
Abstract
Background:
Vascular smooth muscle cells (VSMCs) are a potential autologous cell source for aortic valve tissue engineering, but have a phenotype that differs from that of valvular interstitial cells in vivo. We hypothesized that combining basic fibroblast growth factor (bFGF), epidermal growth factor (EGF), or platelet-derived growth factor (PDGF) with transforming growth factor beta-1 (TGF-β1) would achieve a valvular interstitial cell–like phenotype of VSMCs.
Methods:
VSMC phenotype was assessed by immunofluorescence, proliferation was measured by the tetrazolium reduction (MTT) assay, and extracellular matrix gene expression was determined by real-time polymerase chain reaction.
Results:
Combinations of growth factors that included PDGF showed the greatest increases in proliferation. Immunofluorescence for α-smooth muscle actin demonstrated an inverse correlation between proliferation and a myofibroblast-like phenotype, while combinations of TGF-β1+ EGF+bFGF (TEF) and TGF-β1+EGF+PDGF (TEP) induced the greatest change of α-smooth muscle actin expression compared to untreated controls. Finally, TEP treatment showed an increase in versican, fibronectin, and type I collagen mRNA expression, while decreasing matrix metalloproteinase 1 expression.
Conclusions:
Combination of TGF-β1 with EGF and PDGF induces VSMC proliferation and expression of extracellular matrix constituents found in the aortic valve. In vitro preconditioning of VSMCs provides a potentially viable surrogate cell source for developing a valve graft.
Introduction
VICs populate and maintain the three major layers of the aortic valve.2,5,6 The most superior layer, the fibrosa, is composed primarily of type I and, to a lesser extent, type III collagen. 5 The spongiosa contains abundant proteoglycans to retain water within the tissue and provide viscoelasticity. 7 Finally, the ventricularis contains large amounts of elastin close to the outflow surface of the valve and is therefore responsible for returning the valve to its relaxed state during diastole.8,9 A potential surrogate cell source must maintain each of these valvular attributes to recreate a valve-like tissue suitable for implantation.
The VICs that maintain the trilaminar valve structure in vivo are mostly in G0 arrest and hence undergo a slow rate of turnover.10,11 Further, after tissue injury that may occur under conditions of disturbed hemodynamic flow or in the diseased valve, VICs take on a myofibroblast-like phenotype. 11 Myofibroblast-like VICs express high levels of α-smooth muscle actin (α-SMA); become contractile, proliferative, and highly synthetic; and are commonly observed in calcific valvular sclerosis.6,12–15 While a synthetic and proliferative cell population is beneficial for tissue engineering, control of this myofibroblast-like phenotype is required to prevent valve calcification and degeneration.
VSMCs reside in a complex extracellular matrix (ECM) environment containing types I and III collagen, proteoglycans, and elastin.4,16,17 They transit between a contractile and a synthetic/proliferative phenotype in vivo, the latter being considered a dedifferentiated state.4,18 In vitro, the transition between these two phenotypes is dependent upon culture conditions. For example, transforming growth factor beta-1 (TGF-β1) is known to promote a myofibroblast-like phenotype, similar to the synthetic/proliferative phenotype seen in vivo. 4 However, these myofibroblast-like cells tend to contract and shrink their scaffold in three-dimensional (3D) culture.19,20 This places uncontrolled mechanical stress on the cells and further modifies phenotype. Hence, further modulation to culture conditions must be sought to tailor these cells appropriately for valve engineering.
Basic fibroblast growth factor (bFGF), epidermal growth factor (EGF), and platelet-derived growth factor (PDGF) have pro-synthetic and pro-proliferative effects on VSMCs and antagonize some of the effects of TGF-β1.21–24 In the present study, we show that specific treatment of VSMCs with cocktails of exogenous growth factors can result in the development of a VIC phenotype suitable as a surrogate cell source for valvular tissue engineering applications.
Materials and Methods
Cell isolation and culture
Rat aortic smooth muscle cells (RASMCs) were harvested from adult male Sprague-Dawley rats (Charles River, St. Constant, Canada) as a source of VSMCs. This investigation conforms with the Guide for the Care and Use of Laboratory Animals published by the U.S. National Institutes of Health (NIH Publication No. 85-23, revised 1996). The use of rat tissues for these experiments was approved by the Animal Care and Use Committee at the University of Western Ontario. Aortas were harvested using sterile technique and washed in sterile phosphate-buffered saline (PBS) containing 1% penicillin–streptomycin. Fat and adventitia were removed and the intima was scraped to remove endothelium. The aortas were minced into 1 mm2 pieces and digested for 1.5 h at 37°C in Dulbecco's modified Eagle's medium (DMEM) containing 10 units/mL elastase (Sigma, Oakville, Canada) and 75 units/mL collagenase (Sigma, Oakville, Canada). After digestion, cells were isolated by passing through 40 μm cell strainers (BD Falcon, Bedford, MA) and washed with fetal bovine serum (FBS). Cells in suspension were pelleted in a centrifuge at 1200 rpm for 5 min, resuspended in DMEM, and plated for culture expansion. Cells were then incubated at 37°C in a humidified environment containing 5% CO2 in DMEM supplemented with 10% FBS and 1% penicillin–streptomycin. Medium was changed every 3 days. Before treatment, cultures were switched to serum-free DMEM with 1% penicillin–streptomycin for 24 h.
Growth factor treatment
All growth factors, including TGF-β1, EGF, bFGF, and PDGF, were obtained from R&D Systems (Minneapolis, MN). For treatment, all growth factors were used at a concentration of 10 ng/mL (based on dose–response data not shown) in DMEM (Gibco, Burlington, Canada) growth medium containing 2.5% FBS, 1% penicillin–streptomycin, 2 μg/mL insulin, 50 μg/mL ascorbic acid, and 0.25% L-glutamine. All possible combinations of TGF-β1 plus one to all of the other three growth factors were used in initial experiments.
Cell proliferation
RASMC proliferation was measured using an tetrazolium reduction (MTT) assay kit (Molecular Probes, Eugene, OR). Cells were seeded at a density of 1.0 × 104 cells/well into 96-well plates and allowed to grow for 3 days in DMEM with 10% FBS and 1% penicillin–streptomycin. RASMCs were then serum-starved for 24 h before treatment. After treatment of 1, 3, or 5 days, 10 μL of the MTT reagent, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, was added to each well and incubated at 37°C for 4 h. The cells were then lysed in solubilization buffer to release the MTT reagent and incubated overnight. MTT reagent absorbance was read at 570 nm according to the manufacturer's protocol.
In-cell western
Cells were plated at a density of 1.0 × 104 cells/well into 96-well plates and incubated for 3 days at 37°C. Culture medium was replaced with serum-free medium 24 h before treatment. RASMCs were fixed with 10% formalin at room temperature for 20 min after 72 h of treatment and permeabilized with 0.1% Triton X-100 in PBS. Plates then underwent serial washes with 0.1% Tween-20/PBS between blocking, primary and secondary antibody incubations. Primary antibodies used included mouse anti-rat α-SMA (Sigma, Burlington, Canada), rabbit anti-rat CD90 (Abcam, Cambridge, MA), and rabbit anti-rat h-caldesmon (h-cal) (Abcam). Primary antibody incubation was conducted overnight on a plate shaker at 4°C. Fluorophore-conjugated secondary antibodies for the Odyssey® Infrared Imaging System (LI-COR Biosciences, Lincoln, NE) were incubated for 1 h at room temperature. Normalization to DNA content was achieved by incubating DRAQ5 (Biostatus, Leicestershire, United Kingdom) and Sapphire 700 (LI-COR Biosciences) fluorescent reagents simultaneously with secondary antibody. Signal detection was performed using the LI-COR Odyssey Infrared Imaging System. Target signal was normalized to DNA content signal within each well after subtracting background signal from wells without primary antibody.
Collagen gel constructs
Type I rat tail collagen (BD Biosciences, Bedford, MA) was used at a final concentration of 1.5 mg/mL after dissolving in the treatment medium. Chondroitin-6-sulfate from shark cartilage (Sigma, Oakville, Canada) and fibronectin (Sigma, Oakville, Canada) were added to the collagen gel at concentrations of 1 mg/mL and 50 μg/mL, respectively. For 3D cultures, 500 μL/well of collagen gel was added to 24-well culture plates and allowed to set for 1 h at 37°C, before seeding with 5.0 × 105 cells on the surface of the gel.
RNA isolation and real-time polymerase chain reaction
Total RNA was isolated from primary RASMC cultures using QIAshredder columns and a QIAgen RNeasy Mini kit according to the manufacturer's protocol (Qiagen, Mississauga, Ontario, Canada). Total RNA was isolated from collagen gel constructs (CGCs) using a Trizol (Invitrogen, Burlington, Ontario, Canada) extraction and glass tissue homogenization, followed by QIAgen RNeasy Mini kit. RNA quality and concentration was measured using the ND 1000 Nanodrop spectrophotometer (Nanodrop Products, Wilmington, DE). Real-time polymerase chain reaction (PCR) was performed using the Applied Biosystems 7900HT Real-Time PCR system. Probes were acquired as TaqMan Gene Expression Assays and prepared using TaqMan One Step Mastermix (Applied Biosystems, Foster City, CA). Changes were assessed relative to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) expression. Probes used include procollagen type 1 α-1, procollagen type III α-1, versican, elastin, matrix metalloproteinase 1 (MMP1), α-SMA, fibronectin, and GAPDH. Sequences for these probes are not listed due to their proprietary nature.
Immunofluorescence
RASMCs were seeded at a density of 3.0 × 104 cells/well onto glass coverslips in 24-well plates. Cells were fixed with 10% formalin for 1 h at room temperature after 48 h of treatment and permeabilized with 0.05% Triton-X-100 in PBS and probed with Cy3-conjugated mouse anti-rat α-SMA primary antibody (Sigma, Oakville, Ontario, Canada) and phalloidin conjugated to Alexa Fluor 488 (Invitrogen) to observe F-actin. Coverslips were then mounted with fluorescent mounting medium (Dako, Carpinteria, CA) and imaged using a Zeiss LSM410 confocal microscope (Carl Zeiss, Toronto, Ontario, Canada).
Immunohistochemistry
CGCs were fixed in 10% formalin for 4 h at room temperature, processed, embedded in paraffin, and sectioned at 6 μm thickness. Sections were probed with rabbit anti-rat MMP9 primary antibody (Abcam) and goat anti-rabbit IgG secondary antibody conjugated to HRP (Vector, Burlington, Ontario, Canada). Colorimetric detection was completed using diaminobenzidine substrate (Dako). Nuclei were counterstained with Harris's hematoxylin.
Statistical analysis
All experiments were completed with cells from four or five independent animals. Real-time PCR data were run in triplicate, quantified relative to GAPDH expression, and normalized to the untreated control for each animal. Real-time PCR data and MTT proliferation assay data were analyzed by two-way analysis of variance with a Tukey's post hoc test using GraphPad Prism 4.0 software (GraphPad Software, San Diego, CA). In-cell western data were analyzed by two-tailed Student's paired t-test. In all cases p-values less than 0.05 were considered significant.
Results
Expression of α-SMA in RASMCs
TGF-β1 is known to induce α-SMA expression and promote a myofibroblast-like phenotype in vascular smooth muscle. 4 To investigate TGF-β1–mediated effects in combination with other growth factors (e.g., EGF, bFGF, and PDGF), α-SMA was observed by immunofluorescence. Untreated (control) cultures demonstrated baseline α-SMA expression and TGF-β1 treatment induced α-SMA as expected (Fig. 1). Overall, TGF-β1 combined with EGF+bFGF (TEF) induced the greatest increase, while TGF-β1 combined with EGF+PDGF (TEP) induced the greatest decrease in α-SMA expression compared to untreated controls (Fig. 1). Other combinations of growth factors had little or no detectable effect (not shown). Thus, TEF and TEP treatments were used for subsequent experiments.
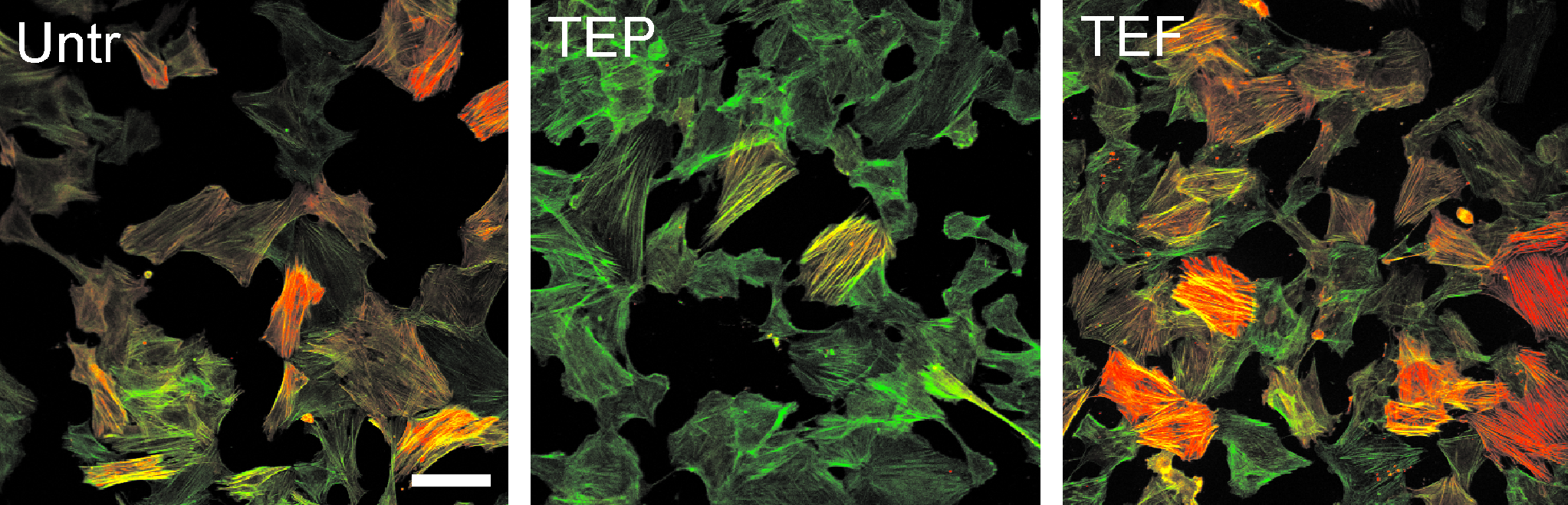
RASMC phenotype by immunofluorescence for α-SMA. RASMCs were grown to 75% confluence on coverslips and treated with various combinations of growth factors, each at a concentration of 10 ng/mL for 48 h. Cells were then fixed and stained with Alexfluor-488–labeled phalloidin to show total actin (green signal) and Cy3-conjugated monoclonal antibody for α-SMA (red signal) and imaged by confocal microscopy. The combination of TGF-β1+EGF+PDGF (TEP) appeared to decrease α-SMA expression, while TGF-β1+EGF+bFGF (TEF) showed an increase in the same. Untr, vehicle only; T, TGF-β1; E, EGF; F, bFGF; P, PDGF. Scale = 100 μm. RASMC, rat aortic smooth muscle cell; α-SMA, α-smooth muscle actin; TGF-β1, transforming growth factor beta-1; EGF, epidermal growth factor; PDGF, platelet-derived growth factor; bFGF, basic fibroblast growth factor.
Changes in RASMC number with growth factor treatments
TGF-β1 in combination with either bFGF or PDGF induced an increase in cell number compared to untreated controls after 24 h of treatment, but only TGF-β1+bFGF maintained this increase after 5 days (data not shown). The greatest increases in cell number were seen when PDGF was added to cultures with two or more other growth factors. TEP treatment caused the largest increase (>twofold) over the 5-day time course (Fig. 2), but no significant changes were observed when TGF-β1 was combined with EGF and bFGF (Fig. 2).

Effects of growth factor treatment on RASMC cell number. MTT assay was used to measure relative numbers of RASMCs after 1, 3, and 5 days of treatment. Cells were grown in monoculture with either no treatment or a combination of growth factors including TGF-β1+EGF+bFGF (TEF) or TGF-β1+EGF+PDGF (TEP) (n = 4 cell isolations from independent animals). TEP induced an increase in cell number, while TEF had no effect. Significantly different means (+standard error of mean) are indicated by different letters (p < 0.05).
ECM gene expression in monolayer culture
Real-time PCR was used to investigate ECM gene expression changes at the transcriptional level. Unexpectedly, both TEF and TEP treatment reduced elastin (Eln) mRNA levels. However, TEF and TEP treatments induced fibronectin 1 (Fn1) and type I collagen mRNA, while only treatment with TEP increased the proteoglycan versican (Vcn) expression (Fig. 3). Taken together, this suggests that TEP treatment decreases nonvascular and increases vascular-specific proteoglycan gene expression in RASMCs. While no effect of treatment was observed on type III collagen, expression of interstitial collagenase (Mmp1) was reduced by TEF and TEP treatments (Fig. 3). Overall, these results suggest that an anabolic profile of matrix gene expression is stimulated in RASMCs in response to TEF and TEP treatment.

Effects of treatment on RASMC expression of extracellular matrix mRNA in monolayer culture. RNA was isolated from RASMCs (n = 4) in monoculture after 48 h of conditioning with no treatment, TGF-β1+EGF+bFGF (TEF), or TGF-β1+EGF+PDGF (TEP) growth factor combinations. Changes in mRNA expression were analyzed by real-time polymerase chain reaction. Both treatments increased fibronectin and type I collagen expression, while decreasing elastin and interstitial collagenase. Only TEP induced a significant increase in versican expression. Significantly different means (p < 0.05) are indicated by letters.
Assessment of RASMC phenotype
Since TEP treatment resulted in increased proliferation and decreased α-SMA expression in addition to inducing the most favorable profile of ECM gene expression, it was selected as the treatment of choice for further investigations. To analyze TEP treatment effects on RASMC phenotype, in-cell westerns were used to detect changes in the expression of specific marker proteins. A decrease in myofibroblast (α-SMA and h-cal) and fibroblast (CD90) phenotypic marker expression was discovered in response to treatment with TEP (Fig. 4A, B). Further, untreated RASMCs seeded on CGCs showed greater contraction of the gel over time than constructs treated with TEP (Fig. 4C). Similarly, immunohistochemistry identified a decrease in expression of MMP9 (Fig. 4D), a known marker of stressed and contractile VSMCs. 25 Collectively, these results indicate that TEP treatment may induce a dedifferentiated, less stressed, and less contractile RASMC phenotype.

Analysis of RASMC phenotype after treatment with TGF-β1+EGF+PDGF (TEP) growth factor combination versus untreated control (Untr). (
ECM gene expression in CGCs
To better understand the effects of TEP treatment in valve-like, 3D conditions, RASMCs were seeded onto type I collagen/chondroitin-6-sulfate/fibronectin gels, treated, and analyzed by real-time PCR. Changes in ECM gene expression induced by TEP treatment in CGCs were similar to those observed in monolayer cultures. α-SMA, Eln, and Mmp1 mRNA expression decreased, while Col1a1, Fn1, and Vcn expression increased, although increases in Col1a1 and Vcn mRNA levels were not statistically significant compared to untreated controls (Fig. 5). Col3a1 mRNA levels did not change with TEP treatment (Fig. 5).

Effects of treatment on RASMC expression of extracellular matrix mRNA in three-dimensional culture. About 5 × 105 RASMCs (n = 4) were seeded onto preset, 500 μL type I collagen/chondroitin-6-sulfate/fibronectin gels in 24-well culture dishes. Treatment with TGF-β1 +EGF+PDGF (TEP) was administered over 72 h. RNA was isolated from RASMCs on CGCs after treatment. Changes in mRNA expression were analyzed by real-time polymerase chain reaction. Treated CGCs saw a significant increase in fibronectin expression with increasing trends in versican and collagen type I. Elastin, MMP1, and α-SMA expression was reduced. Significantly different means (p < 0.05) are indicated by asterisks.
Discussion
A surrogate cell source for VICs is needed for aortic valve tissue engineering. VSMCs may be a viable surrogate, provided that the phenotype of VSMCs may be manipulated to mirror that of endogenous VICs. In this study, we investigated the use of growth factor cocktails to induce VIC-like characteristics in VSMCs. We have shown that specific growth factor treatment in culture may yield increased proliferative capacity and ECM gene expression patterns appropriate for the growth and remodeling of a tissue-engineered valve. In particular, the combination of TGF-β1 with EGF and PDGF (TEP) resulted in increased cell numbers and upregulation of important proteoglycans, such as fibronectin and versican. While TGF-β1 may induce or inhibit proliferation in a context-dependent manner, 26 VSMC proliferation is typically induced by EGF, bFGF, and PDGF through the activation of pro-mitogenic mitogen-activated protein kinase pathways.21–23,27–29 However, in our studies, PDGF and EGF appeared to exhibit synergistic activity, whereas the combination bFGF and EGF antagonized the induction of mitogenesis. Thus, it is possible that EGF and PDGF, in combination, synergistically enhance the activation of select mitogen-activated protein kinase (MEK/ERK or JNK) signaling and may also inhibit p38 mitogen–activated protein kinase. Further, the growth factors used in this study are anti-apoptotic and pro-migratory.30–32 These attributes will aid in the growth, survival, and remodeling capabilities of the VIC surrogate population. In addition, our data also suggest that TEP treatment favors anabolic activity through increases in type I collagen expression, with concomitant decreases in proteolytic enzymes such as MMP1. Maintenance of this gene expression profile in surrogate VICs is necessary for the production of a viable engineered valve, since these components are integral to the structure and function of healthy aortic valves in vivo.2,10,33
VICs in healthy aortic valves do not exhibit characteristics of myofibroblastic cells.12,34 Conversely, the myofibroblast-like cell phenotype is predominantly found in calcified and sclerotic valve tissue and may contribute to these valvular pathologies.10–12,35,36 However, this phenotype has also been found to be predominant in studies of fetal heart valves. 6 For the purposes of tissue engineering, mimicking the embryologic phenotype is best suited to producing appropriate stroma. Hence, while a synthetic, proliferative, myofibroblastic phenotype (as is observed in VICs under stress/fetal development) might be beneficial for bioengineered tissue remodeling a priori, it is an unequivocal sign of valve disease, in vivo, as the tissue matures.3,10,11,35,37 Thus, strategic optimization of both synthetic and nonmyofibroblastic characteristics will be most conducive to engineering healthy valvular tissue. VSMCs treated with TEP appear to fulfill these criteria. Our experiments showed that TEP treatment reduces the expression of markers of stressed, myofibroblast-like cells such as α-SMA and MMP9. 25 Further, TEP-treated VSMCs were highly synthetic and proliferative, but did not exhibit the same contractility in 3D cultures as untreated cells. Thus, an autologous VSMC source treated with TEP may be a viable surrogate for endogenous VICs.
Further evidence that TEP treatment pushes VSMCs away from myofibroblastic and smooth muscle cell phenotypes was given by the observed reduction in CD90 and h-cal. CD90 is commonly found on immune cells and fibroblasts. 31 Conversely, h-cal is known to be an indicator for terminally differentiated, contractile smooth muscle. 32 The reduction of both of these markers in addition to decreased α-SMA expression further demonstrates that TEP treatment stimulates a VIC-like phenotype. Moreover, TEP-treated VSMCs appear to undergo a process of dedifferentiation, likely due to the contribution of PDGF included in TEP treatment. 33 A dedifferentiated state may allow for strategic control of autologous cell phenotype and function, perhaps for individual augmentation of each valve layer.
Current approaches in valve engineering involve seeding autologous cells onto/into a biosynthetic scaffold with the hope that the seeded cells will remodel the provided matrix and eventually replace it with their own.3,6,17,38 Seeded cells must adhere, colonize, and remain viable within the chosen matrix. There is evidence that both VSMCs and VICs are readily viable within type I collagen matrices at the studied concentration.38,39 The surrogate VIC cell source must also exhibit a gene expression profile conducive to the maintenance of each of the three layers of the aortic valve. The fibrosa contains mainly collagens type I and III, the spongiosa contains abundant proteoglycans, and the ventricularis contains a strong elastin component.2,5,7 We have shown that RASMCs increase type I collagen transcription after treatment with TEP, likely due to the contribution of TGF-β1 stimulation.4,40,41 Other ECM components, such as fibronectin, are needed for tissue remodeling and cellular migration. Fibronectin facilitates cell adhesion to the ECM via integrin binding to collagen fibrils and heparan sulfate.36,42 This allows better attachment to the ECM as well as improved distribution of mechanical load throughout the matrix, an important factor to consider in the strained, dynamic environment of the valve. VICs also secrete fibronectin after tissue injury.36,43 The induction of fibronectin gene expression with TEP treatment further demonstrates that TEP-treated VSMCs may take on VIC-like properties.
Due to its abundant proteoglycan content, the spongiosa is highly hydrated and acts as a cushion for the valve tissue as it is cyclically stressed. Three different proteoglycans are most commonly found in the aortic valve, including biglycan, decorin, and versican.2,7 Recent studies by Stephens et al. found that versican is mostly located in the spongiosa layer, suggesting that versican may account predominantly for the shock-absorbing properties of the spongiosa. 7 While our investigations demonstrated an increase in versican expression with TEP treatment, Stephens et al. also found that versican, in particular, colocalized with elastin within the valve. 7 Perhaps the production of versican may serve to make a better environment for the synthesis and secretion of elastic fibers. Unfortunately, we have been unable to show an increase of elastin transcription with TEP treatment. This difficulty in inducing elastin expression has been commonly reported.44,45 Upregulation of elastin expression for the purposes of valve engineering has only been shown in embryonic VSMCs in vitro. 41 It has also been proposed that mechanical stimulation may increase elastin production. 42
In vitro mechanical strain conditioning is an important tool in heart valve tissue engineering.2,4,7,10,46 Other studies have reported that the use of in vitro cyclic strain conditions have increased contractility and α-SMA expression in VSMCs.2,4,47 While not undertaken in the present study, treatment of VSMCs with TEP under cyclic strain conditions may abrogate the induction of the myofibroblast-like contractile characteristics of VSMCs. If so, the proposed culture conditions may constitute an important advance in the field of valve tissue engineering and will be investigated further.
A limitation to this study is the use of animal cells. Ideally, human VSMCs would have been used, as they are the proposed VIC surrogate population. It has been previously shown that human VSMCs may be used in cardiovascular tissue engineering. 48 Additionally, the effects of growth factors on smooth muscle, compared across species, is fairly consistent. However, there is decreased response to certain growth factors, such as bFGF and PDGF, with aging. 49 Given that most heart valve transplants are completed in the elderly, this may be of concern with regard to cell yield and proliferative capacity. Conversely, there is also evidence to indicate that VSMCs from an aged source may have a greater propensity to favor a synthetic rather than contractile phenotype in vitro. 50 This would be beneficial in terms of de novo ECM production. Further study will be required to assess the potential of human VSMCs from an aged population to grow and respond phenotypically in culture.
Control of autologous cell populations for the development of an engineered aortic valve is a key problem in the field. Overall, the culture conditions proposed by this study have yielded a population of VSMCs with desirable phenotypic and functional characteristics as a surrogate cell source for VICs. Specific culture conditions provide an easy method of controlling in vitro conditioning of the valve before implantation. The treatment described in this study elicits favorable effects on proliferation, ECM gene expression, and phenotype of VSMCs for aortic valve tissue engineering and supports the hypothesis that VSMCs are a physiologically relevant, autologous cell source for valve tissue engineering and replacement.
Funding
This work was supported by The Heart and Stroke Foundation of Ontario (Grant number: NA5843).
Footnotes
Acknowledgments
The authors wish to thank Dr. Frank Beier and Vasek Pitelka for their assistance with this study.
Disclosure Statement
No competing financial interests exist.