
Abstract
The nucleus pulposus (NP) of the intervertebral disc in cattle and humans shows the most dramatic changes with aging of any cartilaginous tissue. In humans, notochordal cells disappear from the NP and are replaced with chondrocytic cells by adolescence. However, notochordal cells of the NP persist into adult life in some species, such as rats and rabbits. Therefore, comparison of the metabolic activity of notochordal and nonnotochordal cells is considered to be important for determining the type of cell to use for transplantation to regenerate intervertebral discs. In this study, we investigated the notochordal NP cells of rats and rabbits, as well as nonnotochordal (chondrocyte-like) bovine NP cells, in a three-dimensional culture system to examine whether proteoglycan metabolism varied among these three cell types. As a result, bovine NP cells produced around 0.18 mg/mL of glycosaminoglycan after culture for 5 days, while rat and rabbit NP cells produced about four and two times more glycosaminoglycan than bovine cells, respectively. In conclusion, this study demonstrated marked differences of energy metabolism and production of matrix components between notochordal and nonnotochordal NP cells. Animals with notochordal cells in the NP, such as rats and rabbits, may not provide good models for investigation of biological repair and tissue engineering for human disc disorders.
Introduction
There is now an increasing interest in developing biological repair methods for the treatment of disc degeneration that achieves the correct biomechanical properties for a successful outcome. Many researchers have performed basic studies on regeneration and transplantation of intervertebral discs that have applied tissue engineering using cell culture methods.21–29 So far, rat,30–32 rabbit,33–40 porcine,41,42 canine,43–45 ovine,46,47 and bovine48–51 discs have been used in previous studies as tissue sources. However, the effects of aging and degeneration on the metabolism of human discs have been less frequently studied because suitable human discs for in vitro experiments are difficult to obtain.52,53 Therefore, comparison of metabolic activity between notochordal and nonnotochordal cells is considered to be important for determining the type of cell to use for transplantation when attempting to regenerate intervertebral discs. In this study, we used notochordal cells from the NP of rats and rabbits, as well as chondrocyte-like bovine NP cells. We examined the differences of morphology and glycosaminoglycan (GAG) production between these notochordal and nonnotochordal NP cells.
Materials and Methods
Experiments were carried out under the guidance of the local animal ethics committee in accordance with the guidelines on animal experimentation of our university, the Japanese government animal protection and management law, and the Japanese government notification on the feeding and safekeeping of animals. Experiments were conducted on 60 adult male Sprague-Dawley rats (Clea, Tokyo, Japan) aged 6 weeks with a mean body weight of 205 ± 13.1 g (mean ± standard deviation) and 30 adult male Japanese white rabbits (Ninox Lab. Supply, Ishikawa, Japan) aged 3 months with a mean body weight of 1.45 ± 0.23 kg. These animals were killed, and the lumbar spine was resected aseptically. Bovine caudal discs were obtained from 18- to 24-month-old steers within 2–3 h of slaughter at a local abattoir and were dissected aseptically. Discs from 20 tails were used for the experiments described here.
Preparation for morphological studies in vivo
First we investigated the morphological differences among rat, rabbit, and bovine discs by light and transmission electron microscopy. Histological and electron microscope specimens in vivo were prepared using every five animals. The rats and rabbits were fixed by transaortic perfusion with a solution of 4% paraformaldehyde and 2.5% glutaraldehyde in 0.1 M phosphate buffer (pH 7.4). The lumbar spines of rats and rabbits or the bovine coccygeal spine were removed and fixed by immersion in the same fixative for 1 week. Then the specimens were decalcified in 0.5 M ethylene-diamine-tetraacetic acid. Light microscopy specimens were embedded in paraffin and stained with hematoxylin and eosin (HE). The morphology of NP cells and the number of these cells per unit volume were compared between the three animal species. The number of cells per unit volume was counted in two lumbar discs from five animals of each species. In rats and rabbits, the L4/5 and L5/6 discs were used. In steers, the coccygeal 1/2 and 2/3 discs were used. A total of 100 serial cross sections were prepared that covered the central part of each disc. The number of NP cells was quantified by a real-time image analysis system (LUZEX FT, Nikon, Japan) under a light microscope with a 5× objective. As it was difficult to identify individual notochordal cells, the number of nuclei per 1 mm2 was counted in a region of the central NP of each disc. The nuclei of NP cells have a diameter of about 5 μm, so 5-μm thick sections were observed every third section to avoid counting the same nucleus more than once. A total of 50 sections of each disc were observed to count the number of NP cells per 0.5 mm3, which was then doubled to obtain the number of cells per unit volume. The mean number of NP cells per unit volume was compared between animal species. Electron microscopy specimens were rinsed in 0.05 M Tris–HCl buffer, postfixed at room temperature for 3 h in 2% OsO4 in 0.1 M sodium cacodylate buffer, impregnated with 2% uranyl acetate, dehydrated in a graded ethanol series, and embedded in epoxy resin. For light microscopy, 1- to 3-μm thick sections were stained with toluidine blue. Ultrathin sections were stained with uranyl acetate and lead citrate for examination by electron microscopy (Hitachi-H-7000, Tokyo, Japan).
Cell isolation
NP was removed from six lumbar discs of rats and rabbits using a scalpel, and the pooled tissue was digested enzymatically by incubation for 18–20 h at 37°C in an incubator under 95% air and 5% CO2. The NP was removed from the upper five intervertebral discs of each bovine tail, and the pooled tissues were digested similarly. In one experiment, NP tissues were obtained from the lumbar spines of 10 rats and 5 rabbits, and from 3 bovine tails, after which cells were isolated and cultured repeatedly under the same conditions. The digestion medium consisted of Dulbecco's modified Eagle's medium (DMEM: cat. no 22320–022 with 25 mM HEPES, 1 mM sodium pyruvate, 1000 mg/L glucose, Invitrogen, Carlsbad, CA) containing 1 mg/mL of collagenase-1 (Sigma-Aldrich, Poole, Dorset, United Kingdom), and antibiotics/antimycotics (500 units m/L penicillin G, 500 μg m/L streptomycin sulfate, and 25 μg m/L amphotericin B (Life Technologies, Paisley, Scotland). The medium was adjusted to 400 mOsm with NaCl. After incubation, the suspensions were initially filtered through a coarse filter to remove undigested tissue and then through a 20-μm pore cell strainer (Sigma-Aldrich). The cells in the filtrate were washed three times by centrifugation (2500 rpm for 5 min), and were resuspended in DMEM. The cell suspension was assessed manually for viability and cell numbers by using a hemocytomer and trypan blue dye exclusion. Only cell preparations with a viability >95% were used.
Cell culture
Cells were encapsulated in alginate beads as follows. Briefly, washed NP cells were resuspended uniformly in 1.2% low viscosity alginate (Fluka Biochemika, Gillingham, Kent, United Kingdom) at a controlled density by gentle pipetting. Beads were then formed by expressing the cell suspension through a 21-gauge needle attached to a 5-mL syringe into a 102 mM CaCl2 solution. The beads were washed twice with 25 mL of 0.9% NaCl solution and washed twice more with 25 mL of DMEM, after which the beads were carefully placed into 48-well culture plates (five per well). In each separate experiment, under each condition tested and at each time point, three wells were used for biochemical analysis, three wells were employed for measurement of the sulfate incorporation rate, and one well was used for morphological examination of the beads.
The multiwell plates were cultured for 2 and 5 days at 37°C in DMEM containing 6% fetal bovine serum, 0.5% antibiotic/antimycotic, and 0.5% gentamycin (Life Technologies) in 370-mOsm medium under 5% CO2/95% air. Medium osmolarity was adjusted by addition of NaCl and was monitored by using a freezing point osmometer (Semi-micro osmometer, Knauer, Germany). The medium was changed after 24 h and its lactate content was measured.
After incubation, for each condition examined and at each time point, the beads from three wells were removed, weighed, and dispersed in three volumes of citrate buffer containing 5 mM cysteine hydrochloride and 0.56 units/mL of papain (Sigma-Aldrich). The beads from another three wells per experimental condition were labeled with 35 S-sulfate (Amersham Biosciences, Little Chalfont, United Kingdom) for 4 h. Cell density and viability were measured manually in 20-mL samples of the resulting solution by using a hemocytomer and trypan blue dye exclusion. The number of cells per bead was determined for calculation of lactate production, the GAG concentration, and the sulfate incorporation rate per million cells. The remaining solution was heated at 67°C overnight in a sealed tube to digest proteoglycans (PGs). Aliquots of the solution were then used to measure the total GAG content and the sulfate incorporation rate. The cell viability profile was examined in representative beads from the remaining well, while other beads from this well were fixed, sectioned, and examined morphologically.
Lactate dehydrogenase assay
Lactic acid production was used as a marker of cell metabolism because glycolysis is the major pathway of ATP generation for chondrocytes.54,55 Lactic acid production was measured with a commercial kit (Sigma-product no. 735–10) using lactate dissolved in DMEM as the standard.
Sulfate incorporation rates
To determine the sulfate incorporation rate at each time point, beads were resuspended in 0.5 mL DMEM with 5 μCi m/L of 35 SO42− (Amersham Biosciences) and incubated for a further 4 h at 37°C and 5% CO2. Radiolabeling was stopped by washing the beads twice in ice-cold (4°C) phosphate-buffered saline supplemented with 2 mM CaCl2 and 5 mM Na2SO4, followed by freezing at −20°C. Then labeled alginate beads were defrosted and digested in 0.5 mL of phosphate-buffered saline and 0.5 mL of citrate buffer, after which the solution was dialyzed extensively to separate bound and free 35 SO42−. The dialysis fluid and tubing were placed into a scintillation vial, and 4 mL of scintillant (LSC-5100; Liquidscint, Aloka, Japan) was added. The radioactivity of the dialysate and 20-μL aliquots of the incubation medium were measured with a beta scintillation counter. Then sulfated GAG synthesis was calculated as described previously by assuming that the specific activity of the 35 SO42−sulfate was the same in the medium and in the incorporated GAGs.
GAG content
The GAG content was measured by a modified dimethylmethylene blue assay. 56 The absorbance of aliquots of digested beads added to dimethylmethylene blue buffer (Serva Feinbiochemica, Heidelberg, Germany) was read at 595 nm with a UV/VIS spectrophotometer (UV-160A; Shimazu, Kyoto, Japan). GAG concentrations were estimated from a standard curve that was obtained with chondroitin sulfate (Sigma-Aldrich) in 0.1% alginate in citrate buffer.
In vitro morphological studies
In five separate experiments on the three species, beads were examined by light and electron microscopy. Beads were fixed by immersion in 2.5% glutaraldehyde (0.15 M cacodylate buffer, pH 7.2) with 2 mM CaCl2 and 5 mM Na2SO4 for 4 h at 4°C. After fixation, the light microscopy specimens were embedded in paraffin and stained with HE. The other specimens were postfixed in 1% OsO4 in 0.1 M sodium cacodylate buffer, impregnated with 2% uranyl acetate, dehydrated in a graded ethanol series, and embedded in epoxy resin. Then ultrathin sections were stained with uranyl acetate and lead citrate and were examined under a HITACHI H-7000 electron microscope.
Statistical analysis
Unless otherwise stated, data are presented as the mean ± standard error of the mean for at least five separate experiments (n ≥ 5). Each experiment was carried out in triplicate. The significance of differences was determined by using two-way analysis of variance with repeated measures. When the interaction was positive, we used Scheffe's test. Date were entered into a database and analyzed by SPSS statistical software (version 14.0.J; SPSS, Chicago, IL). A probability value of ≤0.05 was considered to be statistically significant.
Results
Morphological features of rat, rabbit, and bovine discs
Cross sections of the intervertebral discs of rats (Fig. 1A), rabbits (Fig. 1B), and steers (Fig. 1C) showed that a soft NP was surrounded by a firmly elastic annulus fibrosus with a lamellar structure, and the NP was situated approximately at the center. In rats and rabbits, the NP was gelatinous, viscous, fluidic, and extremely fragile. On the other hand, the bovine NP was soft, elastic, and not fluidic. In HE-stained sections of rat NP (Fig. 1D) and rabbit NP (Fig. 1E), cells containing a number of vacuoles, which are characteristic of notochordal cells, were observed and the boundaries between the cells were unclear. In rats, the NP only contained notochordal cells. In rabbits, the NP contained a number of cell clusters about 30–50 μm in diameter (each of which had from 2 to more than 10 cells) in mucoid material. On the other hand, bovine NP cells were scattered discretely and irregularly in a cartilaginous matrix, and each cell was surrounded by a small cavity with a pit-like structure (lacuna) (Fig. 1F). In the bovine NP, only chondrocyte-like cells about 10 μm in diameter were observed, and no notochordal cells were seen unlike the discs of rats and rabbits. The number of cells per unit volume in the rat, rabbit, and bovine discs was 82,251 ± 2676.3, 86,186.4 ±3360.9, and 4264 ± 501.2 cells/mm3, so the number of NP cells was far lower in bovine discs (Fig. 1G).
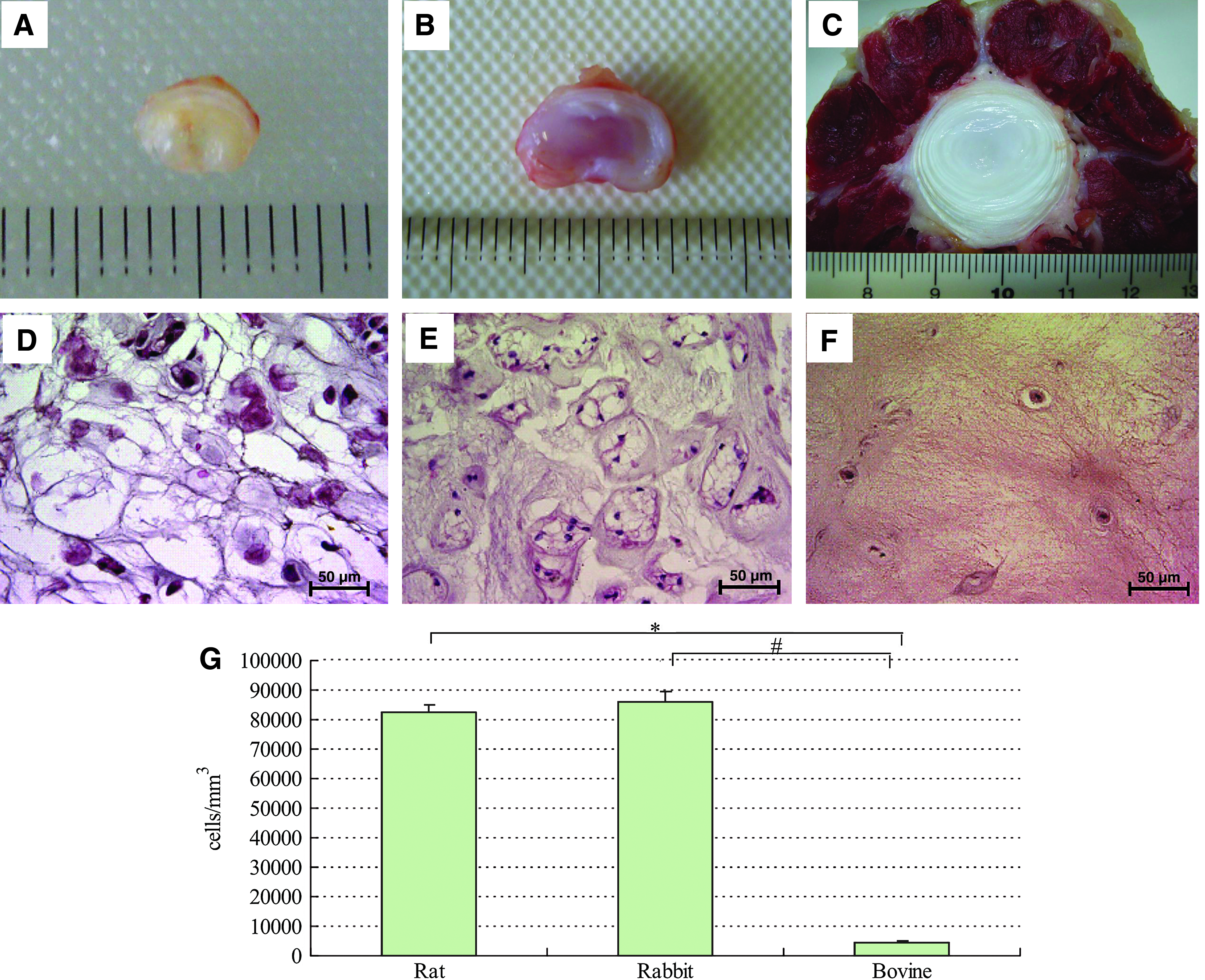
Photographs (
Transmission electron microscopy showed that the notochordal cells of the NP in rats (Fig. 2A, B) and rabbits (Fig. 2C, D) were markedly different from the chondrocyte-like bovine NP cells (Fig. 2E, F). The NP cells observed in rats and rabbits were connected to each other by tight junctions between cell processes or between soma and process, forming an intercellular network. A number of large or small vacuoles, which were empty or contained an amorphous substance, were observed, but no lacunae were found like those around the chondrocyte-like bovine NP cells. Notochordal NP cells and chondrocyte-like NP cells both contained a few organelles such as Golgi apparatus, rough endoplasmic reticulum, and mitochondria, but notochordal cells had more abundant vesicles and vacuoles within the endoplasmic reticulum when compared with chondrocyte-like cells.
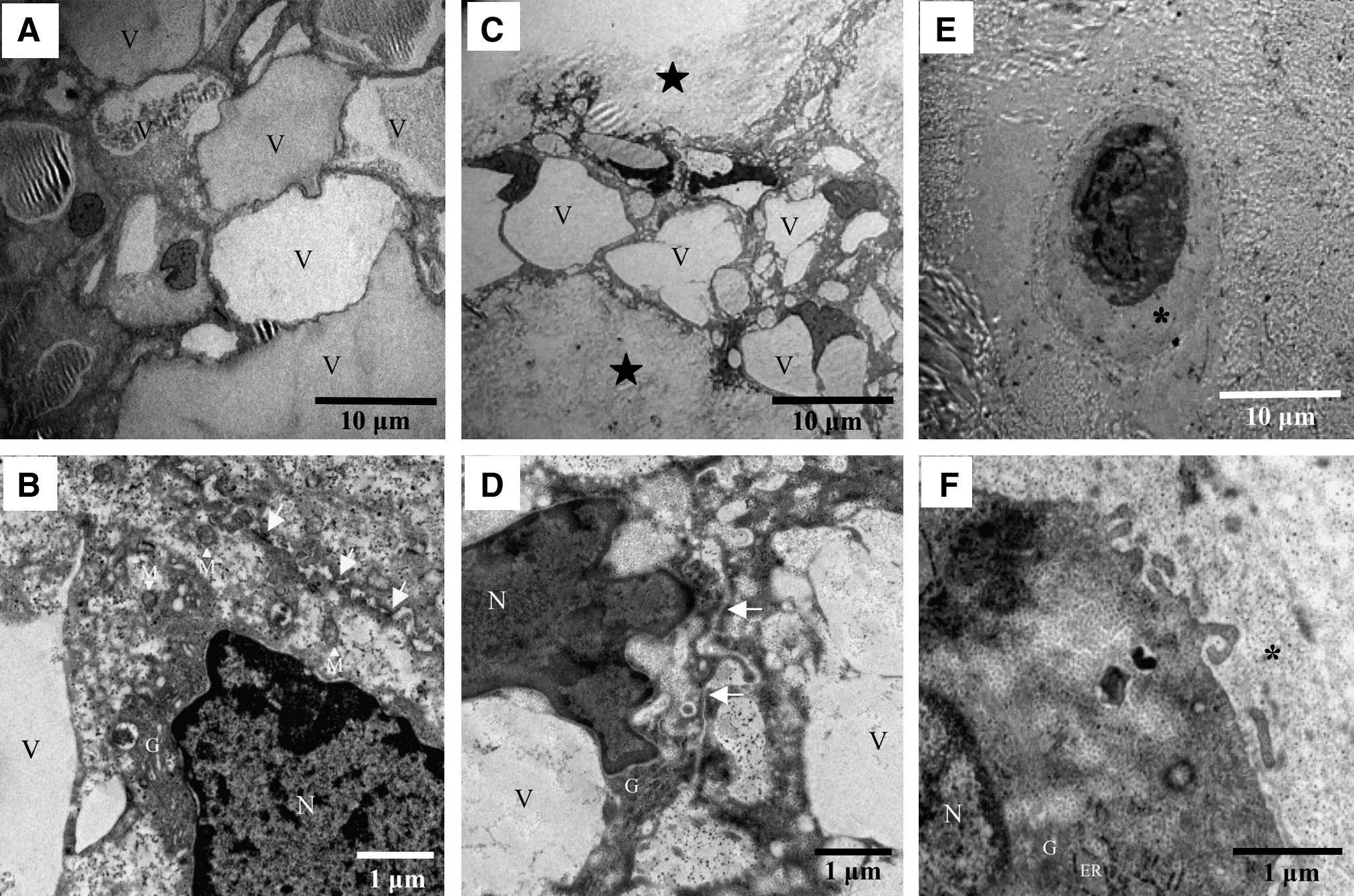
Transmission electron micrographs of the rat NP (
GAG production by cultured cells
The viability of NP cells was more than 90% after culture for 2 days or 5 days in each group. Figures 3 and 4 show pooled data from six separate experiments on cultured NP cells. Figure 3A shows lactate production by NP cells as a marker of total energy production. When lactate production per million cells was investigated, rat and rabbit NP cells produced much more lactate compared with nonnotochordal bovine NP cells. Lactate production decreased significantly over time as culture was continued (p < 0.05). Similarly, the rate of sulfate incorporation per live cell was the highest for rat NP cells and decreased over time as culture was continued (Fig. 3B), while it was the lowest for bovine NP cells. Sulfate incorporation decreased more steeply than lactate production during culture. While the total GAG content per mL of bead volume increased with the duration of culture for all the cells, it was highest for rat NP cells and lowest for bovine NP cells up to 5 days of culture (Fig. 4). On day 5, the total GAG content was 0.782 ± 0.087, 0.497 ± 0.034, and 0.177 ± 0.021 mg/mL of beads (mean ± standard error of the mean) after culture of rat, rabbit, and bovine NP cells, respectively. Rat and rabbit NP cells, respectively, produced about four to five and two to three times more GAGs than bovine NP cells after 5 days of culture. Thus, the notochordal NP cells were more active and produced significantly more GAGs than chondrocyte-like bovine NP cells.
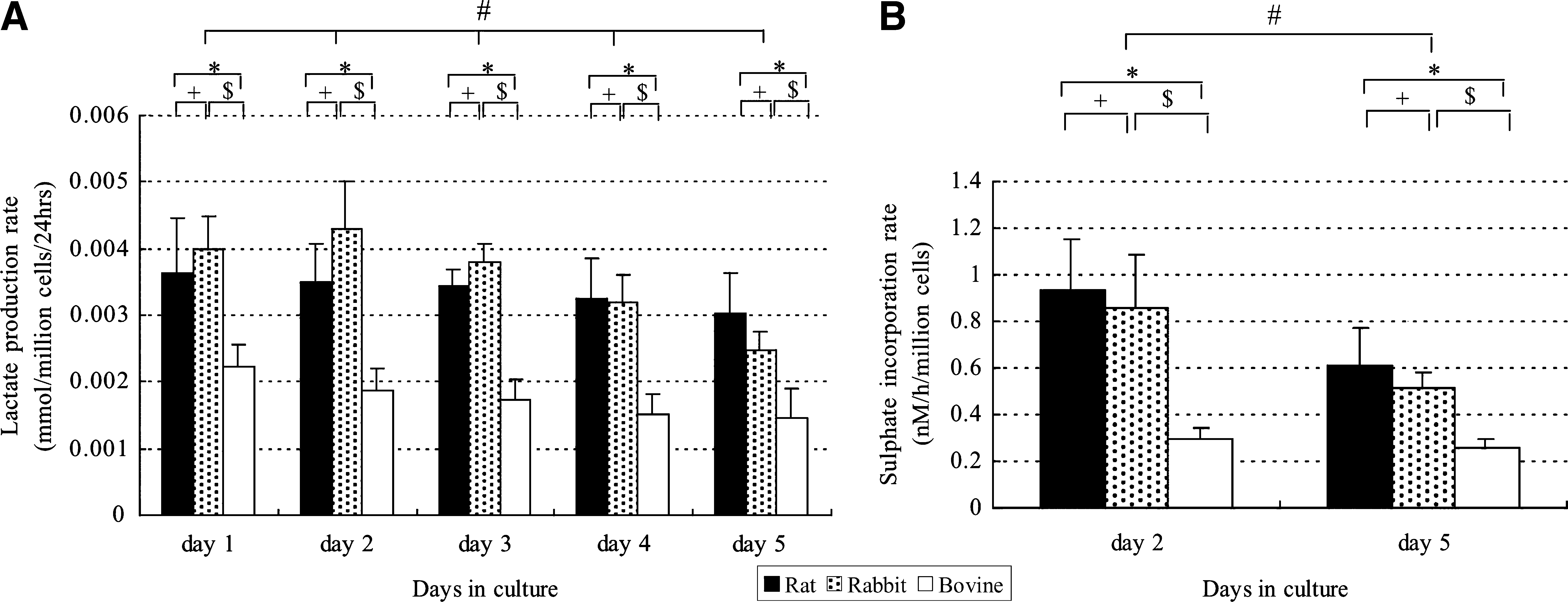
Metabolic activity of rat, rabbit, and bovine NP cells. (
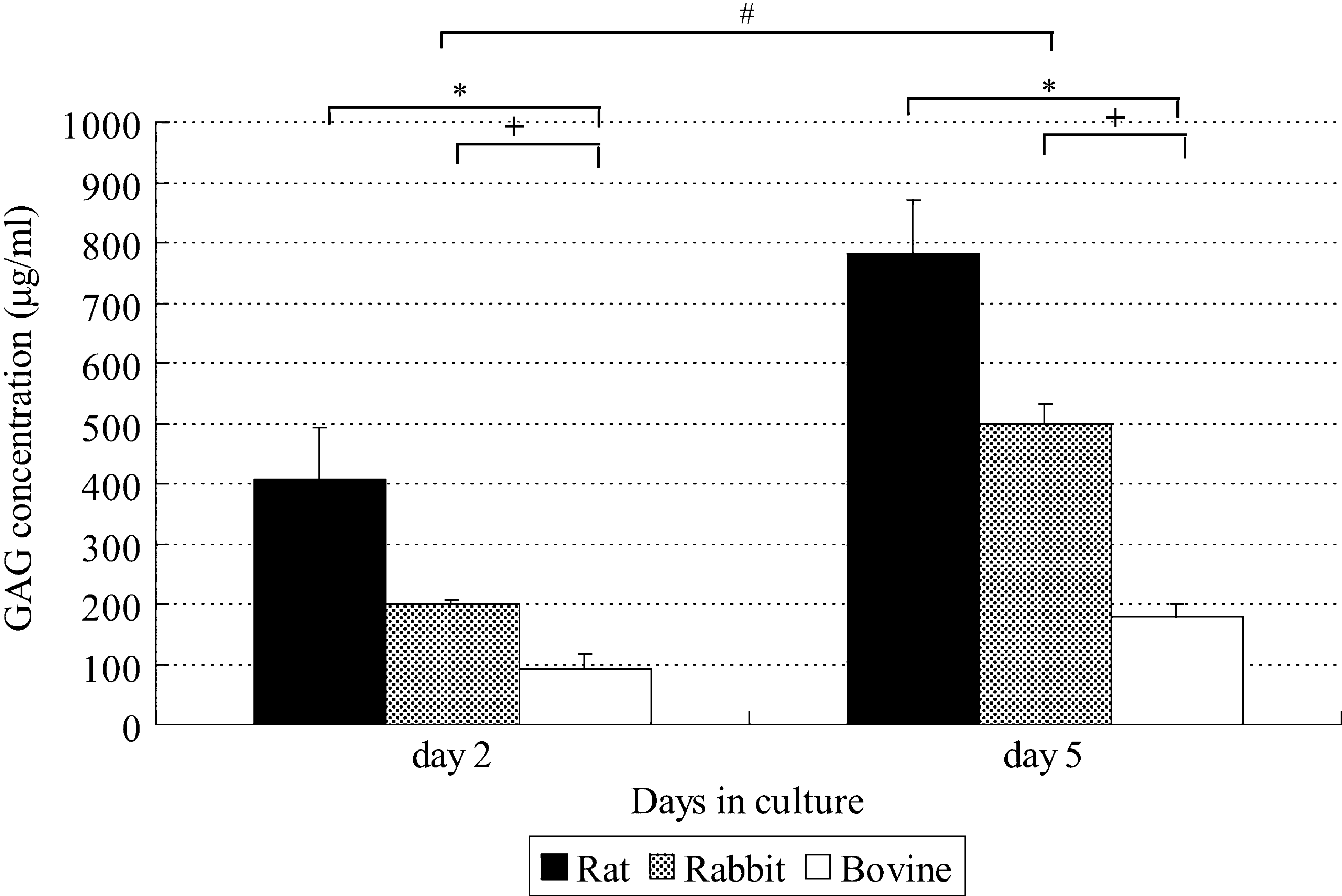
GAG concentration in rat, rabbit, and bovine NP cells after 2 and 5 days of culture. Cells were encapsulated in alginate beads and cultured in Dulbecco's modified Eagle's medium with 6% serum under air. The GAG concentration was measured after 2 and 5 days. Pooled data from six separate experiments are shown. GAG production per 1 mL of beads increased with time (#p < 0.05 by two-way ANOVA with repeated measures for 2 vs. 5 days). After 5 days of culture, the GAG concentration was lowest in the bovine cells. Values are the mean ± standard error. (p < 0.05 by the unpaired t-test for rat vs. bovine cells [*] or rabbit vs. bovine cells [+]). GAG, glycosaminoglycan.
Morphological features of cultured cells
When NP cells were encapsulated in alginate beads, cultured for 5 days, and examined under a microscope, notochordal NP cells of rats and rabbits were larger than chondrocyte-like bovine NP cells, and both cell types were surrounded by gaps (Fig. 5A, B, C). However, electron microscopy only revealed lacunae around the bovine NP cells and not around rat or rabbit NP cells, which contained a number of vacuoles (Fig. 5D, E, F). Both types of cell showed essentially similar findings to those observed in vivo and contained few intracellular organelles.
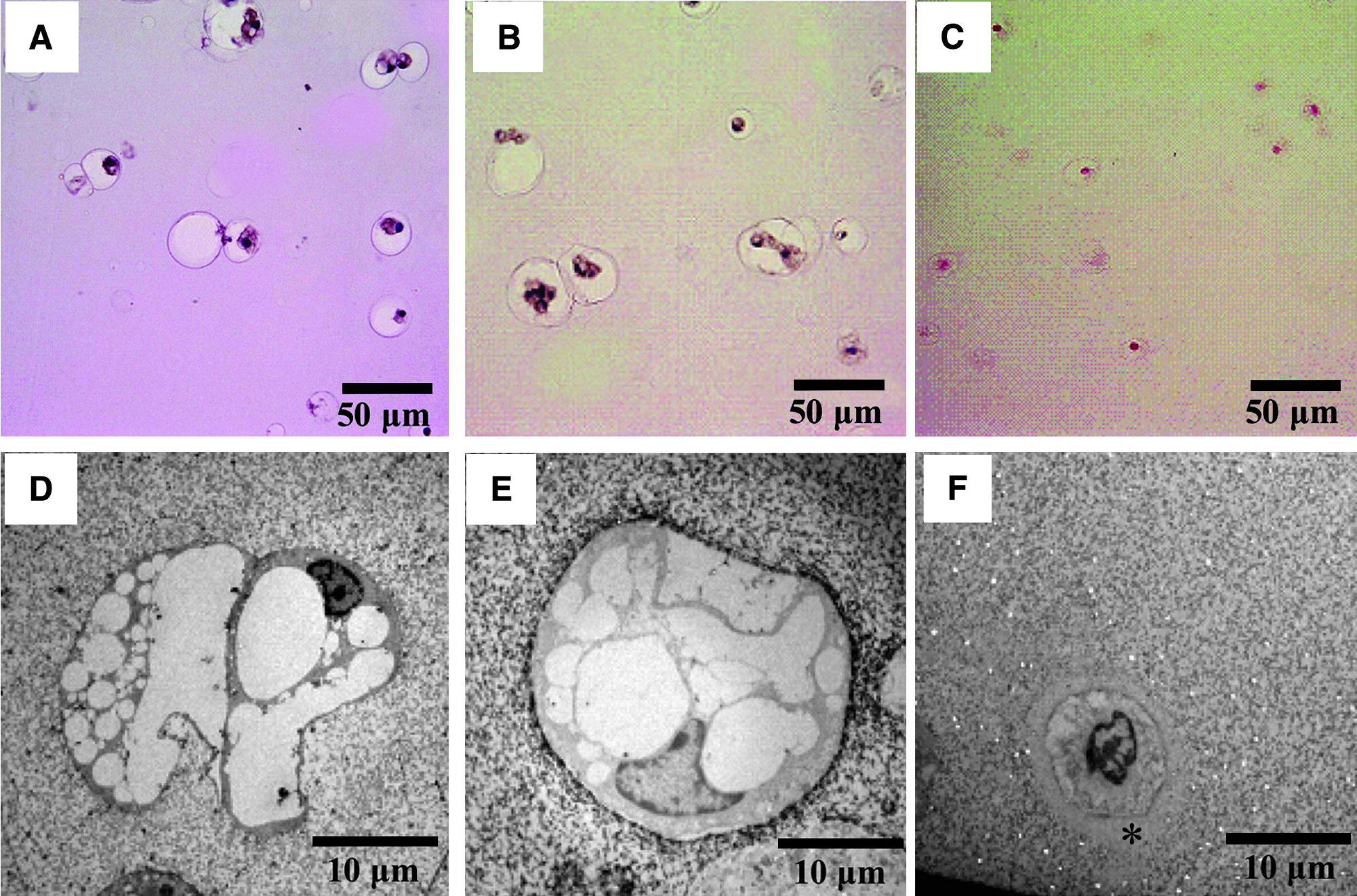
Photomicrographs (
Discussion
It is generally considered that the notochord gives rise to the NP during development of the intervertebral discs. However, it remains unclear why the cells composing the NP differ among animal species and stages of maturation and what role notochordal cells play in the growth process. In the present study, we found that the NP only contained notochordal cells in rats and rabbits, and these cells showed marked morphological differences from the cartilage-like cells observed in the bovine NP. When compared with a report by Trout et al.,57,58 who observed the human NP at various ages from fetal to 91 years by electron microscopy, the morphological characteristics of notochordal cells observed in rats and rabbits in this study were similar to those of NP cells observed in human discs up to adolescence, while the features of cartilage-like cells observed in bovine discs were similar to human NP cells from adolescence onward. That is, the notochordal discs contained physaliphorous cells aggregated in close contact with each other, while nonnotochordal discs contained cartilage-like cells scattered through the matrix and characteristically surrounded by lacunae, which were not observed around notochordal cells. The mechanism leading to the formation of lacunae and their significance remains unclear, but lacunae may reflect the influence of the cell over its immediate pericellular matrix, either through accumulation of excreted products or some other process. The formation of lacunae may also be related to changes of the NP that occur with age, including the apparent increase of some nutrients and substrates, making the extracellular matrix less accessible to cell products.59,60 Changes in the ability of substances to diffuse in or out of the cell and a reduction in the number of viable cells might gradually alter the extracellular matrix and thereby lead to age-related changes in the mechanical properties of the disc.
In this study, we determined the age of the animals based on their skeletal maturity so that NP tissues from different species could be compared under similar conditions. The age when skeletal maturity is reached is 2, 4–6, and 24–30 months after birth for rats,61,62 rabbits,18,63,64 and cattle, 65 respectively. Loss of notochordal cells occurs at 12 and 6 months after birth in rats61,62 and rabbits,18,63 respectively, while notochordal cells are completely replaced by cartilage-like NP at birth in cattle. 66 Accordingly, the discs from 2-month-old rats, 6-month-old rabbits, and 18- to 24-month-old cattle were considered to correspond approximately to the age of adolescence in humans. The cell density of the NP was noticeably higher in notochordal discs and was low (4264 cells/mm3 on average) in bovine discs, which was close to the value (about 6000 cells/mm3) reported for the human NP by Maroudas et al. 67 Butler and Smith 19 classified the NP cells into three stages: in stage 1 the NP consists entirely of notochordal cells; in stage 2 notochordal cells form clusters and the extracellular matrix increases; and in stage 3 notochordal cells show a marked decrease and extracellular matrix and cartilage-like cells become predominant. According to this classification, humans are already in stage 2 at birth and progress to stage 3 in their teens. The cells of the adult NP more closely resemble articular chondrocytes. The rat, rabbit, and bovine NP used in this study were considered to correspond to stages 1, 2, and human stage 3, respectively. Overall, the structure of bovine and human discs is similar, but some differences exist with regard to the diameter, height, and thickness of the wall of the annulus. 68 Although bovine tails support a lower load than human lumbar discs, the swelling pressure of bovine coccygeal discs has been shown to be similar to that of the discs in a person resting in the prone position.69,70 Biochemically, similar rates of in vitro PG synthesis 69 and matrix synthesis in response to hydrostatic pressure70,71 have also been found in human and bovine coccygeal discs. Both bovine and human discs are also similar with regard to the types and distribution of aggrecan and collagen.69,72 The bovine disc closely resembles the human disc because adult human and adult bovine NP have almost no notochordal cells and contain chondrocyte-like NP cells that produce a hyaline cartilage–like matrix. It has been demonstrated that notochordal cells persist throughout life in rats and rabbits, but decrease with growth, and are replaced by hyaline cartilage–like matrix and chondrocyte-like NP cells infiltrating from the annulus fibrosus.1,6,7,73 It is unknown, however, whether this decrease is caused by differentiation of notochordal cells into nonnotochordal cells, apoptosis, or lack of nutrients diffusing from the end plates. Thus, in each animal species, notochordal cells decrease and chondrocyte-like cells become predominant in the matrix of the NP with aging, but the reason for the decrease of notochordal cells and the origin of the chondrocyte-like NP cells remain to be elucidated. Therefore, investigation of the relationship between the decrease of matrix formation by notochordal cells and the onset of intervertebral disc degeneration is an important subject for the future.
PGs, particularly the large PG known as aggrecan, play a major role in load bearing by the intervertebral disc. Because of the high osmotic pressure due to the sulfated GAGs in aggrecan, it absorbs water that expands the collagen network and maintains tissue turgor.74,75 The stiffness of intervertebral disc tissues is thus strongly dependent on the aggrecan content.76,77 Thus, it is necessary to have an adequate content of PGs (GAGs) when attempting to produce intervertebral disc tissue with the mechanical strength to withstand stress of about 2.8–13.0 kN by tissue engineering. 78 The structure and composition of the intervertebral discs are intimately related to their function. By studying species differences and similarities, many researchers have attempted to define the functions of the discs. However, caution is necessary when interpreting and extrapolating to humans from such data, because even though all vertebrates have discs, not all discs are the same. For example, comparison of PG synthesis in the discs of chondrodystrophoid dogs (beagles) and nonchondrodystrophoid dogs (greyhounds) has revealed significantly lower PG synthesis in chondrodystrophoid dogs. 79 The relative risk of disc herniation is about 10 to 12 times higher in nonchondrodystrophoid dogs than in chondrodystrophoid dogs.80,81 Autoradiography with 35 S sulfate, a radio label that is largely incorporated by the GAG chains of PGs, has established that notochordal cells also synthesize PGs. 17 A similar finding has been reported for human fetal and pediatric discs in which notochordal cells are more abundant compared with mature discs.82,83 Aguiar et al. 84 have suggested that the notochordal cells found in the human NP up to approximately 10 years of age play an active role in NP development and in the maintenance of disc integrity through the production of soluble factors that induce NP cells to increase PG synthesis. Interestingly, it has been found that coculture of immature cells such as mesenchymal stem cells with notochordal cells and adult NP cells (chondrocyte-like cells) stimulates PG synthesis more rapidly, and this method has been extensively used in studies on disc regeneration.39,40,85,86 However, previous studies on PG production by notochordal cells and chondrocyte-like NP cells have only compared PG synthesis on the basis of sulfate synthesis, and no studies have compared GAG accumulated by these cells. In the present study, a three-dimensional culture system was employed to compare the metabolism of notochordal cells from rat and rabbit NP, as well as chondrocyte-like cells from the bovine NP. As a result, notochordal NP cells showed levels of lactate production and PG synthesis about twice as high as those of chondrocyte-like NP cells, indicating the high metabolic activity of notochordal cells. After culture for 5 days, accumulative GAG production was higher in notochordal NP cells than in cartilaginous NP cells, being about 156.5, 99.5, and 35.4 μg/mL/day for rat, rabbit, and bovine NP cells, respectively. The structure and composition of the intervertebral disc are intimately related to its function. By studying species differences and similarities, many investigators have attempted to define the functions of the intervertebral disc. However, when interpreting and extrapolating from such data or applying it to humans, it should be remembered that not all vertebrate discs are the same. Based on these results, it was considered preferable to study disc regeneration using discs from animals with chondrocyte-like cells resembling those in the adult human disc.
One index that is used for disc repair and regeneration is the GAG content of the disc. It has been reported that a 7–10% GAG content is required to obtain disc tissue with enough strength for clinical application. 87 Based on the results of the present study, it was calculated that it would, respectively, take at least 1, 2, and 5 years for rat, rabbit, and bovine NP cells to produce a 7% GAG content (i.e., 70 mg/mL). Accordingly, GAG accumulation by chondrocyte-like bovine and adult human NP cells is extremely slow compared with notochordal cells, and in vitro production of a construct with a 7% GAG content by using chondrocyte-like NP cells alone would be assumed to take an extremely long time and thus would be impractical. In fact, transformation of the matrix by cells in human discs is very slow, and studies have shown that PG turnover takes about 20 years 88 and collagen turnover takes more than 100 years. 89 However, autologous transplantation of cells with a high PG production capacity such as notochordal cells and mesenchymal stem cells may lead to successful repair of degenerated discs.39,40,84–86 Even if the transplantation of cells with a high PG production capacity into the NP is feasible, it is also essential for preventing the progression of disc degeneration that the transplanted cells survive in the degenerated disc and continue to produce appropriate macromolecules for maintenance of disc mechanical strength throughout life. The intervertebral disc is avascular, and the metabolic activity of its cells is regulated by various factors in the extracellular matrix, such as oxygen,54,90 osmolality,91–95 and pH.96,97 The cell density of the normal human NP is 2–4 × 106 cells/mL, and the extracellular environment differs markedly from that of other tissues, with an oxygen saturation of 1–5%, pH of 7.2–7.4, and extracellular osmolality of 370–400 mOsm (Fig. 6A). The supply of nutrients from vascular buds at the end plates to the NP of a degenerative disc is likely to be affected, causing the extracellular environment to deteriorate (Fig. 6B).87,98,99 When cells with a high metabolic activity such as notochordal cells are implanted at such a location, it is unclear that the cells will be able to obtain enough nutrients to achieve adequate regeneration of the extracellular matrix. In fact, notochordal NP cells have a higher metabolic activity than chondrocyte-like NP cells, and thus should require a larger amount of nutrients to maintain their metabolism than the chondrocyte-like cells.
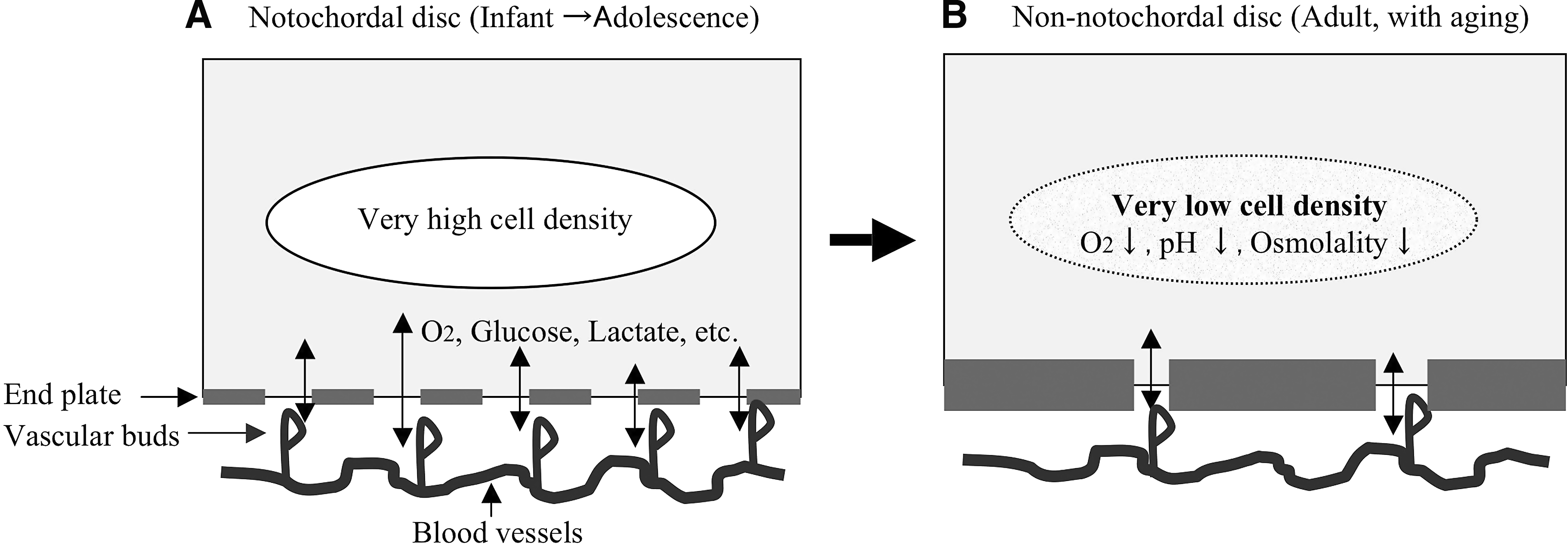
Changes of nutrition and the extracellular environment in NP with aging. The number of disc cells decreases with aging (
It can be assumed from the results of this study that the GAG content of the NP decreases when chondrocyte-like cells take over the major role of extracellular matrix production from notochordal cells during the growth process in humans. The immature disc shows a much higher cell density in vivo than the mature NP with chondrocyte-like cells, and lactate and GAG production were elevated when culture was done at the same cell density (4 × 106 cells/mL). This indicates that notochordal NP cells have a higher metabolic activity than chondrocyte-like NP cells and require more nutrients. 100 One of the initial signs of disc degeneration is said to be a decrease of PG, particularly GAGs, in the NP, and one possible cause may be a difference of metabolic activity between NP cells with different phenotypes. Although such changes are often considered to be signs of degeneration, these changes may merely be stages in the normal evolution of discs subjected to mechanical stress. Also, given that many degenerated discs are asymptomatic in humans,101,102 identification of the features that distinguish physiological from pathological degeneration is critical. That is, the decrease of the PG content in the NP that is observed in human discs during the growth process does not necessarily indicate pathological degeneration. Its major cause is considered to be a decline in the function of chondrocyte-like NP cells or the death of cells due to impairment of nutrient supply from the end plates, which occurs after notochordal cells are completely replaced by chondrocyte-like cells in the NP from adolescence onward. Therefore, it is very important to maintain an appropriate physicochemical environment to achieve successful disc repair by biological methods and tissue engineering procedures.
Footnotes
Acknowledgments
The authors would like to thank Mrs. Kyoko Shimokawa, Ms. Mika Osaki, and Ms.Yukiko Horiuchi for expert assistance in this study. Mr. Naruo Yamashita provided expert help with the photography. This work was supported by Grant-in Aid from the Ministry of Education, Science and Culture of Japan (21591896 and 20791027).
Disclosure Statement
No competing financial interests exist.