
Abstract
Objectives:
It has been well recognized that appropriate vascularization is emerging as a prerequisite for bone development and regeneration. The aim of this study was to test the hypothesis that locally applied granulocyte colony-stimulating factor (G-CSF) enhances bone regeneration via revascularization and osteogenesis.
Methods:
A segmental bone defect (20 mm) was created at the diaphysis of the rabbit ulna. The defects were treated with cationized gelatin hydrogel, which was the drug delivery system, with G-CSF, and then bone regeneration, neovascularization, and osteogenesis properties with G-CSF were assessed.
Results:
Radiographic, computed tomography, and histological findings revealed that bone formation was significantly promoted in G-CSF-treated group as early as 2 weeks. Immunohistochemistry, real-time reverse transcription-polymerase chain reaction, and flow cytometry studies indicated that angiogenesis/vasculogenesis, which are regulated by mobilization and incorporation of CD34+/G-CSF receptor (CSFR+) cells, and osteogenesis, which is regulated by osteocalcin+/G-CSFR+ cells, were also significantly enhanced in the G-CSF group.
Conclusions:
This study suggests that locally applied G-CSF contributes to an ideal local environment for fracture healing by supplying adequate blood flow and stimulating osteogenesis. G-CSF may have the therapeutic potential for bone regeneration.
Introduction
Recently, bone tissue engineering has attracted much attention as a new therapeutic technology that induces bone regeneration by making use of scaffolds, growth factors, cells, or their combination. 5 The majority of methods use mesenchymal stem cells (MSCs) and growth factors to mobilize, proliferate, and differentiate MSCs, which have proven to be effective for bone tissue regeneration. 6 In cytokine therapies, several growth factors, especially bone morphogenetic protein (BMP), fibroblast growth factor-2, insulin-like growth factor, and transforming growth factor-β, have been proven to be effective for bone tissue regeneration. BMP-2 and BMP-7 have been used in clinical studies.7,8
The reestablishment of vascularity is understood to be an early event in fracture healing and appropriate vascularization is emerging as a prerequisite for bone development and regeneration. It is reported that vascular endothelial growth factor (VEGF) has the potential to enhance bone healing via revascularization9–11 ; however, this finding has limited value as VEGF remains unavailable for clinical use.
Granulocyte colony-stimulating factor (G-CSF) has various functions including induction of proliferation, survival, and differentiation of hematopoietic stem cells (HSCs)/endothelial progenitor cells (EPCs), as well as mobilization of bone marrow cells.12,13 Although HSCs/EPCs and MSCs are perfectly distinct lineage cells and G-CSF is recognized to be mainly related with HSCs/EPCs,12,13 the interaction between G-CSF and MSCs remains unclear. Treatment with G-CSF has been reported to mobilize bone marrow stem cells into infarcted hearts and accelerate differentiation into vascular cells and cardiac myocytes. 14 G-CSF has also been proven to contribute to angiogenesis in an appropriate environment. 15 Similarly, G-CSF upregulates VEGF levels and stimulates neovascularization in patients with acute myocardial infarction. 16 It has been recently suggested that both intravenous and local application of G-CSF-mobilized CD34 cells and HSCs/EPCs enriched cell populations and promoted fracture healing via angiogenesis/vasculogenesis and osteogenesis.17,18 On the basis of these findings and as the main purpose of this study, we performed a series of experiments to investigate the hypothesis that locally applied G-CSF may contribute to the promotion of healing in bone defects via enhanced angiogenesis/vasculogenesis, which are regulated by the mobilization and incorporation of G-CSF receptor (CSFR+)/CD34+ cells, and enhanced osteogenesis, which is, in turn, regulated by osteocalcin+(OC+)/G-CSFR+ cells.
In the present study, we designed a biodegradable cationized gelatin (CG) hydrogel 19 with the ability to control the slow release of G-CSF. In this hydrogel system, growth factor physicochemically immobilized in the hydrogel can be released as a result of hydrogel degradation. 20 We investigated the regenerative effects of a locally controlled application of G-CSF on a critical-sized bone defect in a rabbit ulna in vivo.21,22
Materials and Methods
Preparation of CG hydrogels incorporating G-CSF
CG hydrogels were prepared according to a previously reported method.19,23 The CG hydrogels were designed to biodegrade for an average of ∼2 weeks in an in vivo condition 19 and G-CSF remained up to 40% by day 3 in our preliminary study (unpublished data). Briefly, gelatin was prepared through an acid process of pig skin type 1 collagen (Nitta Gelatin). Ethylenediamine and 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide hydrochloride salt were added into 250 mL of 100 mM phosphate-buffered solution (PBS) containing 5 g of gelatin at a molar ratio to the carboxyl groups of gelatin of 50. Immediately after that, the solution's pH was adjusted to 5.0 by adding 5 N HCl. The reaction mixture was agitated at 37°C for 18 h and then dialyzed against double-distilled water for 48 h at 25°C. The dialyzed solution was freeze-dried to obtain a CG.
An aqueous solution of 10 wt% CG was cast into a polytetrafluoroethylene mold and left at 4°C overnight for gelation. CG hydrogels were crosslinked in HCl–acetone (3:7, v/v) containing aqueous glutaraldehyde (Wako Pure Chemical Industries) solution (25 wt%). The crosslinking reaction was allowed to proceed for 24 h at 4°C, and the resulting hydrogel sheets were then immersed in 100 mM glycine aqueous solution at 4°C for 24 h to block the residual aldehyde groups of glutaraldehyde. The crosslinking hydrogel sheets were thoroughly washed with double-distilled water, freeze-dried, and sterilized with ethylene oxide gas. No change in hydrogel shape was observed before or after the freeze-drying and sterilization processes. The CG hydrogel sheets were cut into small squares (7 × 30 mm). Human recombinant G-CSF (Kirin) was added as drops onto the freeze-dried CG hydrogel squares and left overnight at 4°C to allow impregnation of the hydrogels. The total amount of G-CSF applied to one piece of hydrogel square is 5 μg/100 μL, and one piece of hydrogel square was applied to each single bone defect. Similarly, empty CG hydrogels with PBS were prepared as controls.
Animal experiments
All procedures were approved by the Animal Research Committee of Kobe University Graduate School of Medicine. Twenty-four skeletally mature Japanese white rabbits (Kitayama Labs) with a mean weight of 3.5 kg (3.2–4.0 kg) were used in the present study. Surgical procedures were performed according to a previously reported method.24–26 Briefly, rabbits were anesthetized by intravenous injection of 2.0–2.4 mL pentobarbital sodium solution at a dose of 30 mg/kg body weight. The rabbits were placed in the lateral position and surgeries were performed on the bilateral forearms. In each rabbit, the limbs were disinfected and a 4-cm-long anterolateral skin incision was used to expose the forearm. A 2-cm segmental bone defect was made in the midportion of the ulna with a surgical oscillating saw supplemented by copious sterile saline water irrigation. The osteotomies were made perpendicular to the long axis of the ulna. First, the defects were divided into two groups according to treatment as follows: the G-CSF group defects were filled with CG hydrogel with G-CSF (5 μg); the control group defects were filled with CG hydrogel with PBS. Further, the defects were divided into two groups and examined to investigate the effect of locally applied G-CSF as follows: the G-CSF + sFlt1 group defects were filled with CG hydrogel with G-CSF (5 μg) and then daily subcutaneous injection of 20 μg/kg sFlt1 (R&D Systems) was given for 14 days postoperation to block the effect of VEGF; the sG-CSF group defects were kept empty and CG hydrogel with G-CSF (5 μg) was applied into their back to examine the effect of systemic application of G-CSF. The bilateral forearms of the same animals were assigned the same group to avoid any overlap from the effect of systemic factors, G-CSF, to the opposite forearm and to mask the exact results of G-CSF. Fixation of the ostectomized bone was unnecessary because of the fibro-osseous union between the ulna and radius located distal and proximal to the surgical site. 25 The surrounding soft tissues were covered and the skin was sutured with 4-0 nylon in both groups.
After surgery, anteroposterior and lateral views of radiographs were obtained. Standardized exposure times and intensities were used and the positions were kept consistent. Further, high-resolution computed tomography (CT) was performed as described below. Then all rabbits were returned to their cages and allowed to move freely without immobilization. No evidence of postoperative infection at the wound site was observed, and all wounds were healed uneventfully. No secondary fracture of the operated forearm was observed. At 2, 4, and 8 weeks postoperatively, rabbits were euthanized with an intravenous injection of a fatal dose of pentobarbital sodium, and the operated forearm was disarticulated immediately from proximal to distal articulation.
Radiographic and CT analysis
Anteroposterior and lateral radiographic views of the disarticulated forearm were obtained in all samples (n = 8 in each group and at each time point), according to a previously reported method. Further, high-resolution CT (SOMATOM Emotion 6; Siemens) at 0.75 mm of thickness, 10 kV of energy, and 20 mA of intensity was performed to these samples, followed by three-dimensional reconstruction. The newly formed bone was quantified using syngo Volume Calculation software (Siemens). This evaluation was performed by an observer blinded to the group type and the postoperative interval (three times in each specimen) and was evaluated statistically.
Histological assessment of bone healing and morphometric evaluation of capillary density
After obtaining radiographs, four specimens in each group and at each time point were used for histological assessment. Each harvested tissue was embedded in OCT compound (Miles Scientific), snap frozen in liquid nitrogen, and stored at −80°C for histochemical and immunohistochemical staining as described below. The OCT blocks were sectioned along the radial plane, and 6-μm serial sections were collected on slides followed by fixation with 4.0% paraformaldehyde for 5 min and immediate staining.
For histological evaluation to observe endochondral ossification, 2-, 4-, and 8-week sections were stained with toluidine blue (n = 4 in each group).
Histochemical staining with fluorescent-conjugated isolectin B4 (Vector Laboratories) as an endothelial cell marker was performed at 2 and 4 weeks, and capillary density was morphometrically evaluated as the average value in five selected fields of soft tissue in the peridefect site (four sections in each sample and n = 4 in each group). 17 Capillaries were recognized as tubular structures positive for isolectin B4. DAPI solution was applied for 5 min for nuclear staining. The four among five selected fields were randomly obtained from near the cut ends of the defect, and the remaining field was obtained from the center of the defect. Two researchers, unaware of the identity of each group, evaluated the density of capillaries by counting capillaries at the defect.
Immunohistochemical staining
To detect G-CSFR, and various cell types at the bone defect area, immunohistochemistry (n = 4 in each group) was performed with the following antibodies: mouse anti-OC antibody (Takara Bio) for morphometric evaluation of osteoblast (OB) density, 27 mouse anti-CD34 antibody (Dako) for detecting EPCs/HSCs, 27 and mouse anti-human G-CSFR (Abcam PLC) to detect various cells related with G-CSF. The secondary antibodies for each immunostaining were as follows: Alexa Flour 594-conjugated goat anti-mouse IgG1 (Molecular Probes) for OC on OB density, Cy3-conjugated anti-mouse IgG (H + L) (Jackson ImmunoResearch) for CD34 staining, and fluorescein isothiocyanate (FITC)-conjugated anti-mouse IgG (Jackson) for G-CSFR staining. DAPI solution was applied for 5 min for nuclear staining.
OB density morphometrically evaluated the lining or floating cells positive for OC by the average value in four selected fields in the peridefect site. Double immunohistochemistry with OC and G-CSFR was performed to evaluate the existence of G-CSF-reactive OBs at the bone defects. Immunohistochemistry with CD34 was performed to detect the EPCs/HSCs at the bone defect area. The selected fields were randomly obtained from a new bone area near the bilateral cut ends of the defect. All morphometric studies were performed by two examiners blind to the treatment.
Real-time reverse transcription (RT)–polymerase chain reaction
To investigate angiogenesis and osteogenesis-related genes at the bone defect site, we used a real-time quantitative RT–polymerase chain reaction (PCR; Takara Bio). Briefly, total ribonucleic acid (RNA) from bone defects (n = 4; excluded from specimens for histological assessment) was obtained using Tri-zol (Life Technologies) according to the manufacturer's instructions at 1, 2, and 4 weeks after operation. The harvested tissues were obtained both from peripheral zones and center zone of the each bone defect (n = 2 in each zone). Approximately 1 μg of total RNA was reverse-transcripted using oligo (dT) primer, dNTP, 10 × PCR buffer, MgCl2, RNase inhibitor, and MulV Reverse Transcriptase (all from Applied Biosystems) from each sample. The converted cDNA (1 μL) samples were amplified in triplicate by real-time PCR (ABI PRISM 7700) in a final volume of 25 μL using SYBR Green Master Mix reagent at a final concentration of 1 × (Takara Bio).
The primer pairs of rabbit-specific genes were obtained by using a previously reported method 28 and were designed following rabbit-specific primers using Oligo software (Takara Bio). Levels of transcripts for 18S rRNA proved to be stable and were therefore elected as a reference gene for rabbit experiment. 29 Melting curve analysis was performed to ensure that only a single product was amplified, using Dissociation Curves software (Applied Biosystems). Amplified products were separated on 2.5% agarose gels and stained with ethidium bromide for observation. The observed PCR product bands were detected and the specificity of the reactions was confirmed by LAS-3000 mini (Fujifilm Co.). Results were obtained using sequence detection software (ABI PRISM 7700) and evaluated using Microsoft Excel. The following primers were used:
Rabbit G-CSF primer sequence: sense 5′-TCA GAG CAA CTG CGG AAG ATC-3′; antisense 5′-TCA GGG TGG CAC AGC TTG T-3′
Rabbit G-CSFR primer sequence: sense 5′-CAC CAG CCA CAG CTT GAA GA-3′; antisense 5′-GTT CAT ACT GGA CGG TCA TGG A-3′
Rabbit VEGF primer sequence: sense 5′-ATC ATG CGG ATC AAA CCT CA-3′; antisense 5′-CTC GGC TTG TCA CAT TTT TC-3′
Rabbit OC primer sequence: sense 5′-CCG GGA GGA GAT CTG TGA AA-3′; antisense 5′-CTG CCT TCT TCC ACA ATT TTA TCC-3′
Rabbit 18S primer sequence: sense 5′-CGG ACA CGG ACA GGA TTG AC-3′; antisense 5′-CCA GAC AAA TCG CTC CAC CA-3′
Enzyme-linked immunosorbent assay analysis
Peripheral blood (PB) was aspirated from the hearts of rabbits preoperation and at 1, 3, and 7 days postoperation (n = 3 in each group). Serum was collected and enzyme-linked immunosorbent assay (ELISA) was performed to measure the concentration of G-CSF in the PB. Briefly, the PB of the G-CSF, sG-CSF, and control groups was collected and centrifuged for 15 min at 3000 rpm, and then the supernatants were used. Commercial human G-CSF ELISA kits (R&D Systems) were used according to the manufacturer's instructions.
Flow cytometry studies and monoclonal antibodies
PB was aspirated from the hearts of rabbits preoperation and at 1, 3, and 7 days postoperation with PBS–fetal calf serum (n = 3 in each group). PB mononuclear cells (MNCs) were obtained by density gradient centrifugation at 400 g for 20 min with Histopaque-1083 (Sigma). The light-density MNCs were collected, washed twice with Dulbecco's PBS supplemented with 2 mM ethylenediaminetetraacetic acid, and counted manually. Regular flow cytometric profiles were analyzed with a FACS Calibur analyzer and CELLQuest software (Becton Dickinson Immunocytometry Systems) as previously described.17,30 The following monoclonal antibodies were used to characterize the PBMNCs: phycoerythrin-conjugated anti-CD114 (G-CSFR) (BD Pharmingen), mouse anti-CD34 (Zymed), 31 FITC-conjugated anti-mouse IgG1 (BD Pharmingen), IgG1-phycoerythrin isotype controls (BD Pharmingen), IgG1-FITC isotype controls (BD Pharmingen), and PI (Sigma).
Statistical analysis
All values were expressed as mean ± standard error. The comparisons between the two groups were made using the one-way analysis of variance (ANOVA). A probability value of <0.05 was considered to denote statistical significance.
Results
G-CSF and G-CSFR expressions after operation
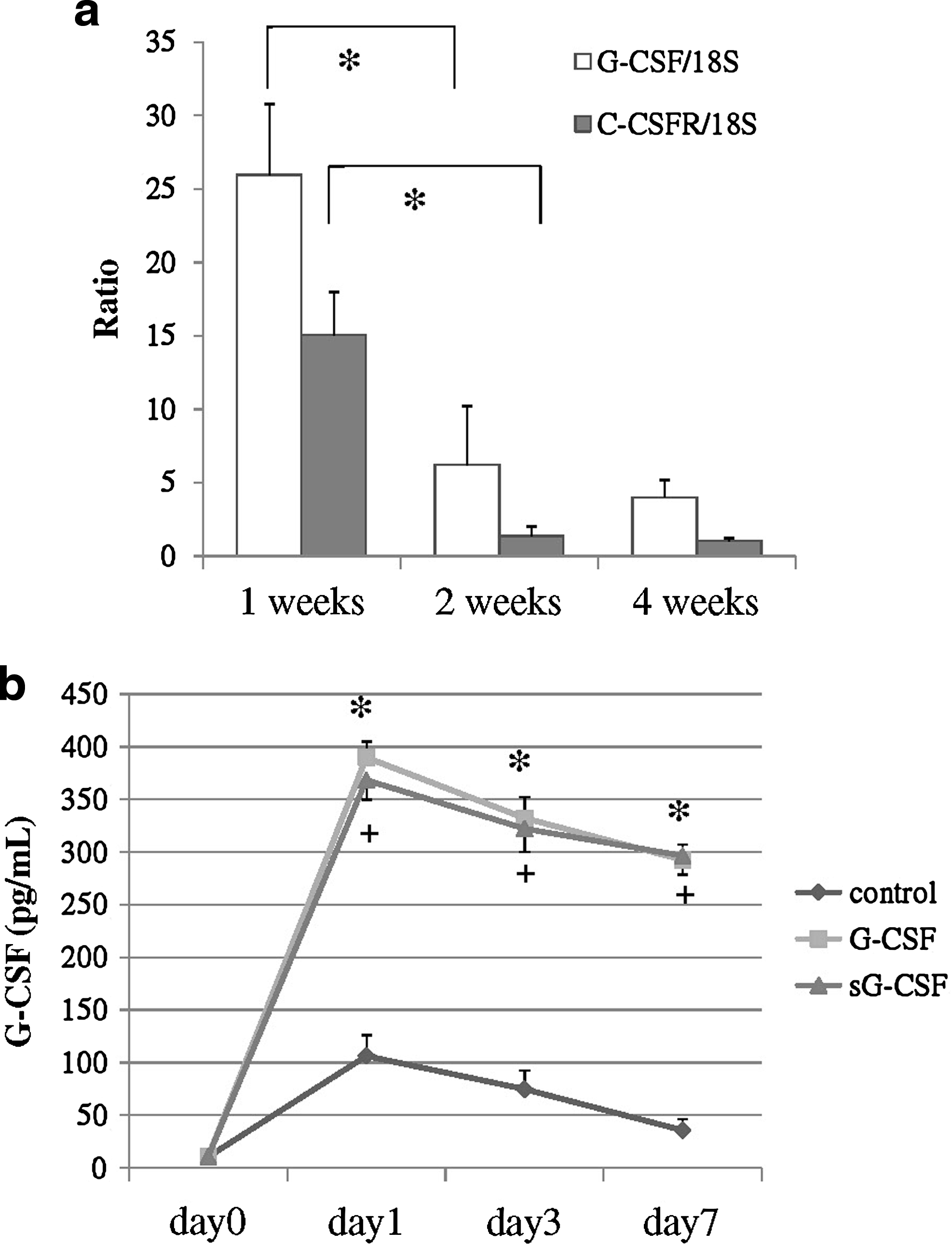
The natural course of G-CSF and G-CSFR expressions in this model was examined using real-time RT-PCR. The results showed that both G-CSF and G-CSFR mRNA expressions reached their peaks at 1 week and then decreased time dependently. Both G-CSF and G-CSFR mRNA expressions were over 10-fold upregulated, compared with the following weeks, with statistical significance (p < 0.05; Fig. 1a). These results showed that G-CSF and G-CSFR were expressed soon after operation in the natural course of this rabbit model. Analysis of G-CSF concentrations using ELISA revealed that G-CSF in PB was significantly elevated in the groups G-CSF and sG-CSF (Fig. 1b). This result revealed that both locally and systemically applied G-CSF elevated G-CSF concentrations systemically.
(
Locally applied G-CSF enhanced bone formation at the segmental bone defect by radiographic, CT, and histological assessments
Figure 2a–p shows X-ray photographs and CT of the bone defects during each week of the study. Bone regeneration at the defect was radiographically detected in the G-CSF-treated groups (groups G-CSF, G-CSF + sFlt1, and sG-CSF). Radiopacity was observed at the cut ends of the bone defect when treated with CG hydrogel only, but there was no progression of radiographic healing. Defect filling with new bone was seen in some G-CSF group samples at 8 weeks and some samples had healed with the cut ends of the cortex indistinguishable. In contrast, defect filling was not observed in any control group sample at 8 weeks. The results of quantitative evaluation with CT are shown in Figure 2q. The ratio of new bone area over time was statistically higher in the G-CSF group than in the control group at every time point (p < 0.05). The ratios of new bone area in the G-CSF + sFlt1 and sG-CSF groups were comparable with the G-CSF group at 2 and 4 weeks; however, the ratios at 8 weeks were significantly inferior to the G-CSF group (p < 0.05). These findings revealed that bone formation was promoted as early as 2 weeks in samples with locally applied G-CSF. Further, these results also revealed that bone formation by G-CSF was reduced by VEGF-blocking treatment and G-CSF treatment outside the bone defect.
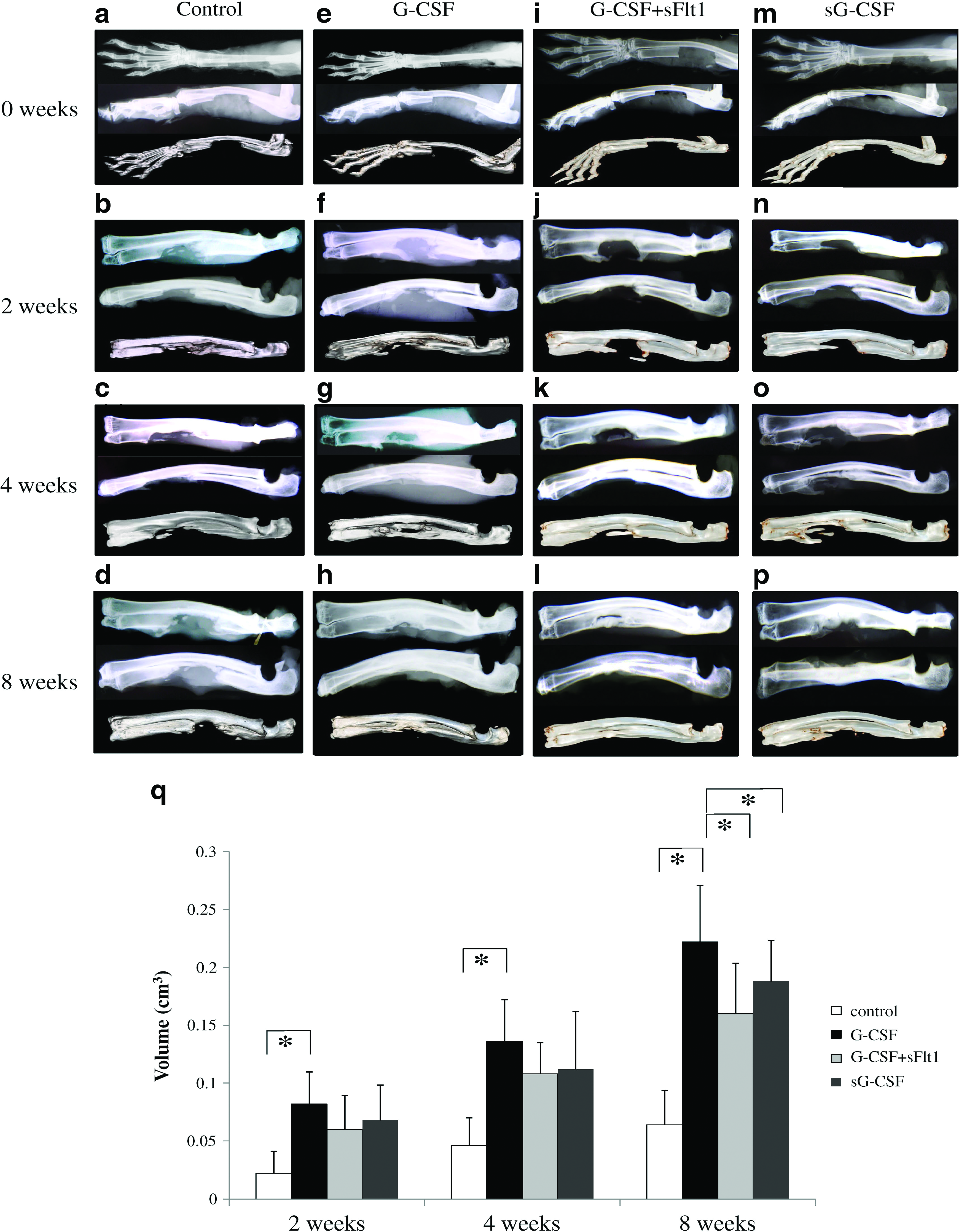
Promotion of bone regeneration by G-CSF. R&C at the segmental bone defect of rabbit ulna. The upper figures in each slide are radiographs with anteroposterior view, middle figures are lateral view, and lower figures are reconstructed CT. (
Figure 3a–g shows histological sections stained with toluidine blue at ulna bone defect at each week after application of CG hydrogel incorporating PBS (control; Fig. 3b–d) or CG hydrogel incorporating G-CSF (G-CSF; Fig. 3e–g). As early as 2 weeks, cartilaginous mass at the margins of the bone defect could be seen; it was especially visible around the cortical bone edge in the G-CSF group. At 4 weeks, obvious new bone formation and rich cartilaginous mass proliferation were seen from the margins of the bone defect in the G-CSF group. Some new bone formation existed around the host bone in the control group; however, lively cartilage proliferation was difficult to observe around the new bone area compared with the G-CSF group. At 8 weeks, in the G-CSF group, new bone formations from both sides had extended and some samples had edges that had bridged with each other. In contrast, in the control group, new bone and cartilage formation had disappeared and the edges of the new bone were clearly distinguished from the peripheral fibrous tissues. The defects were occupied with fibrous tissue or a hypocellular vacant area. These histological findings also revealed that bone formation was promoted when they were treated with locally applied G-CSF.
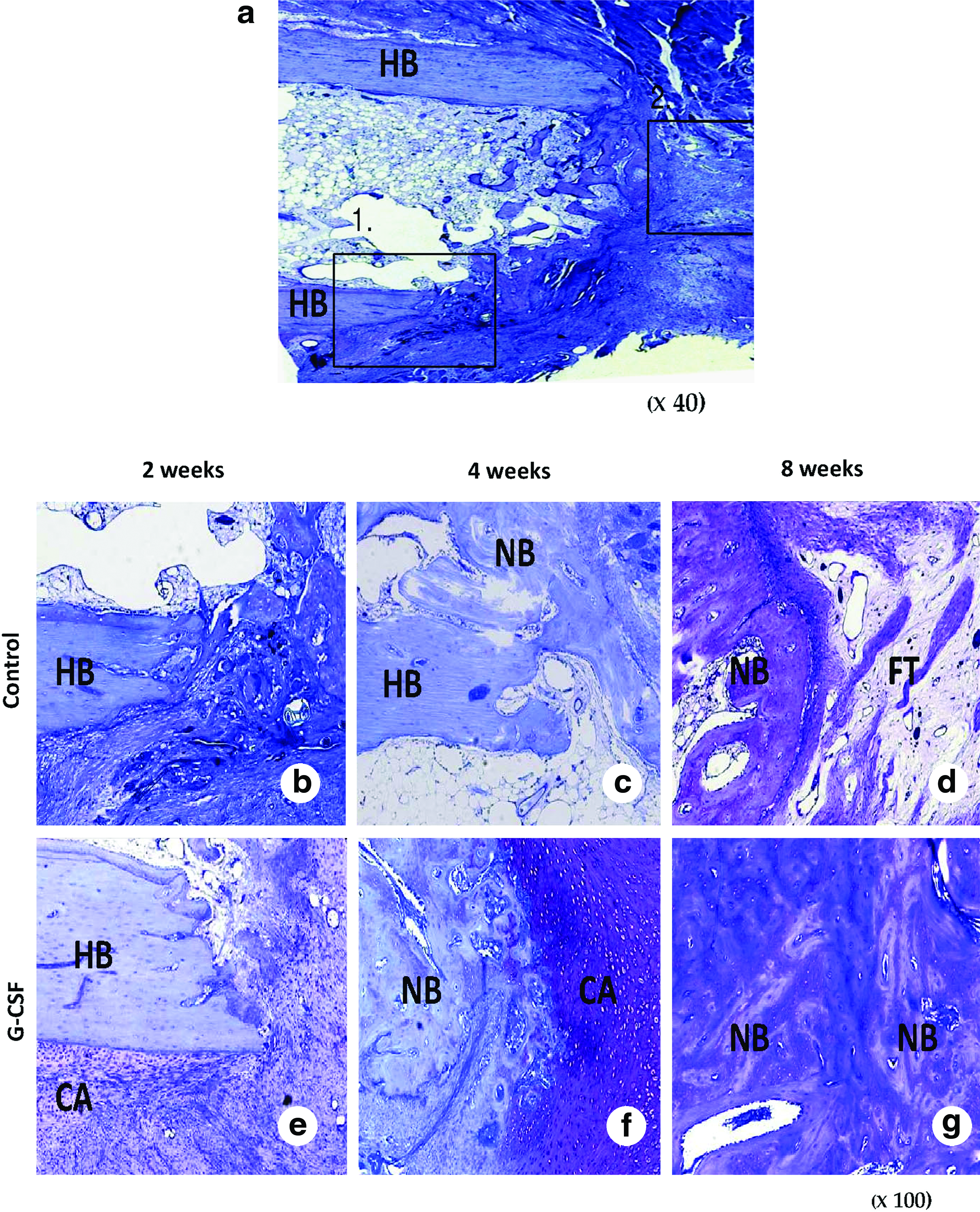
Photomicrographs of the bone defect with toluidine blue staining. (
Locally applied G-CSF promoted angiogenesis and vasculogenesis
First, to examine the angiogenesis by G-CSF treatment, we performed histochemical staining for isolectin B4 and real-time RT-PCR to evaluate VEGF mRNA expression. Figure 4a shows the low-power toluidine blue staining photograph illustrating a section of the high-power photograph. Histological sections to estimate angiogenesis, vasculogenesis, and inflammatory reaction were mainly and randomly obtained from different points (square 1 in Fig. 4a). Sections to estimate osteogenesis were also obtained from the new bone cavity point (square 2 in Fig. 4a). Histochemical staining for isolectin B4 showed that the capillary density was significantly increased in the groups G-CSF, G-CSF + sFlt1, and sG-CSF at 2 weeks, although the increment in the G-CSF + sFlt1 group was significantly inferior to those of the G-CSF and sG-CSF groups (Fig. 4b–e, j). No significant changes were seen again at 4 weeks (Fig. 4f–i, j). The results of angiogenesis-related mRNA expression are shown in Figure 4k. The results are reported as ratios compared with 18S rRNA expression. The results showed that VEGF mRNA expression had about a threefold increase in the G-CSF group as early as 1 week (*p < 0.05), and about a twofold increase in the G-CSF group at 2 weeks (*p < 0.05), although no significant changes were seen at 4 weeks. These results revealed that G-CSF increased angiogenesis at the bone defect in the early stages.

Angiogenesis by G-CSF. (
Next, to investigate mobilization into PB and incorporation into bone defect site of CD34+ cells and EPC-enriched cell population, we performed fluorescence-activated cell sorting (FACS) analysis of PB cells and immunohistochemical staining at the bone defect area. FACS analysis, showing the mobilization of CD34+ and G-CSFR+ cells into PB (Fig. 5a–d), revealed that both CD34+ and G-CSFR+ fractions in PB reached their peaks at 1 day postoperation and decreased gradually, and that CD34+ and G-CSFR+ fractions were significantly increased in the G-CSF group at 1, 3, and 7 days after operation (p < 0.05, n = 5; Fig. 5b, c). CD34/G-CSFR+ cell fractions also reached their peaks at 1 day postoperation, and the cell fractions significantly increased in the G-CSF group at 1, 3, and 7 days postoperation (p < 0.05, n = 5; Fig. 5d).
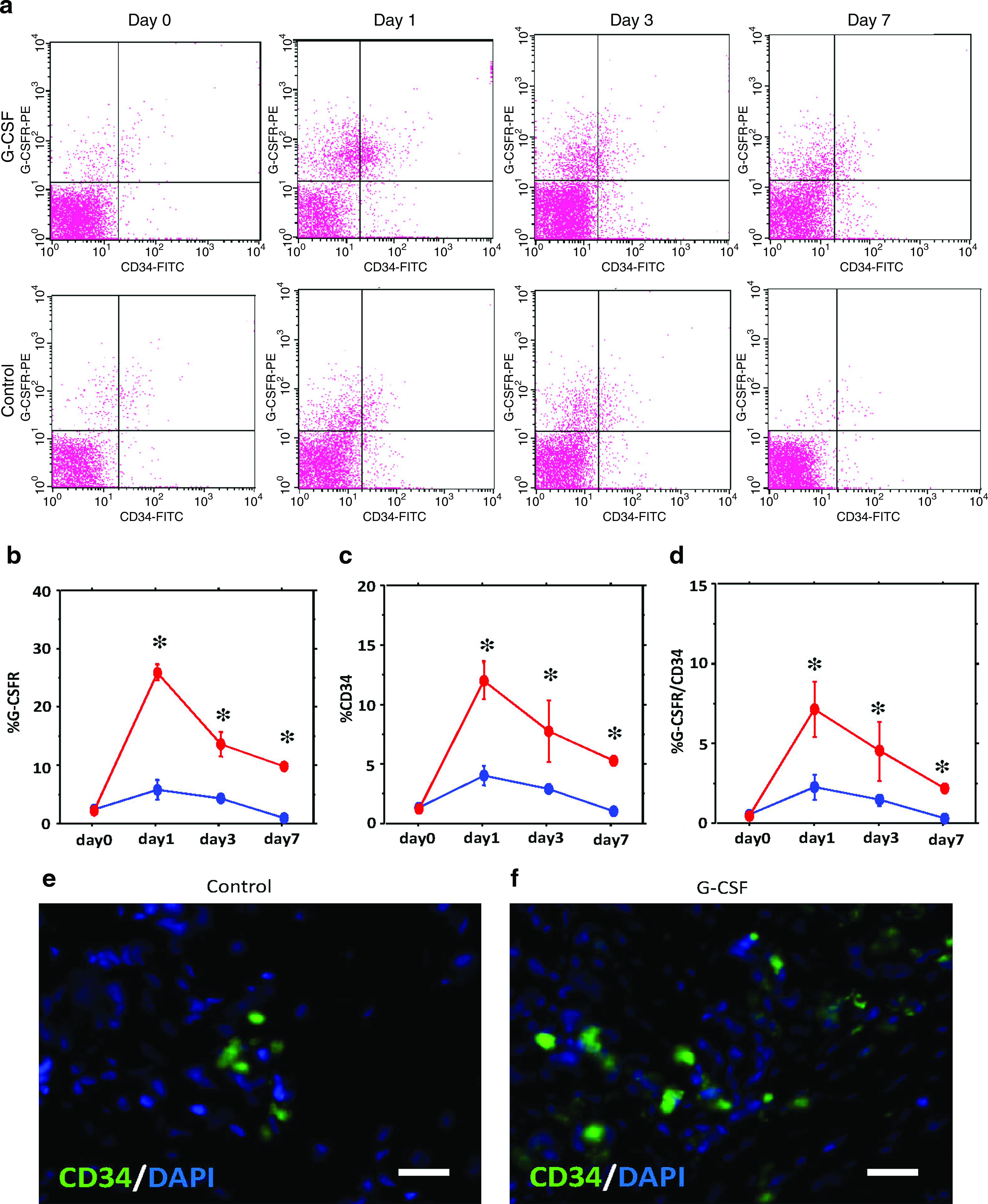
Vasculogenesis by G-CSF in rabbit bone defect model. (
Immunohistochemical staining of CD34 revealed that the bone defect in the G-CSF group expressed more CD34+ cells than the control group (Fig. 5e, f). Almost all CD34+ cells existed in the capillary structure, and not along the bone cavity, suggesting that CD34+ cells mainly promote neovascularization. These FACS and immunohistochemical staining results indicated that G-CSF mobilizes EPC into PB and incorporates into the bone defect.
These series of results strongly indicated that locally applied G-CSF promoted neovascularization via angiogenesis and vasculogenesis in the early stages at the bone site.
Recruitment of OB lineage cells
To examine osteogenesis at the bone defect area, we performed immunohistochemical staining for OC and real-time RT-PCR to evaluate OC mRNA expression. Further, to detect G-CSFR-positive OBs, double immunohistochemical staining for OC and G-CSFR was performed. Immunohistochemical staining for OC showed that OB density significantly increased in the G-CSF group at 2 weeks (Fig. 6a, b, e). Further, double immunohistochemical staining for OC and G-CSFR revealed that more G-CSFR/OC+ cells were incorporated into the bone defect area (Fig. 6c, d). Quantitative gene analysis revealed that OC mRNA expression experienced about a three-fold increase in the G-CSF group at 1 and 2 weeks (*p < 0.05), although no significant changes were seen at 4 weeks (Fig. 6f). These results indicated that G-CSF mobilized more OB at the bone defect area and incorporation of G-CSF-mediated OB was confirmed in the early stage at the bone defect site.
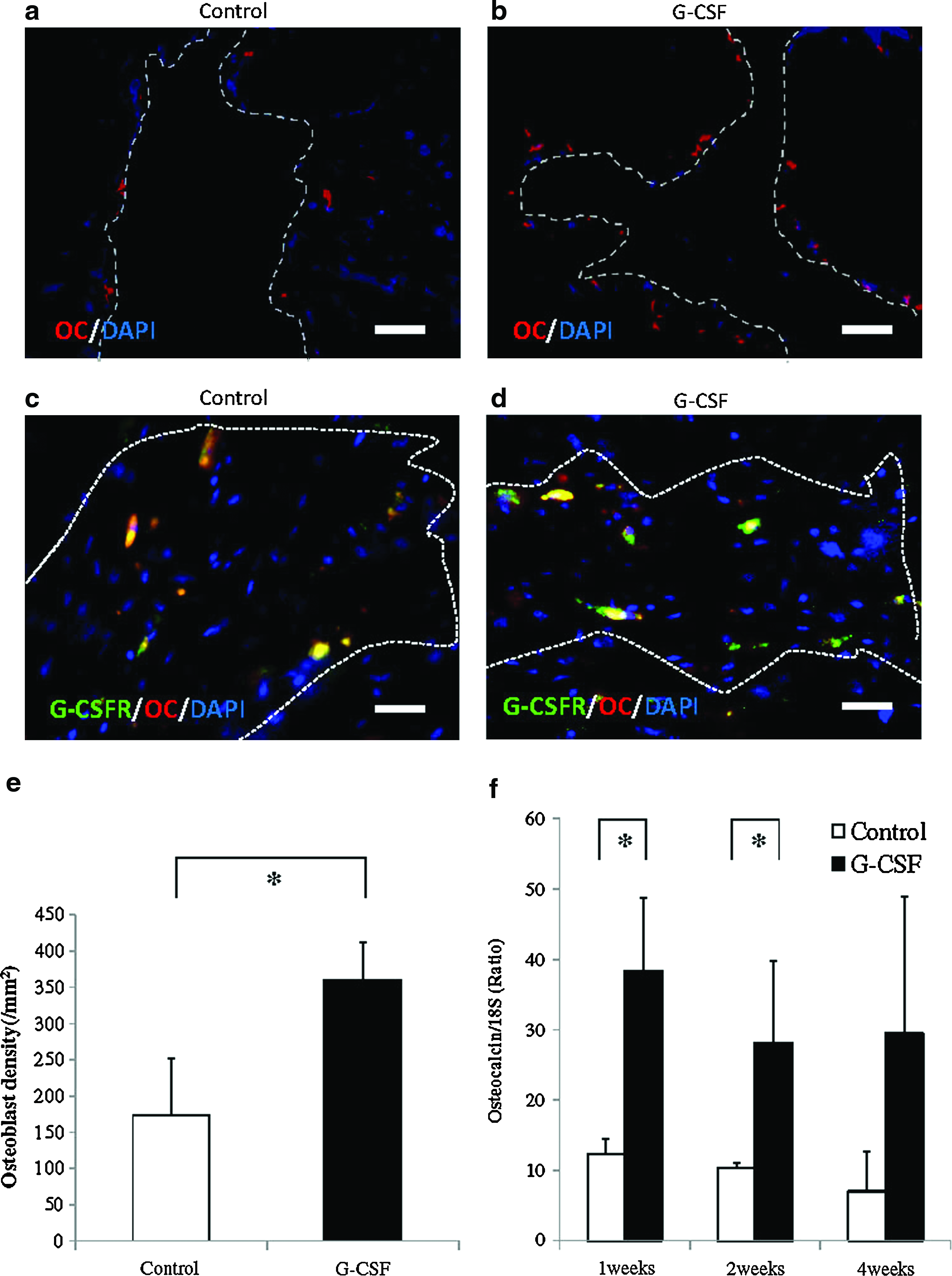
Osteogenesis by G-CSF immunohistochemical staining for OC at 2 weeks showing osteoblast formation (red, OC; blue, DAPI; magnification × 200; scale bar: 50 μm); (
Discussion
Many researchers have attempted to develop technologies and methodologies to facilitate stronger and faster repairing of bone tissues. Endochondral and intramembranous ossification, two distinct yet interactive responses ultimately responsible for bone healing, require an adequate blood supply and so efforts to enhance fracture neovascularization by supplementing recognized proangiogenic cytokines have yielded promising results.10,11,32 However, it remains unclear which cell types contribute to the development of the endochondral field and how the neovascularization process originates and is enhanced.
G-CSF is a hematopoietic growth factor produced by monocytes, fibroblasts, and endothelial cells,33,34 which enhances the proliferation and differentiation of neutrophil progenitor cells. G-CSF leads to an increase in bone marrow progenitor cells. Particularly, it prolongs the survival time of neutrophils, retarding the apoptosis of neutrophil.34–37 G-CSF is already used clinically in the treatment of bone marrow transplantations and secondary granulocytopenia after chemotherapy. 37 It is reported that the angiogenesis-promoting effect of G-CSF is partly mediated by enhanced recruitment of mature HSCs, and that G-CSF promotes neovascularization by releasing VEGF from neutrophils. 15 Ohki et al. further indicated that local administration of G-CSF into ischemic tissues in adults might be a promising treatment strategy by easily and effectively promoting collateral vessel formation by modulating the cellular and angiogenic growth factor setting in the muscle niche. 15 G-CSF has been reported to have not only an angiogenesis-promoting effect, but also to promote cardiomyocytes, neurons, and liver tissues regeneration, via bone marrow cell migration into injured organs.38–41 These reports suggested that G-CSF can mobilize bone marrow stem cells to various injured organs.
In this study, almost all defects in the G-CSF group achieved bone union at 8 weeks. Further, histological analysis revealed that cartilaginous mass formation could be seen at the margins of the bone defect as early as 2 weeks in the G-CSF group, and obvious new bone formation and rich cartilaginous mass proliferation were seen from the margins of the bone defect in the G-CSF group at 4 weeks. There have been a few reports associating G-CSF and fracture healing,42,43 but the mechanism for the healing was unknown. 42 Although Kaygusuz et al. revealed that G-CSF could prevent the delay of fracture healing by naproxen, by raising the transforming growth factor-β1 level in blood samples, 43 the pathophysiology of fracture healing by G-CSF remains unknown at present. To investigate the pathophysiology of bone promotion by G-CSF, we first examined the natural course of G-CSF and G-CSFR expressions in this bone defect model by using real-time RT-PCR. The results showed that both G-CSF and G-CSFR mRNA expressions remarkably increased at 1 week postoperation in our bone defect model. In general, the process of fracture healing started with an initial stage in which a hematoma was formed at the fracture site following vascular invasion as a result of an inflammatory response. In this step, mesenchymal or undifferentiated cells gathered and proliferated, resulting in the formation of reparative granuloma at the fracture site, called an external callus. 44 The results of our study suggested that G-CSF and G-CSFR expressed and contributed to bone healing in this initial stage in our model.
Although locally applied G-CSF could be expected to increase the concentration of G-CSF at the bone defect, the lives of many growth factors are often too short to sustain biological activity in vivo. This is also true for G-CSF, which should be administered by daily subcutaneous injection to achieve sufficient biological effect. Although subcutaneous injections are simple to administer, it is generally accepted that only a low percentage of the injected drug reaches the target location. Further, the daily injections are bothersome and painful for patients. Thus, a more efficient, advanced application is desirable. We used CG hydrogels in this study as a source of sustained delivery of G-CSF. G-CSF release can be regulated by changing the levels of hydrogel degradability, which can be controlled by regulating the crosslinking extent of the hydrogel. CG hydrogels have been shown to degrade gradually and completely in mouse muscle by day 14. 19 It is expected that controlled release from CG hydrogel would enable not only an increase in the local and systemic G-CSF concentration, but also a prolongation of the period of enhanced concentration at the implanted site of the hydrogels. In this study, we could not detect any remnants of the hydrogel at 2 weeks on histological analysis. The results indicated that the effect of the hydrogels in this study did not continue up to 2 weeks. However, the examination to investigate the concentration of G-CSF in the systemic circulation using ELISA revealed that the concentration of G-CSF was significantly upregulated in the G-CSF group as well as the sG-CSF group, compared with the control group, up to 1 week postoperation. The result indicated that the release of the G-CSF continued at least for 1 week postoperation. Taking these data into consideration, we suggest that the slow release of the G-CSF from the hydrogels continued for about 1–2 weeks postoperation.
In our study, G-CSFR+ and CD34+ cell fractions were mobilized into PB after bone defect operations and the ratios were significantly upregulated by G-CSF treatment up to day 7 by flow cytometry analysis. Immunofluorescent staining with CD34 revealed that CD34+ cells were incorporated into bone defects after 2 weeks and the CD34+ cells were increased in the G-CSF group. Vasculogenesis by EPCs, which is involved in the development of the blood vessel system in the embryonic stage,45,46 was not identified as a mechanism of postnatal endothelial regeneration until the discovery of BM-derived and circulating EPCs in adults.47–49 Since then, many researchers have applied EPCs to therapeutic neovascularization and acquired beneficial results50–52 A recent report revealed that circulating human CD34-positive cells mobilized by G-CSF were demonstrated to have a therapeutic potential for promoting both neovascularization and osteogenesis in damaged skeletal tissue, which also allows complete healing. 17 In our results, we demonstrated the possibility of contributing to bone regeneration via mobilization of CD34+ cells both in PB and at the bone defect with locally applied G-CSF. Recently, the notion of “circulating OB lineage cells” following fractures has also been reported.53–55 In these reports, the concentration of circulating cells positive for OC increased after fracture, suggesting that they might play a functional role in the healing of the fracture, 54 and 37% of OC-positive cells also expressed CD34. 55 It is known that CD34-positive cells are also mobilized into circulation following an acute musculoskeletal traumatic event in humans, 56 and G-CSF-mobilized PB human CD34-positive cells could differentiate into OBs in an in vitro study. 18 Thus, it has been suggested that systemic reactions happen following local fracture events, including osteogenesis and hematopoiesis/vasculogenesis, and these reactions closely regulate each other in terms of microenvironmental interaction for regenerative activity in bone repair. 17 In this study, the systemically applied G-CSF (the sG-CSF group) also promoted bone regeneration. However, the results also revealed that the bone healing capability in the sG-CSF group was inferior to that of the locally applied G-CSF. We suggested that the effect of G-CSF did not totally depend on the systemic effect but on the direct local effect, and the locally applied G-CSF promoted a more favorable condition for bone regeneration in this model. In this study, CD34+ cells mobilized into bone defects existed mainly in the capillary, rather than the lining of the bone cavity. This result suggested that CD34+ cells at the bone defect area contributed mainly to neovascularization via angiogenesis/vasculogenesis, not to osteogenesis.
In the present study, whether G-CSF gathered circulating CD34/OC+ cells, so-called “circulating OB lineage cells,” into bone defects could not be confirmed. It also remained unknown as to the rate at which CD34+ cells were mobilized from PB. Although it is tempting to consider the main origins of CD34+ cells were neighboring soft tissues, we believe that some CD34+ cells may have been derived from circulating mobilized cells. For example, estimating vascular invasions at the bone defects, histochemical staining for isolectin B4 revealed that capillary density was remarkably increased at 2 weeks in the G-CSF, G-CSF + sFlt1, and sG-CSF groups. However, the increment in the G-CSF + sFlt1 group was significantly inferior to that of G-CSF and sG-CSF groups at 2 weeks, and the bone formation in the G-CSF + sFlt1 group was significantly reduced, compared with the G-CSF group at 8 weeks. Gene expression analysis also revealed that VEGF mRNA expressions were upregulated significantly at 1 and 2 weeks in the G-CSF group. These results indicated that neovascularization via angiogenesis, which was partially mediated by VEGF, and vasculogenesis were promoted by locally applied G-CSF and partially contributed to promoting bone formation; however, it was impossible to confirm whether the capillaries were living, which remained a limitation of this study.
In contrast, immunohistochemical staining with OC showed that OC+ cells were significantly increased at the bone defect in the G-CSF group and real-time RT-PCR analysis also revealed that OC mRNA expression was upregulated at 1 and 2 weeks postoperation in the G-CSF group, which was significant. Further, double immunohistochemistry with OC/G-CSFR confirmed that more OC/G-CSFR+ cells were incorporated into the bone defect site by G-CSF. These results indicated that G-CSF promoted bone regeneration via OB incorporation at the bone defect site.
Indeed, there have been controversial reports indicating that G-CSF inhibited osteogenesis. 57 Christopher et al. reported that a very high dose of daily 250 μg/kg/day injection of G-CSF induced OB apoptosis and inhibited OB differentiation in a mouse model. Their results appear to contradict our study; however, our study examined the effect of G-CSF, not under normal conditions but in a fracture environment, which is totally distinct from their study. It is possible that their results were also true, because our preliminary studies found that local application of 50 μg G-CSF in vivo did not induce bone regeneration, and the in vitro study examining the migration activity of MSCs by G-CSF found that the migration activity of G-CSF had not upregulated dose dependently (it reached its peak at 5 ng/mL; data not shown in this study). Generally, almost all growth factors require an appropriate dosage to stimulate bioactive effects and this is also true for G-CSF; it has been accepted that G-CSF at low doses (3.5–6 μg/kg/day) can be successfully administered with minimal toxicity in normal subjects, which consistently increases the number of circulating progenitor cells of multiple hematopoietic lineage. There were also some suggestions that high doses of G-CSF might cause sarcoma production.58,59 Therefore, we need to carefully calculate the appropriate dose of G-CSF for each condition and patient. Taking into consideration that only a low percentage of the injected drug reaches the target location, the dosage of 5 μg G-CSF applied locally in this study may be more favorable and appropriate for clinical use.
In conclusion, G-CSF appeared to have a bone formation-promoting capability in vivo. It is likely that locally applied G-CSF contributes to an ideal local environment for fracture healing by supplying adequate blood flow and stimulating osteogenesis. Our study showed one method for promoting a native biological reaction after fracture via angiogenesis/vasculogenesis regulated by CD34+ cell mobilization and via osteogenesis regulated by OB stimulation. Further investigation is needed to evaluate the biomechanical properties of this reparative bone. Although an examination to investigate a more detailed mechanism, such as how OB was stimulated by G-CSF, was not performed and these are limitations of this study, our study indicates that G-CSF may have the therapeutic potential for bone regeneration.
Footnotes
Acknowledgment
We would like to thank Ms. Janina Tubby for her editing assistance in preparing this manuscript.
Disclosure Statement
No competing financial interests exist.