
Abstract
Collagen is the main protein component of the extracellular matrix of bone, and it has structural and instructive properties. Collagen undergoes many post-translational modifications, including extensive crosslinking. Although defective crosslinking has been implicated in human syndromes (e.g., osteogenesis imperfecta or Ehlers-Danlos syndrome), it is not clear to what extent crosslinking is necessary for collagen's instructive properties during bone formation. Here we report that inhibition of collagen crosslinking in the mouse pre-osteoblast cell line MC3T3-E1 impairs the osteogenic program. Genome-wide expression profiling of beta-aminopropionitrile–treated and control cells revealed that matrix deposition by MC3T3-E1 cells provides a feed back signal, driving cells through the differentiation process, that is strongly impaired when crosslinking is inhibited. Inhibition of crosslinking did not affect osteogenic differentiation of human mesenchymal stem cells (hMSCs), shown by the expression of alkaline phosphatase and genome-wide gene expression analysis, although it enhances matrix mineralization. In conclusion, collagen crosslinking harbors instructive properties in MC3T3-E1 differentiation but plays a more-passive role in differentiation of bone marrow-derived hMSCs.
Introduction
ECM also plays a critical role in osteogenic differentiation. MC3T3-E1 cells is a murine pre-osteoblastic cell line in which the osteogenic program is regarded as a matrix-driven process. Proliferation and differentiation of MC3T3-E1 cells into osteoblasts is influenced by matrix stiffness and it is critically dependent on the presence of ECM.12,13 For instance, in the absence of ascorbic acid, collagen type I protein production is impaired in MC3T3-E1 cells, resulting in inhibition of osteogenic differentiation. Moreover, treatment of the cells with an inhibitor of collagen synthesis or with antibodies against α2β1 integrins, the transmembrane receptors mediating collagen–cell interaction, results in abolishment of the osteogenic program.13,14
The instructive property of ECM may be an interesting tool to direct the osteogenic differentiation program of human mesenchymal stem cells (hMSCs) for bone tissue engineering applications. hMSCs are bone marrow–derived cells that were isolated for the first time because of their ability to adhere to tissue culture plates. 15 hMSCs, also referred to as colony forming unit fibroblasts, marrow stromal fibroblasts, bone marrow–derived fibroblasts, mesenchymal progenitor cells, and bone marrow–derived stromal cells, are able to differentiate into several different lineages, including the chondrogenic, adipogenic, myogenic, and osteogenic lineages.16–21
During osteogenic differentiation of hMSCs, the uncommitted multipotent cell differentiates into a functional mature osteoblast, which secretes a matrix that will later be mineralized. The differentiation of hMSCs into the osteogenic lineage is a stepwise process in which different signalling pathways (e.g., transforming growth factor beta and Wnt signalling) and nuclear receptors (e.g., glucocorticoid and vitamin D3) play a complementary role. 22 Several soluble factors, such as dexamethasone, cyclic adenosine monophosphate (cAMP), bone morphogenetic protein 2 (BMP-2), and vitamin D3, have the potential to induce hMSCs into the osteogenic lineage. 22 Exposure of hMSCs to these compounds is mainly limited to the in vitro expansion phase because in vivo–controlled release of small molecules is far from trivial. An alternative strategy relies on the use of natural or synthetic ECM proteins to induce differentiation into the desired lineage. Proteins can be combined with scaffold materials, and proof of principle has been shown in the literature for this approach. For example, when cells were cultured in a collagen type I matrix with a controlled stiffness, they irreversibly differentiated into the osteogenic lineage. 3 In addition, hMSC differentiation into the osteogenic lineage was enhanced when the cells were exposed to ECM proteins such as vitronectin, laminin-5, and collagen type I.6,23,24 This effect might be mediated through ECM–focal adhesion kinase (FAK)–extracellular signal-related kinase 1/2 (ERK) activation.25,26 Thus, ECM proteins represent a potential pro-osteogenic stimulus for bone tissue engineering.
It is not only the presence of collagen, but also the type and structure that control the function of ECM. Examples in which the structure of collagen affects bone are patients suffering from Bruck syndrome and OI. In the first case, deficiency of bone-specific telopeptide lysyl hydroxylase results in aberrant crosslinking of bone collagen, leading to osteoporosis, joint contractures, fragile bones, and short stature. 27 In the case of OI, mutations affecting the structure or the abundance of collagen type I can be found in the majority of patients, resulting in bone abnormalities ranging from bone fragility to high bone mineralization.28–31 Moreover, when hMSCs were proliferated on denatured collagen, they showed greater retention of osteogenic potential than expansion on tissue culture plates. 32
Structural changes in the molecules that are the constituents of the matrix will probably result in structural changes in the matrix itself and, concomitantly, in changes in cell signalling. 33
Collagen undergoes several post-translational modifications that contribute to its structural and mechanical properties. Interference with some of these modifications results in severe dysfunction of the tissue in question. One of the last steps in the formation of the collagen molecule is the cleavage of the N and C propeptides, spontaneous self-assembly of the resulting collagen molecules into fibrils, and formation of covalent crosslinks. 34 Collagen crosslink formation occurs in the extracellular space, and the conversion of specific lysine or hydroxylysine residues into the aldehydes allysine or hydroxy allysine, respectively, initiate it. The enzyme lysyl oxidase catalyzes this crosslink reaction.35,36
The post-translational modifications of collagen in bone and other mineralizing tissues differ from those in other types of collagen matrices. 37 Proper collagen crosslinking may be important for the binding of collagen to its receptors, but it may also be important to regulate the availability of growth factors present in the ECM and for the mechanical properties of the ECM.38–40
In this article, we describe the effect of collagen crosslinking on the differentiation of MC3T3-E1 cells and hMSCs into the osteogenic lineage.
Materials and Methods
Cell culturing
Bone marrow aspirates (5–15 mL) were obtained from patients who had given written informed consent. hMSCs were isolated and proliferated as described previously. 41 hMSC proliferation medium was composed of alpha minimal essential medium (α-MEM; Gibco, Carlsbad, CA), 10% fetal bovine serum (Lonza, Verviers, Belgium), 0.2 mM ascorbic acid (Sigma Aldrich, St. Louis, MO), 2 mM L-glutamine (Gibco), 100 U/mL of penicillin (Invitrogen, Carlsbad, CA), 100 μg/mL of streptomycin, and 1 ng/mL of basic fibroblast growth factor (Instruchemie, Delfzijl, The Netherlands). Basic medium consisted of hMSC proliferation medium without basic fibroblast growth factor, hMSC osteogenic medium consisted of hMSC basic medium supplemented with 10−8 M dexamethasone (Sigma), and hMSC mineralization medium consisted of basic medium supplemented with 10−8 M dexamethasone and 0.01 M β-glycerophosphate (Sigma).
The MC3T3-E1 cell line was purchased from Riken Cell bank (RCB 1126) and cultured in MC3T3-E1 basic medium consisting of α-MEM (Biowhittaker, Thermo Fisher, Waltham, MA) 10% fetal calf serum (Hyclone), 2 mM L-glutamine (Invitrogen), 100 U/mL of penicillin (Invitrogen), and 10 μg/mL of streptomycin (Invitrogen). MC3T3-E1 control medium consisted of basic medium supplemented with 50 μg/mL of ascorbic acid (Sigma) and 10 mM β-glycerophosphate. MC3T3-E1 mineralization medium consisted of control medium supplemented with 100 ng/mL of recombinant human BMP-2 (R&D Systems, Abingdon, United Kingdom).
Assessment of cell proliferation was performed using an Alamar blue assay according to the manufacturer's protocol. Briefly, culture medium was replaced with medium containing 10% (vol/vol) Alamar blue solution (Biosource, Camarillo, CA), and cells were incubated at 37°C for 4 h. Fluorescence was measured at 590 nm on a Perkin Elmer LS50B plate reader (Wellesley, MA).
Alkaline phosphatase flow cytometry and biochemical analysis
The effect of beta-aminopropionitrile (BAPN) on alkaline phosphatase (ALP) expression of hMSCs was studied using flow cytometry on cells seeded at 1000 cells/cm2 and grown under various conditions. Each experiment included a negative control (cells grown in hMSC basic medium), a positive control (cells grown in hMSC osteogenic medium), and one or more experimental conditions (0.04 to 0.5 mM BAPN). After 7 days of culture, cells were trypsinized, and ALP expression was analyzed as described previously. 42
For MC3T3-E1 cells, we analyzed ALP activity biochemically. To this end, cells were lysed in 0.2% Triton X-100 buffered with 0.1 M potassium phosphate at pH 7.8. Subsequently, 40 μL of CDP-star reagent (Roche, Almere, The Netherlands) was added to a 10-μL aliquot of cell lysate and incubated for 30 min in the dark. Chemoluminescence was measured in a Victor plate reader (Perkin Elmer). Acidic phosphatase (ACP) activity was used to correct for the cell number. To this end, 5 μL of cell lysate was combined with 100 μL of buffered substrate solution (2.7 mM 4-nitrophenyl phosphate/0.1 M sodium acetate, pH 5.5). After 90 min, the reaction was terminated by addition of 10 μL of 1 M sodium hydroxide, and the absorbance was determined at 405 nm.
Mineralization and calcium assay
For mineralization experiments, MC3T3-E1 cells were seeded at 5 × 104 cells/cm2 in control and mineralization medium in the presence or absence of 1.6 mM BAPN for 10 days. After this period, the cells were lysed, and matrix-associated calcium was solubilized in 150 μL of 0.5 M hydrochloric acid (HCl) at 37°C overnight. A 2-μL aliquot of the solubilized calcium was combined with a solution of 0.25 M 2-amino-2-methyl-1,3-propandiol, 8.5 mM 8-hydroxyquinoline, and 190 μM o-cresolphthalein complexone (Sigma-Aldrich, Zwijndrecht, The Netherlands). Absorption was measured at 600 nm and compared with that of a standard curve of hydroxyapatite dissolved in 0.5 M HCl.
hMSCs were seeded in triplicate at 5000 cells/cm2 in T25 culture flasks and incubated with 0.5 mM BAPN for 21 days. In each experiment, hMSC mineralization medium was used as a positive control and hMSC basic medium containing β-glycerophosphate as a negative control. The total calcium deposition was assayed using a calcium assay kit (Bioassays Systems, Hayward, CA) according to the manufacturer's protocol. Briefly, the culture medium was aspirated, and cells were washed twice with calcium- and magnesium-free phosphate buffered saline (PBS; Life Technologies) and incubated overnight with 0.5 M HCl on an orbital shaker at room temperature. The supernatant was collected, and the calcium content was measured at 575 nm (Perkin Elmer). To visualize the calcium deposits, hMSCs were grown as mentioned above and fixed in formalin after 21 days of culture. One gram of alizarin red (Sigma) was added to 50 mL of demiwater and the pH was adjusted to 4.1 to 4.3. The solution was added for 3 min to the hMSCs previously fixed with formalin.
Collagen biochemical analysis
MC3T3-E1 cells were seeded onto 10-cm dishes in the presence of mineralization medium and cultured in the presence or absence of 1.6 mM BAPN. Collagen analysis was performed as previously described. 35 Briefly, MC3T3-E1 cells were grown for 16 days. Cultures were rinsed three times with PBS and extracted with 500 μL/10-cm dish of 1 M sodium chloride, 0.05 M Tris, 1 mM ethylenediaminetetraacetic acid, 10mM N-ethylmaleimide, and protease inhibitor (Roche) pH 7.5. After 5 min of incubation, cells were detached using a cell scraper, homogenized and centrifuged at 5 × 104 g for 2 h at 4°C. The resulting supernatant was designated the neutral salt extract. Fifty μg of neutral salt extract was loaded on a 4% to 12% BIS/TRIS sodium dodecyl sulfate polyacrylamide gel electrophoresis gel and stained with Coomassie blue.
To determine the level of collagen crosslinking and the collagen secreted by hMSCs, we cultured the cells in basic or mineralization medium in the presence or absence of 0.5 mM BAPN. After 21 days, samples were washed three times with PBS and hydrolyzed (110°C, 20–24 h) with 750 μL of 6 M HCl, dried and redissolved in 800 μL of water containing 10 μM pyridoxine (internal standard for hydroxylysylpyridinoline and lysylpyridinoline), and 2.4 mM homoarginine (internal standard for amino acids; Sigma). Samples were diluted 5 times with 1% (vol/vol) heptafluorobutyric acid (Fluka, Sigma) in 10% (vol/vol) acetonitrile for crosslinking analysis; aliquots of the 5-fold diluted samples were diluted 20 times with 0.1 M sodium borate buffer (pH 8.0) for amino acid analysis. Derivation and subsequent chromatography of the amino acids and chromatography of the crosslinks was performed as previously described.43,44 Collagen content was calculated from the total amount of hydroxyproline in each sample, assuming 300 Hyp residues per collagen molecule and a molecular weight of collagen of 300,000 D.
RNA isolation and quantitative polymerase chain reaction
The effect of inhibition of lysyl oxidase on expression of osteogenic marker genes was analyzed by seeding hMSCs at 5000 cells/cm2 in T25 flasks in osteogenic medium with or without BAPN for 10 days. Total RNA was isolated using an Rneasy mini kit (Qiagen, Venlo, The Netherlands) and on column DNase treated with 10 U of RNase free DNase I (Gibco) at 37°C for 30 min. DNase was inactivated at 72°C for 15 min. The quality and quantity of RNA was analyzed using gel electrophoresis and spectrophotometrically. One μg of RNA was used for first-strand complementary DNA (cDNA) synthesis using superscript II (Invitrogen) according to the manufacturer's protocol. One μL of 100 × diluted cDNA was used for collagen type I and 18S ribosomal RNA (rRNA) amplification, and 1 μL of undiluted cDNA was used for other genes. Polymerase chain reaction (PCR) was performed on a Light Cycler real-time PCR machine (Roche) using a SYBR green I master mix (Invitrogen). Data was analyzed using Light Cycler software version 3.5.3 using the fit point method by setting the noise band to the exponential phase of the reaction to exclude background fluorescence. Expression of osteogenic marker genes were calculated relative to 18S rRNA levels using the comparative ΔCT method. 45
Microarray analysis and statistical analysis
To analyze the effects of inhibition of lysyl oxidase on the gene expression profile of MC3T3-E1 cells and hMSCs undergoing osteogenic differentiation, we performed a whole genome analysis.
A time course analysis was performed on MC3T3-E1 cells cultured in mineralization medium in the presence or absence of BAPN. At least two microarrays were hybridized per time point (0, 24, 36, 48, 60, and 72 h). RNA was isolated using the Trizol reagent according to the protocol supplied by the manufacturer (Invitrogen). cRNA labeling, hybridization of Affymetrix Mouse Genome 430A Genechips (Affymetrix, Santa Clara, CA), and data processing was performed as described previously. 46
Normalization and statistical analysis of the data was performed using the error model developed for Affymetrix GeneChips performed using Rosetta Resolver Version 4.0. 47 In the case of hMSCs, 5000 cells/cm2 were seeded in T25 flasks in mineralization medium in the presence or absence of 0.5 mM BAPN. Total RNA was isolated using an Rneasy mini kit (Qiagen) and on-column DNase treated with 10 U RNase free DNase I (Gibco) at 37°C for 30 min. DNase was inactivated at 72°C for 15 min. The quality and quantity of RNA was analyzed using gel electrophoresis and spectrophotometrically. Affymetrix 900649 GeneChip Human Exon 1.0 ST Arrays were hybridized, and the fluorescence intensity was normalized by dividing the fluorescence intensity of each sample by the median intensity of the array. Statistical analysis was performed by application of a two-way analysis of variance model on each gene's profile using treatment and time as factors, resulting in p-values for the effect of treatment and time and for the interaction between treatment and time. These p-values were corrected for multiple testing using Benjamin-Hochberg, resulting in estimated false discovery rates (FDRs). Genes were ordered based on minimal FDRs for the treatment effect or the interaction effect. Genes were defined as significantly differentially expressed if this minimal estimated FDR was 1% or less.
Focal adhesion assembly
To analyze the assembly of focal adhesions by hMSCs, we cultured cells on glass coverslips in basic or osteogenic medium in the presence or absence of 0.5 mM BAPN. After 7 days, cells were fixed with 70% ethanol, permeabilized with 0.2% Triton X-100 in PBS for 20 min at room temperature, and washed with PBS. Samples were then blocked with 10% fetal bovine serum in PBS for 1 h at room temperature. A mouse anti-human vinculin monoclonal antibody (clone V284, Santa Cruz Biotechnology, Santa Cruz, CA) was used as a primary antibody. Samples were incubated with 1:100 diluted primary antibody for 2 h at 37°C, washed with 0.05% Tween in PBS, and incubated with 1:200 diluted secondary antibody (goat anti-mouse conjugated with AlexaFluor 488) and 1:40 phalloidin conjugated with rhodamine (Invitrogen) for 1 h at 37°C. Afterwards, samples were washed with 0.05% Tween in PBS, dried, and fixed with mounting medium containing 4',6-diamidino-2-phenylindole (Vectashield, Vector Laboratories, Amsterdam, The Netherlands). Cells were imaged using a Vector Laboratories BD Pathway Bioimager (BD Biosciences, Breda, The Netherlands).
Results
Inhibition of lysyl oxidase inhibits osteogenic differentiation of MC3T3-E1 cells
To analyze the role of collagen crosslinking in osteogenic differentiation of MC3T3-E1 cells we used BAPN, an irreversible inhibitor of lysyl oxidase, and analyzed its effects on collagen crosslinking, proliferation, ALP expression (an early marker of osteogenic differentiation), and calcium accumulation (a late marker of osteogenic differentiation). The time points selected to analyze each of these parameters was based on previous studies for BMP-induced osteogenic differentiation of MC3T3-E1 cells. 48
First, to confirm that BAPN was able to inhibit collagen crosslinking, we analyzed the form of collagen extracted from MC3T3-E1 cells cultured for 16 days in the presence or absence of 1.6 mM BAPN. In the presence of BAPN, we observed an accumulation of non-crosslinked α1 and α2 tropocollagen chains (Fig. 1A), indicating that BAPN leads to a decrease in the level of collagen crosslinking, causing the accumulation of tropocollagen molecules in the neutral salt extract.

Inhibition of lysyl oxidase impairs in vitro osteogenesis of MC3T3-E1 cells. (
Second, to examine whether collagen crosslinking influences proliferation of MC3T3-E1 cells, we exposed the cells to increasing concentrations of BAPN in control or mineralization medium. We observed that, independent of the medium, BAPN did not affect proliferation of MC3T3-E1 cells (Fig. 1B).
Third, we analyzed ALP expression of MC3T3-E1 cells upon exposure to different concentrations of BAPN in control and mineralization medium. As expected, a more-robust induction of ALP levels was evident for the MC3T3-E1 cells in mineralization medium than in control medium because of the exposure to BMP-2 (Fig. 1C). No significant change in ALP expression was observed in MC3T3-E1 cells cultured in control medium in the presence of increasing concentrations of BAPN. In contrast, when cells were cultured in mineralization medium in the presence of increasing concentrations of BAPN, we observed a dose-dependent decrease in ALP expression (Fig. 1C).
Finally, to analyze the ability to deposit calcium, we cultured MC3T3-E1 cells in control or in mineralization medium in the presence or absence of 0.4mM BAPN for 10 days. Exposure of MC3T3-E1 cells to mineralization medium for 10 days resulted in calcium accumulation three times as great as in control medium. When MC3T3-E1 cells were cultured in mineralization medium in the presence of BAPN, we observed a 2-fold decrease in calcium accumulation relative to MC3T3-E1 cells cultured in mineralization medium alone (Fig. 1D).
In conclusion, BMP-2-induced ALP expression and calcium accumulation in MC3T3-E1 cells were repressed when the level of collagen crosslinking was reduced.
Lysyl oxidase activity is required for ECM gene expression of MC3T3-E1 cells
To unravel the molecular mechanisms by which BAPN inhibits osteogenic differentiation of MC3T3-E1 cells, we analyzed the gene expression profile of MC3T3-E1 cells undergoing osteogenic differentiation in the presence or absence of 1.6 mM BAPN. Figure 2A shows the expression intensities of several relevant transcription factors and ECM-related genes in BAPN-treated and control cells as a function of time. In accordance with the effect of BAPN on ALP enzymatic activity, expression of the cognate ALP gene (AKP2) was repressed in BAPN-treated MC3T3-E1 cells (Fig. 2A).
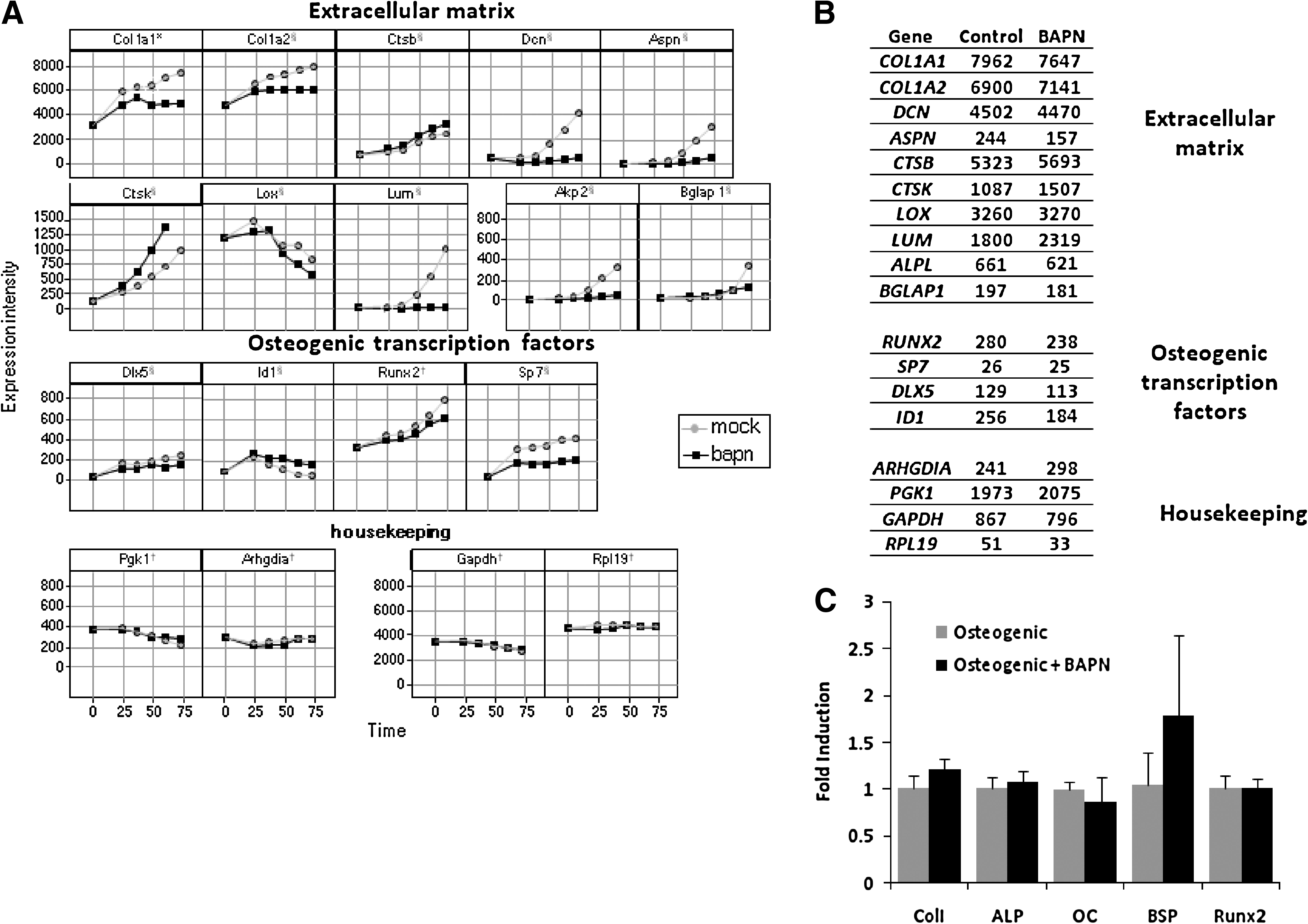
Gene expression profile of MC3T3-E1 and human mesenchymal stem cells (hMSCs) upon treatment with a lysyl oxidase inhibitor. (
Moreover, we observed that many ECM-related genes and osteogenic transcription factors were expressed at a lower level in MC3T3-E1 cells exposed to BAPN than in the control. Expression of the collagen type I α1 chain and the collagen type I α2 chain occurred as early as 24 h in mineralization medium and was lower when MC3T3-E1 cells were exposed to BAPN. The steep rise in collagen expression in control medium coincided with an increase in the expression of the enzyme responsible for the crosslinking lysyl oxidase. Treatment with BAPN led to a lower level of lysyl oxidase expression. Additionally, genes such as decorin, asporin, and lumican, which are known to be involved in matrix assembly and are highly expressed in later stages of differentiation, were not induced when MC3T3-E1 cells were exposed to BAPN. The osteogenic marker bone gamma-carboxyglutamate protein, related sequence 1 (BGLAP1) was also expressed at a lower level upon exposure to BAPN. The lack of induction of the BGLAP1 and ALPL genes coincides with the lack of induction of other ECM-related genes, pointing at a relationship between the expression of bone markers and ECM genes in osteogenic differentiation of MC3T3-E1 cells. Transcription factors that control the osteogenic differentiation program (Runt-related transcription factor 2 (Runx2), transacting transcription factor 7 (Sp7), and distal-less homeobox 5 (Dlx5)) showed lower expression upon exposure to BAPN than control cells. The lack of induction of the early transcription factors may explain the effects of BAPN on LOX, COL1A1, and other ECM-related genes, because these are genetically downstream of the Runx2-Sp7 pathway. 49 Although our data indicate that the majority of genes involved in osteogenic differentiation failed to be induced upon exposure of MC3T3-E1 to BAPN, some genes were upregulated. Among those, we observed an upregulation of cathepsins and some metalloproteinases (Fig. 2A and Supplementary data). A well-known BMP-2 target gene (ID1) was upregulated upon exposure of MC3T3-E1 cells to BAPN.
BAPN did not affect the expression of housekeeping genes, indicating that the effect observed in ECM genes and in genes encoding transcription factors was not the result of an aspecific effect of BAPN on the cellular processes.
Together, these results show that BAPN inhibits collagen crosslinking and thereby affects the transcriptional program underlying osteogenic differentiation of MC3T3-E1 cells.
Inhibition of lysyl oxidase influences in vitro osteogenic differentiation of hMSCs
Next, we examined whether inhibition of lysyl oxidase had similar effects on our cell model for tissue engineering applications—hMSCs. When hMSCs were exposed to 0.5 mM BAPN, crosslinks were absent, as indicated by the undetectable levels of the pyridinoline crosslinks hydroxylysylpyridinoline and lysylpyridinoline of the collagen present in the deposited matrix (in the absence of lysyl oxidase activity, the telopeptide hydroxylysine residues cannot be modified into aldehydes, and as so, crosslinking is absent), demonstrating that BAPN, as in the case of MC3T3-E1 cells, was able to inhibit the activity of lysyl oxidase in hMSCs (Fig. 3A). This concentration of BAPN can be considered equipotent to the one used in the case of MC3T3-E1 cells, because in both cases, there was inhibition of lysyl oxidase. Moreover, upon exposure of hMSCs to BAPN, the collagen content was lower than in the respective controls (54% lower in the case of basic versus basic + BAPN and 93% lower in the case of mineralization versus mineralization + BAPN) (Fig. 3B). Additionally, there was a decrease in the Hyp:Pro ratio, which represents the ratio of collagen to noncollagenous proteins. When hMSCs were exposed to BAPN, the Hyp:Pro ratio was 32% lower the case of basic versus basic +BAPN and 57% lower in the case of mineralization versus mineralization + BAPN (Fig. 3C). Taken together, these results showed that BAPN was able to inhibit lysyl oxidase activity and impair collagen crosslinking of hMSCs.

Collagen biochemical analysis in the presence of an inhibitor of collagen crosslinking. (
Given the fact that, upon exposure to BAPN, hMSCs showed changes in collagen structure and synthesis, we analyzed whether those changes would have an effect on integrin signalling by analyzing focal adhesion assembly in the presence or absence of BAPN. Our analysis showed that BAPN did not affect focal adhesion assembly (Fig. 4A).

Inhibition of lysyl oxidase influences in vitro osteogenesis of hMSCs. (
Subsequently, we analyzed the effect of lysyl oxidase inhibition on proliferation and ALP expression. We cultured hMSCs in basic (data not shown) and osteogenic medium in the presence of increasing concentrations of BAPN for 7 days. BAPN did not affect proliferation of hMSCs during the period of culture considered (Fig. 4B). Exposing hMSCs to increasing concentrations of BAPN did not affect dexamethasone-induced ALP expression (Fig. 4C).
Next, we performed a whole genome analysis on hMSCs exposed to BAPN. Exposure of hMSCs to BAPN for 10 days had little or no effect on gene expression. In contrast with MC3T3-E1 cells, we saw that, upon exposure of hMSCs to BAPN, no ECM genes were downregulated at the time point considered. Additionally, there were no drastic changes in the expression of osteogenic transcription factors, or in the housekeeping genes, when hMSCs were exposed to BAPN (Fig. 2B). To validate the results from the microarray, we performed quantitative PCR (qPCR) for a panel of osteogenic marker genes in hMSCs cultured for 10 days in osteogenic medium in the presence or absence of 0.5 mM BAPN. Similar to the results of the microarray, no statistically significant changes in the expression of these osteogenic genes was observed (Fig. 2C). These results demonstrate a good correlation between the microarray measurements and qPCR results, as previously shown. 46 Because lysyl oxidase is involved in the crosslinking of collagen, which is the main component of the matrix deposited by terminally differentiated osteoblasts, we analyzed the effect of BAPN on matrix mineralization. We cultured hMSCs in basic or mineralization medium in the presence or absence of 0.5 mM BAPN for 21 days and measured calcium accumulation. hMSCs treated with BAPN deposited up to six times as much calcium as hMSCs treated with mineralization medium alone for all six donors tested. The greater calcium deposition was observed only for hMSCs cultured in mineralization medium (Fig. 4D).
Discussion
In vitro osteogenic differentiation of MC3T3-E1 cells and effects of lysyl oxidase inhibition
Differentiation of MC3T3-E1 cells into the osteogenic lineage is regarded as a matrix-driven process. In the presence of ascorbic acid, the cells deposit a collagen matrix that later will be mineralized in the presence of inorganic phosphate. 50 The synthesis and deposition of this collagen matrix provides a positive feedback signal to activate and maintain the osteogenic program. In the absence of that signal, the cells do not undergo osteogenic differentiation. 13 The structure and abundance of collagen can affect the positive feedback signal provided by the ECM. It has been shown that proliferation, as well as calcium deposition by MC3T3-E1 cells grown on denatured collagen, can be enhanced, dose-dependently, by adding increasing concentrations of microfibrillar collagen to the culture system. 51 This shows that it is possible to interfere with the signal provided by the collagen matrix to enhance the osteogenic potential of MC3T3-E1 cells.
As previously shown, 35 inhibition of lysyl oxidase inhibits collagen crosslinking and affects collagen structure, resulting in the accumulation of soluble tropocollagen. Similar to the results showing a decrease on BMP-induced ALP expression upon blocking of integrins, we observed a dose-dependent decrease in ALP expression upon exposure of MC3T3-E1 cells to BAPN. 52 Moreover, our results showed that calcium deposition by MC3T3-E1 was also lower when collagen crosslinking was inhibited. In the absence of collagen crosslinking, the collagen is weakly associated with the cell membrane, and it is more prone to proteolytic degradation. 35 Therefore, we cannot exclude that the effects we observed are the result of lower levels of collagen.
It has been shown that proliferation of MC3T3-E1 cells was impaired in the absence of ascorbic acid. 53 In our case, we did not observe effects on proliferation upon exposure to BAPN, which indicates that sufficient levels of collagen were still present in our culture.
Treatment of MC3T3-E1 cells with BAPN led to a lack of induction of expression of the transcription factors Dlx5, Runx2, and Sp7. Because these factors are known to initiate osteogenic differentiation, their downregulation suggests a feedback mechanism in which ECM supports maintenance of expression of these early genes (Fig. 5). BAPN inhibited the positive feedback signal provided by collagen upon stimulation with BMP-2, leading to a lack of induction of ECM-related genes. The decrease in the expression of ECM genes supports the lower mineralization because these proteins may act as mineralization nucleators. 54 Impairment of differentiation was not a result of inhibition of BMP signalling because Id1, a direct target of BMP-2-induced Smad signalling, was not negatively affected but was actually slightly upregulated upon exposure of MC3T3-E1 cells to BAPN. This suggests that the collagen matrix acts downstream of Smad signalling to enhance Dlx5, Runx2, and Sp7. The exact nature of the signal is currently unknown.

Proposed mechanism of action of BAPN on MC3T3-E1 cells. The figure depicts the osteogenic differentiation of MC3T3-E1 cells upon exposure to bone morphogenetic protein 2 and extracellular matrix. In black are the genes and proteins negatively affected by BAPN, and in white are the genes and proteins that were upregulated. Proteins are represented by square boxes, and RNAs are in circles. Rounded dot squares indicate the phenotype on hMSCs and MC3T3-E1 cells upon exposure to BAPN, whereas rounded dashed squares represent a mechanism conserved in both cell types.
Effects of lysyl oxidase inhibition on in vitro differentiation of hMSCs
Because collagen plays an essential role in osteogenic differentiation of MC3T3-E1 cells, we were interested in analyzing its role in hMSCs, which are widely used in bone tissue engineering. So far, mainly soluble factors have been used to enhance osteogenic differentiation of hMSCs, but it is technically not trivial to present these factors to the cells after implantation. 22 Exposure of implanted cells to ECM-coated scaffolds could be an interesting alternative. Discovering the role of the collagen matrix on osteogenic differentiation of hMSCs is therefore required. 9 In that respect, work done by others has showed that ECM deposited by MC3T3-E1 cells can affect differentiation into the osteogenic lineage. 55 The immense potential of ECM has been demonstrated using decellularization methods to remove the cellular components and seeding new cells onto the matrix. The ECM produced in this way was able to influence the differentiation of the newly seeded cells and at the same time provided an appropriate structure to support cell growth.2,56–58
We made two interesting observations. First, osteogenic differentiation of hMSCs is not sensitive to the level of crosslinking of the matrix, in contrast to MC3T3-E1 cells. Not only did inhibition of collagen crosslinking not influence ALP activity in hMSCs, but also the global gene expression profile of hMSCs was virtually unchanged after exposure to BAPN for 10 days. Compared with untreated cells, only 20 genes were positively or negatively regulated 2 or more times in the presence of BAPN. Among those, no ECM genes were present. This indicates that the positive feedback loop provided by the crosslinked collagen matrix observed in MC3T3-E1 cells is absent in hMSCs.
The decrease in collagen abundance observed in hMSCs exposed to BAPN could be attributed to an increase in proteolytic degradation of collagen due to a reduction in the number of crosslinks or the fact that most of the collagen was soluble and, as such, was not accounted for in the analysis.
The second interesting observation is the increase in calcium deposition upon exposure of hMSCs to BAPN. Normally, calcium deposition starts at specific residues in the collagen molecule, in particular in the gap junctions between adjacent collagen molecules. Inhibition of crosslinking may expose cryptic nucleation sites, leading to enhanced mineralization.59–61 Gene expression data exclude an increase in matrix components as an explanation for enhanced mineralization. As such, we hypothesize that the absence of crosslinks in the collagen matrix provides more space for crystal growth in the collagen fiber. The observed increase in mineralization might be similar to the enhanced mineralization in patients with OI, which can be due to an increase in the density of nucleation centers in the bone matrix. 29 Although collagen provides the structural framework for mineral deposition, noncollagenous proteins are responsible for the initiation and regulation of biomineralization. 54 Another hypothesis for the increase in calcium accumulation is that improper deposition of collagen results in the release of some noncollagenous proteins, which will act as mineralization nucleators.
The results presented herein highlight the fact that the results gathered in one cell type cannot be directly extrapolated to another cell type without taking into consideration the species difference or even the origin and differentiation state of the cells. 62 In this particular case, we analyze the role of collagen crosslinking in a commonly used cell line in bone cell biology—MC3T3-E1 cells—and compare it with the more clinically relevant hMSCs. These cells are from different origins and show a different differentiation potential. Whereas hMSCs can differentiate into several lineages depending on the initial trigger, MC3T3-E1 are precommitted to the osteogenic lineage. Furthermore, MC3T3-E1 cells originate from murine calvaria, whereas hMSCs originate from human bone marrow. As such, we cannot exclude that the differences observed herein are due to the different origins of these two cells types.
Conclusion
In conclusion, we show that the differentiation of MC3T3-E1 cells into the osteogenic lineage is impaired as a result of inhibition of collagen crosslinking. Exposure of MC3T3-E1 cells to BMP-2 was sufficient to drive the osteogenic differentiation program, leading to collagen production, which in turn fed back to osteogenic transcription factors. The inhibition of collagen crosslinking abolishes this positive feedback loop, resulting in impaired osteogenic differentiation (Fig. 5). In contrast, the levels of collagen crosslinking did not affect the osteogenic differentiation process of hMSCs, although we observed an interesting effect of crosslinking on mineralization.
Footnotes
Acknowledgments
We gratefully acknowledge Roland Heerkens and Rianne Assink for technical assistance. The research of HF, KD, EvS, AL and JdB was sponsored by a grant from Senter/Novem.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.