bone tissue in vitro. C2C12 cells are pluripotent myogenic cells able to transdifferentiate into a variety of tissues such as bone and adipose tissue. Therefore, they are often used as a model for mesenchymal stem cells. 17 Cells were cultured in Dulbecco's modified Eagle medium (DMEM) (C2C12) or alpha-minimal essential medium (MC3T3-E1) supplemented with 10% fetal calf serum and antibiotics (100 U/mL penicillin G and 100 mg/mL streptomycin). Cells were trypsinized and passaged every 2 to 3 days, and cultures were never allowed to become confluent. All cells were grown at 37°C in humidified air mixed with 5% carbon dioxide.
Human bone marrow–derived stromal cells
The human bone marrow–derived stromal cells were a gift from Dr. Lorenz Meinel (ETH, Zurich, Switzerland). They had been isolated using density gradient centrifugation from whole bone marrow (25-cm2 harvests) obtained from Clonetics (Santa Rosa, CA) as described previously. 18 Briefly, bone marrow suspensions were overlaid onto a polysucrose gradient (δ = 1.077 g/cm3) and centrifuged at 800 g for 30 min at room temperature. After lysis of the remaining red blood cells, the cell layer was pelleted, resuspended in expansion medium (DMEM, 10% fetal bovine serum, 1 ng/mL basic fibroblast growth factor), and seeded at a density of 5 × 104 cells/cm2. The adherent cells were allowed to reach 80% confluence. The second-passage cells were frozen and used for experiments. All cell batches were characterized with respect to expression of specific cell markers 18 and for their ability to differentiate into chondrogenic and osteogenic lineages. Osteogenesis was studied in expansion medium supplemented with 50 μg/mL of ascorbic acid and 7 mM ß-glycerolphosphate (Sigma-Aldrich).
ALP activity assay
ALP activity was measured as a marker of osteoblastic differentiation in MC3T3-E1 and C2C12 cells. Cells were seeded at a density of 5 × 104 cells/cm2 in 6- or 24-well plates (n = 3 per group). Treatment with NMP, BMP-2, and noggin diluted in the tissue culture medium started 5 h after seeding, and incubation was continued for 6 more days. rhBMP-2 used in this study is produced in our laboratory 19 to a quality that allows its clinical application. 13 Noggin was a gift of Peter Hortschansky (Jena, Germany) and was applied at 1 μg/mL. 20 After 6 days of incubation, medium was removed, and cells were washed with PBS and then scraped in 0.56 M 2-amino-2-methyl-1-propanol, pH 10.5. The pellets were then homogenized for 10 s. After centrifugation, the supernatant was collected and tested for ALP activity using p-nitrophenyl phosphate (Sigma-Aldrich) as a substrate. 20 The protein content of the lysates was determined using Bradford protein assay reagent (Bio-Rad Laboratories; Hercules, CA). Experiments were performed independently in triplicate.
Calcium deposition by preosteoblastic cells
MC3T3-E1 cells were maintained in alpha-modified minimum essential medium (Life Technologies, Inc., Grand Island, NY) containing 10% fetal calf serum (Life Technologies, Inc.), and 50 μg/mL gentamicin was plated at a density of 30,000 cells/cm2 in 6-well plates. Starting at 80% confluence (normally day 3 after plating), the culture medium was supplemented with 50 μg/mL of ascorbic acid, 10 mM of β-glycerophosphate, and the different molecules. Medium change and NMP and BMP-2 additions were performed once a week, and calcium deposition was determined after 4 weeks in culture. For Alizarin red staining of calcium, cultured MC3T3 or human bone marrow–derived stromal cells were washed once with PBS and fixed for 1 h in 70% ethyl alcohol. After the cells were rinsed with water, 4 mL of an Alizarin red solution (40 mM, pH 4.2) was added, and the cells were stained for 10 min. Non-specific staining was removed using several washes with water, and the stained wells were photographed.
Quantitative reverse transcription polymerase chain reaction
Messenger RNA (mRNA) was isolated from MC3T3 cells after 6 days of stimulation using the RNeasy Mini Kit (Qiagen, Hilden, Germany). After performing the reverse transcription (RT) reaction (first-strand complementary DNA (cDNA) Synthesis Kit for RT-polymerase chain reaction (PCR), Roche, Mannheim, Germany), first-strand cDNA samples were quantified using serial dilutions in a Light Cycler system (LightCycler 2.0 Real-Time PCR System, Roche, Mannheim, Germany) using SYBR green (Fast start DNA master SYBR Green 1, Roche) and normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and control samples, respectively. The conditions for the 42 cycles were denaturation for 15 s at 95°C and annealing for 10 s at 58°C for GAPDH, 60°C for osterix, and 61° for osteocalcin, followed by extension for 20 s at 72°C. The following primers were used: osterix forward: 5′-CCCCTGCTCCTTCTAGGC-3′, osterix reverse: 5′-CCGTCAACGACGTTATGC-3′, osteocalcin forward: 5′-ACAAGTCCCACACAGC-AGCTT-3′, osteocalcin reverse: 5′-GCCGGAGTCTGTTCACTACCT-3′, GAPDH forward: 5′- AACGACCCCTTCATTGAC-3′, GAPDH reverse: 5′-TCCACGACATACTCAGCA-3′.
Western blotting analysis
MC3T3-E1 cells treated with the different compounds were rapidly frozen in liquid nitrogen and stored at −80°C for further analysis. In experiments aimed at testing for the effect of NMP on p38, cells were made quiescent for stress and MAPK by switching to 1% fetal calf serum 24 h before and during the study. 21 Cells were lysed at 4°C in lysis buffer containing 50 mM Tris (pH 7.4), 150 mM sodium chloride (NaCl), and 1 mM phenylmethyl sulfonyl fluoride supplemented with a protease inhibitor (Protease Inhibitor Cocktail Set III, Calbiochem, Gibbstown NJ), 1% Nonidet P-40, 1% sodium deoxycholate, and 0.1% (w/v) sodium dodecyl sulphate (SDS) for 10 min. Lysates were then cleared by centrifugation at 6000 × g for 30 min. Proteins were fractionated onto a 12% SDS polyacrylamide gel electrophoresis gel, transferred to Immobilon P membranes (Millipore, Zug, Switzerland), and immunoblotted with specific antibodies.22,23 Detection was performed using peroxidase-coupled secondary antibody, enhanced chemiluminescence reaction (Amersham ECL Western Blotting Detection Reagents, GE Healthcare Europe GmbH, Otelfingen, Switzerland), and visualization using autoradiography. Membranes that were reprobed had been stripped in stripping buffer (62.5 mM Tris-hydrochloric acid, pH 6.8, 2% (w/v) SDS, 100 mM β-mercaptoethanol) according to the manufacturer's protocol (Millipore).
Biosensor interaction analysis
The BIA2000 system (BIAcore, GE Healthcare, Europe, Otelfingen, Switzerland) was used to record the binding of BMP-2 to immobilized receptor ectodomains at different NMP concentrations. The biotinylated receptor proteins were fixed separately to a streptavidin-coated matrix of biosensor CM5 in flow cells 2, 3, and 4 at a density of 190 resonance units (RUs) corresponding to 190 pg of protein (∼14.5fmol of receptor) per mm2, 0.23 to 7.2 μg/mL of BMP-2 (6.25-200 nM) in 10 mM of N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid pH 7.4, 500 mM NaCl, 3.4 mM ethylenediaminetetraacetic acid, and 0.005% surfactant P20 were perfused over all four flow cells in series at a flow rate of 10 μL/min at 25°C, and sensograms were recorded at a data sampling rate of 2.5 Hz. The association period was 5 min, and the dissociation period was set to 3 min. Free receptors were regenerated by perfusion with 0.1 M acetic acid and 1 M NaCl for 2 min. A background sensogram (streptavidin control) was subtracted from the sensograms obtained in the flow cells with attached BMPR IA. The different sensograms were evaluated according the fitting routine 2 provided by BIA evaluation 2.2.4 software (BIAcore). The equilibrium binding of rhBMP-2 to the ectodomain of BMPR IA at steady state was determined in triplicate at six different concentrations of BMP-2 to obtain KD values.
Statistical analysis
SPSS 10 (SPSS, Inc., Chicago, IL) was used to calculate the mean values, standard deviations, and standard errors of the measurement (SEMs). Unless otherwise indicated, standard deviations are given. For statistical analysis, the Student t-test was applied after a normal deviation was confirmed using the Kolmogorov-Smirnov and Shapiro-Wilks test.
Results
The presence of NMP in a biodegradable GBR membrane accelerates bone regeneration in vivo
In the course of the development of a biodegradable GBR membrane, we performed an in vivo study to screen several membrane prototypes. The drawback in using biodegradable polylactide- and polyglycolide-based membranes, such as the InionGTR membrane, is their inherent rigidity, which compromises their applicability in the clinic. One strategy to overcome this problem was to use NMP as a plasticizer to generate more-flexible membranes. 15 In the initial screening, the best-performing membranes all contained NMP. To further investigate this finding, we performed another animal study using eight rabbits (Fig. 1), comparing bone regeneration in untreated defects with regeneration in defects treated with a solid polylactide- or polyglycolide-based membrane with and without NMP (InionGTR) and a woven polylactide- or polyglycolide-based membrane (OsseoQuest). The latter was included as a biodegradable flexible control membrane to discriminate the plasticizing aspect of NMP from its pharmaceutical one. When left untreated, bone regeneration within the model system was 28.4 ± 3.2% (SEM), whereas the application of a flexible woven membrane (OsseoQuest) increased bone regeneration to 57.0 ± 9.2% (SEM), and application of solid rigid membranes yielded 49.3 ± 8.7% (SEM) bone regeneration. The difference in bone regeneration facilitated by the flexible woven and rigid solid membrane was not significant (p = 0.46), suggesting that an increase in flexibility of the membrane had no positive effect on bone regeneration in this model system. When the same solid membrane was loaded with NMP, bone regeneration increased to 79. 27 ±5.6% (SEM) (Fig. 1). The increase in bone regeneration was significantly greater than with the untreated rigid solid (p <0.02; n = 8), or flexible woven membrane (p < 0.02; n = 8), suggesting that NMP not only serves as a softener of the rigid solid membrane, but also enhances bone regeneration.
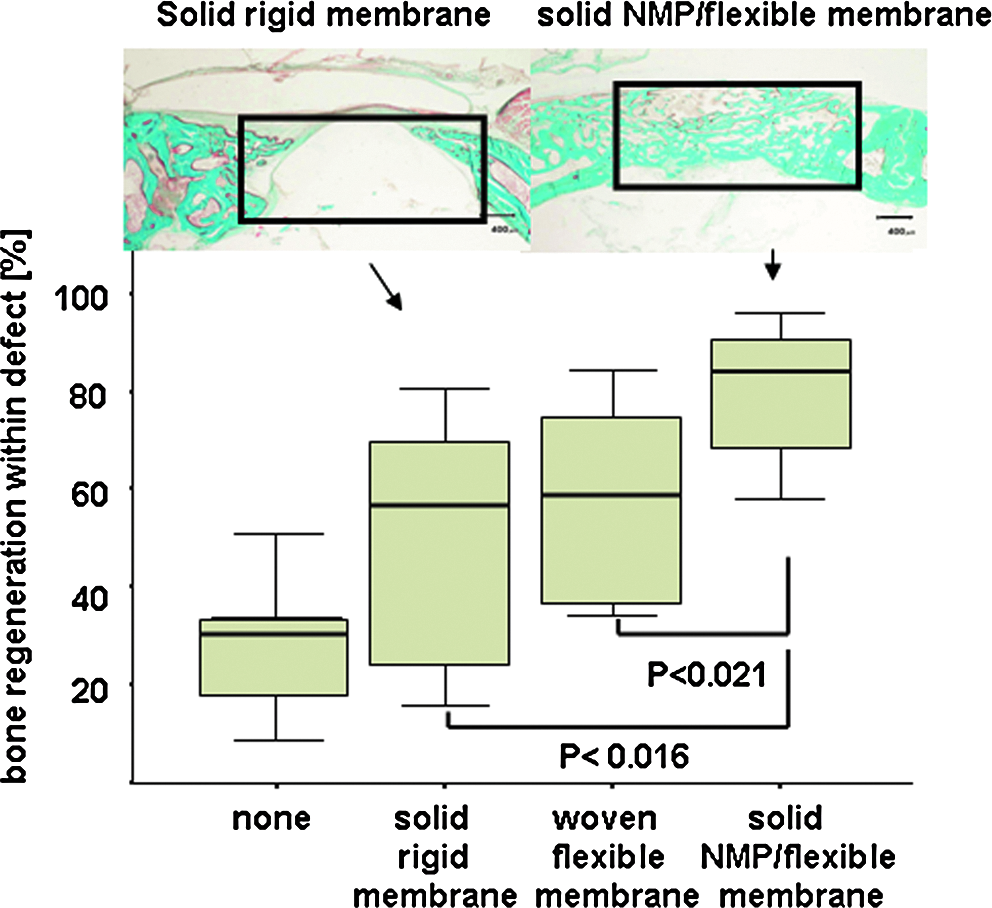
N-methyl pyrrolidone (NMP) enhances bone regeneration in vivo. Subcritical-size defects were generated in calvarial bones of eight rabbits and treated with a solid rigid membrane, a woven flexible membrane, or a solid NMP flexible membrane or left untreated. After 4 weeks, bone regeneration within the defect was evaluated based on the middle cross-sections (upper panel, defect area is indicated, scale of 400 μm is indicated). Histomorphometrical analysis (lower panel) revealed significantly greater bone regeneration when the solid membrane contained NMP than the same membrane without NMP (p = 0.02). The NMP-induced increase in flexibility of the solid membrane had no influence on bone regeneration in this model system, because the solid rigid and the woven flexible membranes performed equally well. Histomorphometrical results are displayed (lower panel) as box-plots ranging from the 25th (lower quartile) to the 75th (upper quartile) percentile, including the median and whiskers showing the minimum and maximum values. The Goldner Trichrome–treated sections stain bone green (upper panel).
In preosteoblasts, NMP increases early markers for bone formation in a concentration-dependent fashion, and this effect requires autologous BMP
To study the biological effect of NMP in more detail, we used the MC3T3-E1 cell line as a model for preosteoblastic cells in vitro. In bone, autologous BMP is deposited in the extracellular matrix. In vitro culture of MC3T3-E1 preosteoblastic cells can be used to model the situation in vivo, because they mature to osteoblasts dependent on BMPs, which they secrete and deposit into the extracellular matrix. When we subjected MC3T3-E1 cells to NMP, ALP activity, an early maker for bone formation (Fig. 2A), showed a concentration-dependent increase.

Enhancing effect of NMP on the maturation of preosteoblastic cells. (
To test whether the bioactivity of NMP is dependent on autologous BMP, we employed the BMP-antagonist noggin to bind and neutralize autologous BMP.7,24 Upon noggin application, ALP activity decreased to 51.8 ± 6.4% of the control level and could not be restored by NMP application (64.7 ± 11.9%; p < 0.12; n = 9). Thus, the effect of NMP on preosteoblasts depends on the presence of autologous BMP.
Effect of combinations of NMP and BMP-2 on preosteoblastic cells
To further investigate the interplay between NMP and BMP in bone formation and repair in greater detail, preosteoblastic cells were treated with combinations of NMP and BMP-2. At 1 μg/mL of BMP-2 and increasing concentrations of NMP, ALP activity increase was much greater than with BMP-2 or NMP alone (Fig. 2A). A late marker for osteoblast maturation is the mineralization of the extracellular matrix. When we exposed preosteoblasts to increasing concentrations of NMP, calcium deposition was not affected (Fig. 2B). In combination with 1 μg/mL of BMP-2, however, rising NMP concentrations (Fig. 2B) resulted in markedly greater calcium deposition than BMP-2 or NMP treatment alone. Thus, NMP has an enhancing effect with BMP-2 treatment on early and late markers of osteoblast maturation. Next we performed real-time quantitative PCR to determine mRNA levels of markers of bone formation in preosteoblastic cells. Osteocalcin mRNA, a late marker for bone formation, was 19.8 ± 0.2 times as great in preosteoblasts treated with 5 mM of NMP and 1 μg/mL of BMP-2, 1.3 ± 0.1- times as great with 5 mM of NMP, and 14.7 ± 1.1 times as great with 1 μg/mL of BMP-2 as with control untreated cells (Fig. 2C). The enhancing effect was also detectable for mRNA content of the late bone-specific transcription factor osterix II. 25 mRNA levels of osterix were 7.7 ± 0.4 times as great in preosteoblasts treated with 5 mM of NMP and 1 μg/mL of BMP-2, 1.4 ± 0.1 times as great with 5 mM of NMP, and 4.3 ± 0.2 times as great with 1 μg/mL of BMP-2 as with control untreated cells (Fig. 2D).
NMP is an enhancer of BMP-2 activity
To discriminate between enhancing and synergistic effect of combinations of NMP and BMP-2, we used the pluripotent mesenchymal cell line C2C12. Unlike MC3T3-E1 cells, C2C12 cells do not express BMPs. However, exposure of C2C12 cells to BMP is known to convert these cells from the myoblastic to the osteoblastic lineage. The conversion in response to BMP treatment can be monitored according to the BMP concentration-dependent increase in ALP activity. 26 In the absence of NMP, an increase in ALP activity was induced starting at a BMP-2 concentration of 100 ng/mL of BMP-2 (Fig. 3A). In the absence of BMP, increasing NMP concentrations ranging from 1 to 5 mM had no effect on ALP activity. At 50 ng/mL of BMP-2, ALP activity was not greater than with untreated cells. However, in combination with 5 mM of NMP, ALP activity was significantly greater (p = 0.04). This was even more pronounced at higher BMP-2 concentrations (Fig. 3B). The maximum enhancing effect of 5 mM of NMP was seen at 100 ng/mL of BMP-2—8.4 ± 1.5 times as great, declining to 1.9 ± 1.5 times as great at 800 ng/mL of BMP-2, and remaining in this range up to a BMP-2 concentration of 1200 ng/mL. To show the clinical relevance of our finding, we tested the enhancing effect of 5 mM of NMP and 100 ng/mL of BMP-2 on human bone marrow–derived stromal cells. When we exposed primary human bone marrow stromal cells to 100 ng/mL of BMP or 5 mM of NMP alone (Fig. 4A, B), calcium deposition was not induced. However, in the presence of 5 mM of NMP, the activity of 100 ng/mL of BMP-2 was enhanced to an extent that nodule formation occurred (Fig. 4A, B), and calcium deposition was significantly greater (p < 0.001) (Fig. 4C).

NMP is an enhancer of BMP activity. (
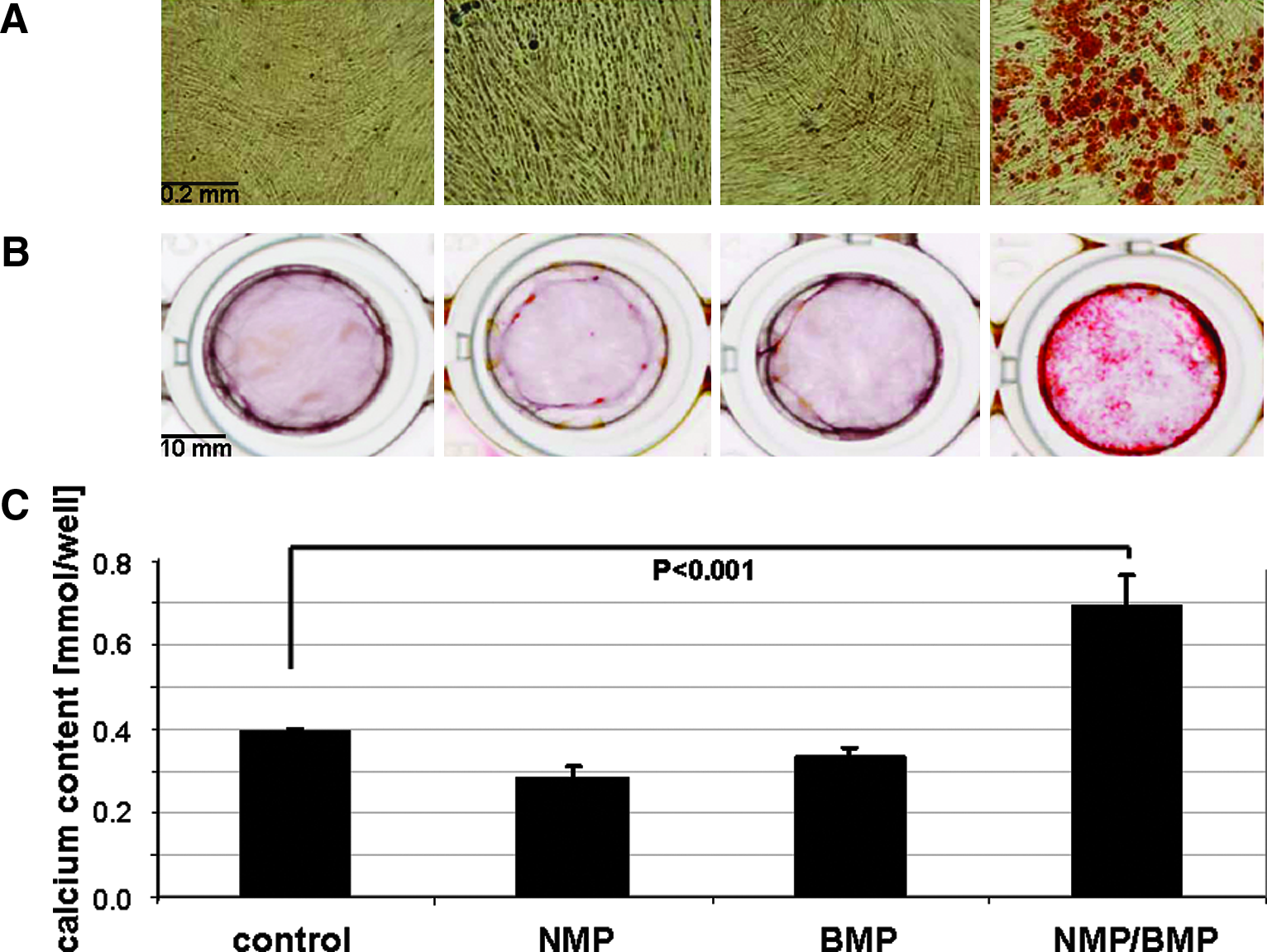
Effect of a low BMP dosage and NMP and their combination on human bone marrow–derived stromal cells. Human bone marrow–derived stromal cells were exposed to 5 mM of NMP, 100 ng/mL of BMP-2, or the combination thereof. After 5 weeks of exposure, the cells were stained with Alizarin-red. (
NMP enhances the kinase activity of the BMP receptor complex for Smads and p38
To explore the mechanisms by which NMP enhances BMP-2 activity, we first studied the effect of NMP on BMP-2 binding to the extracellular portion of BMPR IA. The BMPR IA high-affinity receptor for BMP was chosen because type II receptors make only a minor contribution to the affinity of BMP-2 for the receptor on whole cells. 2 The binding of BMP-2 to the ectodomain of the recombinant BMPR IA with varying concentrations of NMP was analyzed using biosensor interaction analysis. 27 With the NMP concentrations tested (0–100 mM), no change in the affinity of BMP-2 to the BMPR IA ectodomain was observed (Fig. 5A). The apparent KD value with increasing BMP-2 concentrations (6.25 to 200 nM) was 1.67 nM in the absence and 1.63 nM in the presence of 50 mM of NMP (Fig. 5B).
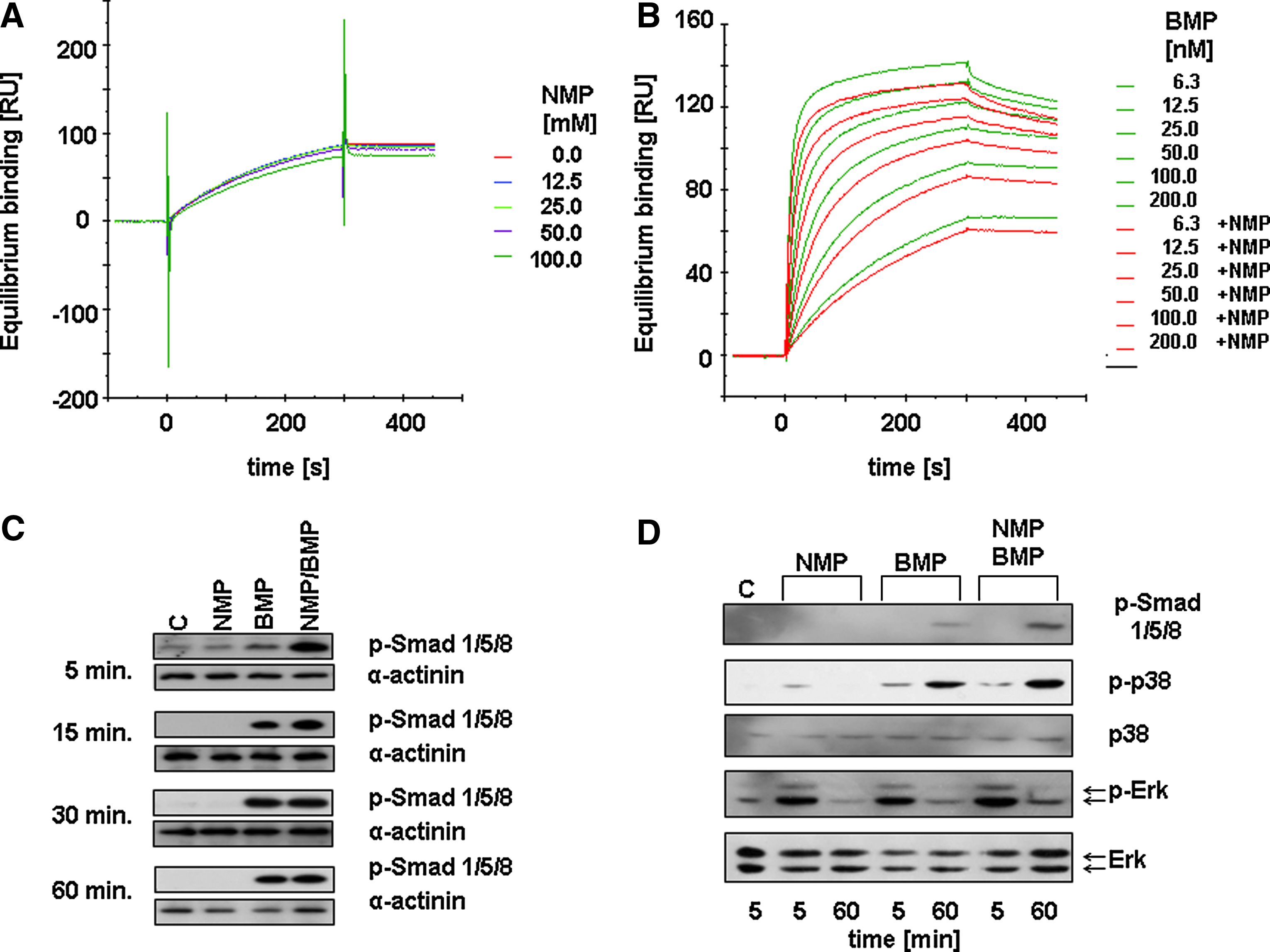
Effect of NMP on BMP signaling. (
Next, we examined the effect of NMP on the kinase activity of the intracellular portion of BMPR I in preosteoblastic cells. In the first 15 min after application of 5 mM of NMP and 1 μg/mL of BMP-2, the phosphorylation of BMPR-specific Smad proteins was induced (Fig. 5C). However, after 15 min, any further increase was undetectable. In addition to Smad phosphorylation, BMP also signals via p38 phosphorylation, 4 which can only be studied at low serum concentrations. When the cells were treated under low serum conditions with 5 mM of NMP and 1 μg/mL of BMP-2, Smad and p38 phosphorylation increased after 1 h, suggesting a longer-lasting effect of NMP on BMP signaling (Fig. 5D).
Discussion
The starting point of this study was the discovery that NMP, initially used as plasticizer for a rigid, solid polylactide- or polyglycolide-based membrane, also enhances bone regeneration by a factor of two in a calvarial bone regeneration model in vivo (Fig. 1). Following this discovery of a potential pharmaceutical effect of NMP made in an in vivo model system with relevance to the clinical situation, we started to characterize the bioactivity of NMP in greater detail.
The enhancing effect of NMP suggested by the calvarial defect model was confirmed in preosteoblastic cells of murine origin (MC3T3-E1), where NMP mediated a concentration-dependent increase in ALP activity, an early bone marker (Fig. 2A). Next we showed that NMP action needs endogenous BMP, because addition of the BMP-binding and -neutralizing antagonist noggin reduced the effect of NMP substantially. Next we looked at the effect of combinations of NMP and BMP-2. In the preosteoblastic cell line MC3T3-E1, NMP enhanced BMP activity, as illustrated by higher ALP activity, mineral deposition, osteocalcin transcription, and osterix transcription levels above the BMP only values (Fig. 2). Therefore, the effect of NMP on preosteoblasts can be linked to the BMP pathway.
To discriminate between enhancing and synergistic effects of NMP and BMP-2 (as reviewed in 28 ), we employed the multipotent C2C12 cells, which lack autologous BMP expression. The results showed that NMP alone is not osteogenic, because it is insufficient to direct C2C12 cells toward the osteoblastic lineage. In combination with BMP-2, however, NMP markedly enhanced the osteogenic effect of BMP-2 determined by the increase in ALP activity (Fig. 3B). Even BMP-2 concentrations as low as 50 ng/mL, normally not capable of converting the fate of C2C12 cells into the osteogenic lineage, are sufficient to increase ALP activity when applied in combination with 5 mM of NMP. To show the clinical relevance of our finding, a low BMP-2 concentration of 100 ng/mL was chosen and applied to human bone marrow–derived stromal cells. In the absence of 5 mM of NMP, 100 ng/mL of BMP-2 did not induce mineral deposition, but in combination with 5 mM of NMP, mineral deposition was induced. Therefore, the enhancing effect of NMP at low BMP-2 concentrations is also evident with human bone marrow–derived stromal cells (Fig. 4), which is a clinically used source for multipotent adult mesenchymal stem cells. 29 In essence, adult human mesenchymal stem cells exposed to suboptimal BMP-2 dosages are guided toward the osteogenic lineage in the presence of NMP. Thus, NMP is an enhancer of the osteogenic effect of BMP-2, and its application in combination with BMP-2 could potentially decrease BMP-2 dosages needed for bone regeneration and bone tissue engineering approaches in medicine. To make this vision come true, additional studies and the development of an optimal dual delivery device for BMP-2 and NMP are needed. For smaller bone defects, normally not treated with BMPs, the enhancing effect of NMP on the autologous BMP deposited in bone is capable of accelerating bone healing, as illustrated in vivo in the calvarial defect model (Fig. 1). Therefore, NMP alone is suitable for enhancing bone regeneration at orthotopic sites.
At the molecular level, a BMP receptor complex is needed for BMP signaling. Extracellular BMP binds with high affinity to BMPR I, which contributes the majority to the affinity of BMP-2 for the receptor on whole cells. 2 Our results show that NMP does not affect the binding of BMP-2 to the ectodomain of BMPR IA (Fig. 5A, B). However, in the evanescent field–based BIAcore system, only the binding of single BMP-2 dimers to single BMPR IA ectodomains were determined, leaving out possible contributions of the second BMPR IA or the two BMPR II receptor ectodomains to the binding affinity. Therefore, an effect of NMP on the binding of BMP-2 to the whole extracellular part of the BMP receptor complex can not be totally excluded, because in vivo it involves a complex composed of two BMPR I molecules, two BMPR II molecules, and one BMP-2 dimer. Another open question is the biological effect of metabolites of NMP on receptor binding. In humans and rats, the main metabolite of orally administered NMP is 5-hydroxy-N-methylpyrrolidone, and it is excreted in urine. 30 In humans, a major metabolic pathway from 5-hydroxy-N-methylpyrrolidone to N-methyl succinimide to 2-hydroxy-N-methylsuccinimide has been suggested. 31 However, a biological effect of any of these metabolites has not been reported.
The elements of the BMP pathway just downstream of the BMP receptors are Smads and p38.32,33 The combined application of BMP-2 and NMP substantially increased Smad phosphorylation 5 min after NMP application (Fig. 5C), and the effect continued for more than 1 h at low serum concentration for Smad and p38 phosphorylation (Fig. 5D). This series of experiments supports the enhancing mode of NMP on BMP-2 activity, because NMP alone has a limited effect on the pathways involved in BMP signaling, but in combination with BMP-2, Smad phosphorylation is enhanced within minutes. Thus, NMP enhances the kinase activity of BMPR I in the BMP receptor complex, which is responsible for Smad and p38 phosphorylation in a way that does not involve protein synthesis.
The most important modulator of BMP initial signaling is BMPR II, 34 with its extended intracellular domain, known to interact with a variety of proteins comprising MAPKs and protein kinase C. 6 Because the affinity of BMP-2 to the high-affinity ectodomain of the receptor is NMP independent (Fig. 5A, B), but the initial BMP signal for Smad and p38 phosphorylation is greater under the influence of NMP, these data suggest that NMP probably acts at the level of the intracellular portion of the BMP receptor complex. Intracellularly, the receptor complex is associated with several proteins, 6 including c-kit 5 and probably additional, unrecognized elements. One could speculate that the enhancing effect of NMP on BMP activity derives from a direct binding of NMP to the receptor or receptor-associated proteins, as has been shown for ethanol, 35 or by interaction of NMP with membranes due to the plasticizing potential of NMP. The latter could perturb the curvature of membranes, receptor-associated lipid rafts, or the spatial arrangement of membrane proteins and, by doing so, enhancing the kinase activity of BMPR I for Smads and p38. Because of the complexity of these possible effects, additional work is needed to pinpoint the direct target of NMP in the BMP receptor complex or its membranous environment.
More than 50,000 metric tons of NMP are produced and used per year, mainly for cleaning purposes but also for plastic engineering and Food and Drug Administration–approved medical devices. Despite this broad use of NMP, nothing is known about its enhancing effect on the BMP pathway, essential during early and late developmental stages but also later in life. Thus, our results highlight the need for a better understanding, beyond toxic and carcinogenic aspects, of the effect of industrial chemicals on biological systems. In the case of NMP, a thorough investigation identified a novel, locally applicable drug for bone repair, accelerated bone regeneration, or potentially even bone tissue engineering applications in combination with rhBMP. In the clinic, NMP could prove useful as single drug for the treatment of small bone defects. In combination with rhBMPs, NMP could serve as enhancer of BMP activity to treat critical size defects with much lower, and thus safer, and less-expensive BMP dosages.
Footnotes
Acknowledgments
The authors would like to thank Inion OY, Tampere, Finland, for the support of M.E. and B.SM. This research work was partly supported by a grant from the AO Foundation (Davos, Switzerland) through the Biotechnology Advisory Board and the Swiss National Funds (310000-116240). We also would like to acknowledge the technical assistance of Alexandr Tchouboukov, Yvonne Bloemhard, and Flora Nichols.
Disclosure Statement
No competing financial interests exist.