
Abstract
Smooth muscle cells (SMCs) play an essential role in maintaining the structural and functional integrity of blood vessel and thus is a critical element for blood vessel construction via tissue engineering approach. Adipose-derived stem cells (ASCs) represent a reliable source of mesenchymal stem cells with multidifferentiation potential. In this study, the feasibility of differentiation of human ASCs (hASCs) into cells with phenotypic and functional properties of SMCs was explored. hASCs isolated from human lipoaspirate were expanded to passage 5 and then induced with administration of transforming growth factor-β1 (TGF-β1) and bone morphogenetic protein-4 (BMP4) either alone or in combination with culture medium. Expression of SMC-related markers including alpha-SM actin (α-SMA, SM22α, calponin, and SM myosin heavy chain) were detected by immunofluorescent staining, reverse transcription (RT)-polymerase chain reaction, and western blot analysis. It was found that only under the circumstance of a combined stimulation with TGF-β1 and BMP4, both early and mid markers (α-SMA, SM22α, calponin) as well as a late marker (SM myosin heavy chain) of SMC differentiation were identified to similar levels as those in human umbilical artery SMCs. More importantly, these SM differentiated cells showed the function of contracting collagen matrix lattice when they were entrapped inside. The contractile function of differentiated hASCs was further enhanced by direct exposure to 60 mM KCl, consistent with what occurred in human umbilical artery SMCs. These results provide evidence that ASCs possess the potential to differentiate into contractile SM-like cells when stimulated by TGF-β1 and BMP4 together. SMCs differentiated from hASCs may provide an abundant source as seed cells for blood vessel engineering.
Introduction
Mesenchymal stem cells (MSCs) have a self-renewal capacity, long-term viability, and a potential to differentiate into diverse cell types, such as adipogenic, osteogenic, chondrogenic, and myogenic lineages.3,4 It has been demonstrated that under the stimulation of transforming growth factor-β1 (TGF-β1), bone marrow-derived MSCs (BMSCs) could differentiate along SMC pathway characterized by the expression of specific contractile proteins including alpha-SM actin (α-SMA) and calponin as well as SM myosin heavy chain (SM-MHC).5,6 Further, a small-diameter vascular graft with an intact SM layer engineered from BMSCs displayed long-term potency after implantation in a canine model. 7 More importantly, it was observed that SMCs differentiated from BMSCs synthesized higher amount of elastin after implantation for 6 months when compared with vascular-derived SMCs. 8 However, the characteristics of BMSCs, such as low frequency of existence, complications in harvesting, and time consuming in expansion, urged us to find a superior one to BMSCs for such purpose. Meanwhile, it is notable that proliferative and multipotent capacity of BMSCs decreased dramatically in donors with age as the aging population most likely suffers from cardiovascular or peripheral blood vessel diseases.9,10 Adipose tissue represents a promising reservoir of MSCs with multidifferentiation potential into adipogenic, osteogenic, chondrogenic, and myogenic lineages under appropriate conditions. 11 Adipose-derived stem cells (ASCs) are easier to acquire, have relatively lower donor site morbidity and a higher yield at harvest, and can expand more rapidly in vitro when compared with BMSCs. 12 In addition, multipotency of ASCs has been shown to be independent of age of donors.13,14 Thus, ASCs could be a preferred novel cell source for blood vessel engineering.
The critical role of TGF-β1 in vasculature formation and SMC specification during development was demonstrated in TGF-β1−/− mice which died at E10.5–E11.5 with defective hematopoiesis and vasculogenesis due to the decrease in endothelial–mesenchymal cell contact. 15 TGF-β1 was also shown to induce in vitro differentiation of embryonic 10T1/2 cells 16 and Monc-1 neural crest stem cell line 17 into SMCs, respectively. In combination with platelet-derived growth factor BB. The platelet-derived growth factor (PDGF) family of disulfide-linked dimeric proteins consists of four homodimeric proteins, PDGF-AA, PDGF-BB, PDGF-CC and PDGF-DD, and one heterodimeric protein, PDGF-AB 18 or ascorbic acid, 19 BMSCs could be induced by TGF-β1 to differentiate into cells with SMC-like phenotype and function. Besides TGF-β1, bone morphogenetic protein-4 (BMP4) pathway was reported to act as a major regulator in vasculogenesis during embryonic development. 20 According to Boyd et al., 21 formation of primitive vascular network in human embryonic stem cell-derived embryoid was remarkably promoted by stimulation with BMP4. In addition, the expression of SMA and maintenance of the differentiated SM phenotype was enhanced in human pulmonary artery SMCs by treatment with BMP4. 22 Further, stimulation of procaspase-activating compound 1 (PAC-1) cells with TGF-β1 and BMP4 together induced a synergistic activation of SMA, indicating a potentially distinct mechanism of inducing SM genes by TGF-β1 and BMP4 combinational signaling pathway. 22
Inspired by the above findings, this study was aimed to investigate the potential of human ASCs to differentiate into SMC phenotype upon induction by TGF-β1 and BMP4 in low-serum medium. Expression of characteristic SMC markers in the resulting ASCs was examined at both proteinic and genetic levels. Moreover, the contractile function of induced ASCs featured by the ability of cells to contract collagen gel was further demonstrated.
Materials and Methods
Isolation and culture of human ASCs
Fresh human lipoaspirates were obtained from five healthy female patients (at an average age of 30 years) who had received abdominal liposuction in the Department of Plastic and Reconstructive Surgery of Shanghai 9th People's Hospital. All protocols of human tissue handling were approved by the Research Ethical Committee of the Hospital. Processed lipoaspirate cell isolation and culture were performed as previously described.11,23 Briefly, fresh lipoaspirates were washed intensively with phosphate-buffered saline (PBS) and then treated with 0.075% type I collagenase (Sigma-Aldrich, St. Louis, MO) under shaking at 37°C for 60 min. The enzyme activity was neutralized with low-glucose Dulbecco's modified Eagle's medium (LG-DMEM; Gibco, Grand Island, NY) containing 10% fetal bovine serum (FBS; HyClone, Logan, UT). The digested lipoaspirates were then centrifuged at 1200 g for 10 min to obtain a high-density stromal vascular fraction which was further filtered through a 50-μm nylon mesh to remove undigested tissue and then centrifuged again at 600 g for 10 min. The supernatant was discarded and the pellet was resuspended in LG-DMEM (Gibco) supplemented with 10% FBS, 100 U/mL penicillin (Sigma-Aldrich), and 100 mg/mL streptomycin (Sigma-Aldrich) (defined as growth medium). The cells were then plated in Φ 100-mm culture dishes (Falcon, Oxnard, CA) at a density of 4 × 104 cells/cm2, with the medium changed twice a week. When reached 70–80% confluence, cells were passaged and human ASCs (hASCs) of passage 5 were used in the following study. The characterization of hASCs was determined by their CD marker profile and their ability to differentiate into osteogenic, adipogenic, and chondrogenic lineages as previously reported (data not shown). 11
For cell proliferation assay, hASCs were plated at 3 × 103 cells/cm2 in Φ 35-mm dishes. Cell numbers in different media were determined by DNA assay as previously reported. 24 Briefly, cells at indicated time points were crushed for full lysis with 0.5 mL proteinase K (0.5 mg/mL; Sigma-Aldrich) at 56°C overnight. The resulting mixture was subjected to centrifugation and aliquots (40 μL) of the supernatants after mixing with 160 μL Hoechst 33258 dye solution (0.1 g/mL; Sigma-Aldrich) were transferred to black flat-bottomed 96-well plates (Corning Costar, Cambridge, MA). DNA content was quantified spectrofluorometrically using a Varioskan multimode detection reader (Thermo Electron, Waltham, MA) at a wavelength of 465 nm (the emission wavelength of 360 nm) by correlating with a DNA standard curve which was generated by lysing serial dilutions of a known concentration of hASCs.
Induction of SM differentiation
To evaluate the effects of TGF-β1 and BMP4 on the differentiation of hASCs into SMCs, hASCs reaching subconfluence were cultured in different media and assigned into five respective groups as follows: (1) DMEM supplemented with 10% FBS (growth medium); (2) DMEM supplemented with 1% FBS, defined as the basal medium (BM); (3) DMEM supplemented with 5 ng/mL recombinant human TGF-β1 (R&D Systems, Minneapolis, MN) and 1% FBS; (4) DMEM supplemented with 2.5 ng/mL recombinant human BMP4 (R&D Systems) and 1% FBS; (5) DMEM supplemented with 5 ng/mL TGF-β1, 2.5 ng/mL BMP4, and 1% FBS. Human umbilical artery SMCs (hUASMCs) isolated as described previously were set as a positive control. The culture media were changed every 2 days. Cell characterization and functional evaluation were performed after 7 days of culture.
Immunofluorescent staining
Immunofluorescence was performed on methanol-fixed cells using the following primary antibodies: mouse monoclonal anti-α-SMA (C6198; Sigma-Aldrich), rabbit polyclonal anti-SM22α (ab14106; Abcam, Cambridge, United Kingdom), rabbit monoclonal anti-calponin (ab46794; Abcam, Cambridge, United Kingdom), and mouse monoclonal anti-SM-MHC (M7786; Sigma-Aldrich) antibodies. After reaction with primary antibodies for 60 min at room temperature, the cultures were washed with PBS for three times. Fluorescein isothiocyanate (FITC)-conjugated goat anti-rabbit secondary antibody (Millipore, Billerica, MA) was used to detect the localization of anti-SM22α and anti-calponin antibodies, respectively. FITC-conjugated goat anti-mouse secondary antibody (Millipore) was used to detect the localization of anti-α-SMA and anti-SM-MHC antibodies. Cell nuclei were stained with propidium iodide. Control samples consisted of cells without primary antibody and were used to assess background fluorescence. The images were viewed by a fluorescence microscope (Nikon, Tokyo, Japan).
RNA isolation and reverse transcription-polymerase chain reaction
For the reverse transcription (RT)-polymerase chain reaction (PCR) detection of the SMC-specific markers (α-SMA, SM22α, calponin, SM-MHC), total cellular RNA was extracted from cells cultured under different conditions for 7 days using Trizol Reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's protocol and complementary DNA (cDNA) was synthesized using PrimeScript 1st Strand cDNA synthesis kit (TaKaRa Biotechnology, Dalian, P.R.China). The reactions were performed and monitored in a T3 thermocycler (Biometra, Goettingen, Germany) for 35 cycles after the initial 3-min denaturation at 95°C. The same reaction profile was used for all primers including human α-SMA, SM22α, calponin, and SM-MHC. The primer sequences designed using the Primer Express software are listed in Table 1. The reactions were performed with 35 cycles as follows: 95°C for 30 s, 56°C for 30 s, and 72°C for 60 s. The amplified products were analyzed by 1.5% agarose gel electrophoresis and visualized by ethidium bromide staining.
α-SMA, alpha-smooth muscle actin; SM-MHC, smooth muscle myosin heavy chain; ALP, alkaline phosphatase; OCN, osteocalcin; Cbfa1, core binding factor alpha 1.
Quantitative real-time PCR
Total RNA was extracted from cells using an RNAprep Micro Kit (TianGen Biotech, Beijing, China) according to the manufacturer's instruction. The RNA concentration of the extract was determined from the optical absorbance at 260 nm. cDNA was synthesized using PrimeScript 1st Strand cDNA synthesis kit (TaKaRa). The reactions were performed and monitored in a T3 thermocycler (Biometra). Real-time PCR was performed using quantitative real-time amplification system (MxPro-Mx3000P; Stratagene, La Jolla, CA). SybrGreen PCR MasterMix (Applied Biosystems, Foster City, CA) was used in each reaction. Relative expression levels for each gene of interest were calculated by normalizing the quantified cDNA transcript level (cycle threshold) to that of the β-actin. For detecting the possibility of osteogenic differentiation of hASCs treated with the above different media, gene expression of alkaline phosphatase (ALP), osteocalcin (OCN), and core binding factor alpha 1 (Cbfa1) was evaluated by real-time PCR. hASCs cultured in osteo-induced medium (LG-DMEM, 10% FBS, 0.01 μM 1,25-dihydroxyvitamin D3, 50 μM ascorabate-2-phosphate, 10 mM β-glycerophosphate) for 14 days were set as a positive control in this experiment.
Flow cytometry analysis
For flow cytometry analysis, cells were harvested, fixed for 30 min in ice-cold 2% paraformaldehyde, and washed in flow cytometry buffer (FCB: 1 × PBS, 2% FBS, 0.2% Tween-20). Single-cell suspensions of 106 cells/mL were incubated with primary antibodies: α-SMA (Sigma-Aldrich; 1:100), SM22α (Abcam; 1:250), calponin (Abcam; 1:100), or SM-MHC (Sigma-Aldrich; 1:50) in PBS for 60 min on ice, then washed three times with PBS, and incubated for 30 min on ice with FITC-conjugated anti-mouse secondary antibody (α-SMA, 1:100; SM-MHC, 1:50) or FITC-conjugated anti-rabbit secondary antibody (SM22α, 1:250; calponin, 1:100) (Millipore). After three further washes, flow cytometry was performed on a FACS Caliber flow cytometer (Becton Dickson, San Jose, CA). In each immunofluorescence experiment, an isotype-matched IgG control was also used.
Western blot analysis
Western blotting was performed to identify vascular SMC-specific cytoskeletal proteins. Briefly, when it came to the predetermined time points of each group, cultures were washed three times with ice-cold PBS and total cell lysate was prepared by extracting proteins with a lysis buffer (50 mM Tris-HCl, pH 7.4, containing 150 mM NaCl, 1% Nonidet P-40, 0.1% sodium dodecyl sulfate, and 0.1% deoxycholate). Proteins were separated by 8% polyacrylamide gel electrophoresis containing 0.1% sodium dodecyl sulfate and subsequently transferred to nitrocellulose membrane. The nitrocellulose sheet was blocked with 2% nonfat dry milk and 3% BSA in Tris-buffered saline. The monoclonal antibodies, rabbit anti-SM22α, anti-calponin, and mouse anti-α-SMA, anti-SM-MHC, anti-β-actin were applied to detect SMC-specific markers. The blots were developed using an IRDye 700DX-and IRDye 800CW-conjugated secondary antibody (Rockland Immunochemical, Gilbertsville, PA), and proteins were visualized by the Odyssey system (LI-COR Biosciences, Lincoln, NE). The western-blotting results were quantified using Gel-Pro Analyzer (Version 4.5) software and the results were expressed as the ratio of α-SMA, SM22α, calponin, or SM-MHC/β-actin.
Collagen gel lattice contraction assay
The contractility of cells was measured as previously described. 25 Briefly, cells were trypsinized with trypsin–ethylenediaminetetraacetic acid and resuspended in serum-free DMEM at a density of 1 × 106 cells/mL. To prepare collagen gels, a mixture of 800 μL collagen solution (rat tail collagen type I powder [Sigma-Aldrich] dissolved in acetic acid, 4.0 mg/mL) and 100 μL of 10 × DMEM was neutralized to pH 7.0 with sodium bicarbonate. Cells (5 × 105) suspended in 100 μL fresh FBS were added into the above mixture. Then the resulting cell/collagen mixture was transferred to 24-well culture plates for 1 h to polymerize the collagen cell lattices. The lattices were then transferred to six-well culture plates by gently pipetting medium at the lattice–dish interface from the bottom of the 24-well culture plates and initiated collagen gel contraction. Serum-free medium was added to facilitate cell growth, and gels were incubated for an additional 48 h. The extent of gel contraction of each group at different time points was calculated by measuring the dimensions of the gel lattices and viewed by a digital camera (T70; Sony, Tokyo, Japan). The area of gel lattices was analyzed using NIH ImageJ software. Quantitative results were interpreted by measuring the relative lattice area which was obtained by dividing the area at each time point by the initial area of the lattice.
Statistics
Each experiment was performed with three of five different donors and in each group we repeated the experiment for at least three times. The data were expressed as mean ± standard deviation. Independent-sample T tests (Student's t-test) assuming equal variance were performed using SPSS 11.0 software for statistical analysis. A p-value of less than 0.05 was considered statistically significant.
Results
Effects of various concentrations of TGF-β1 and BMP4
As shown in Figure 1A, first, the effects of different doses of TGF-β1 on SMC differentiation was evaluated, and it could be seen that TGF-β1 at a dose of 5 ng/mL had a higher induced level than that at 1 and 2.5 ng/mL, respectively. With the further increase to 7.5 and 10 ng/mL, respectively, expression of SM22α and calponin were raised slightly, but the expression of SM-MHC was lowered. Thus the dose of TGF-β1 was fixed to 5 ng/mL in the following studies. Second, we compared different doses of BMP4 (1, 2.5, 5, and 10 ng/mL, respectively) combined with 5 ng/mL TGF-β1 to see their synergetic effects on differentiation of hASCs into SMC lineage (Fig. 1B). It was shown that 2.5 ng/mL BMP4 plus 5 ng/mL TGF-β1 group had the highest expression levels of α-SMA and calponin. With the increase to 5 and 10 ng/mL, respectively, no significant enhancement in the transcript levels of SM22α and SM-MHC could be observed. Hence, from the viewpoint of transcript expression of these SMC-specific markers, we chose TGF-β1 at 5 ng/mL plus BMP4 at 2.5 ng/mL to induce the differentiation of hASCs into SMC lineage in this study.
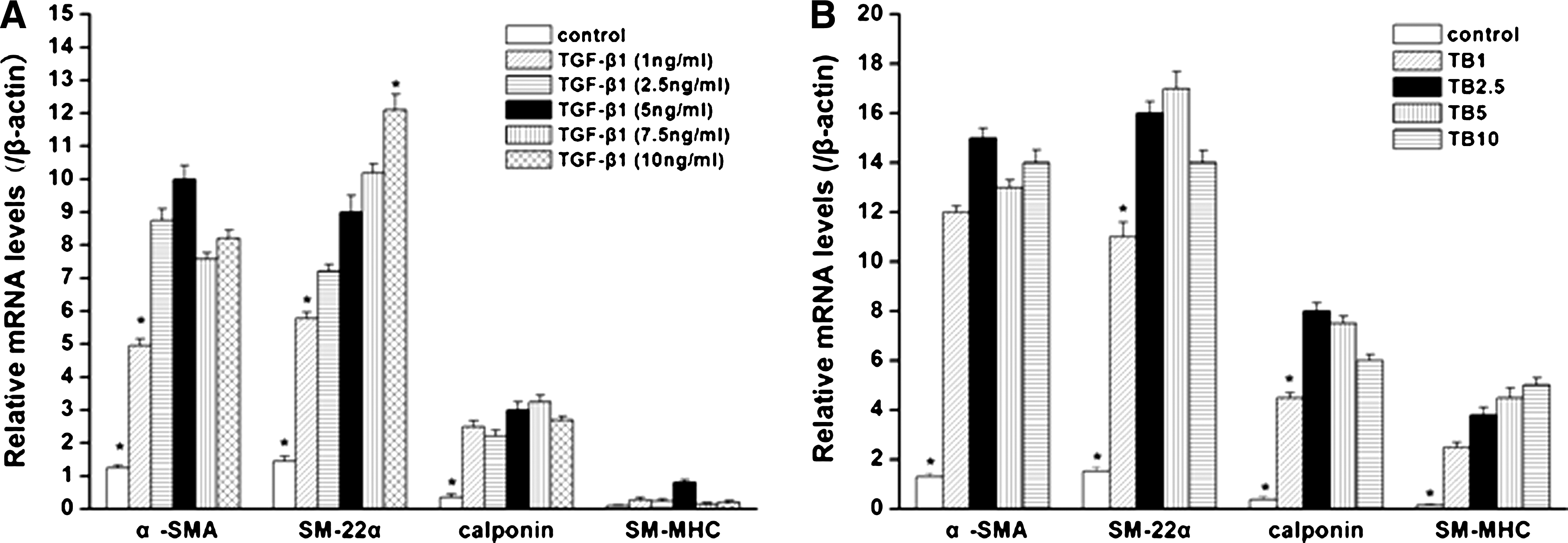
SMC-specific transcript expression (α-SMA, SM22α, calponin, SM-MHC) in hASCs with various concentrations of TGF-β1 and BMP4. (
Acquisition of SMC morphology by hASCs induced with TGF-β1 and BMP4
At the fifth passage, hASCs appeared as a relatively homogenous population that exhibited a fibroblast-like spindle morphology during culture (Fig. 2A). These cells when cultured in BM exhibited a flat morphology (Fig. 2B). When the cells were treated either with TGF-β1 (Fig. 2C) or BMP4 (Fig. 2D) as well as the combination of them (Fig. 2E) for 7 days, hASCs acquired a spindle morphology and grew in a “hill and valley” pattern similar to what was observed in primary isolated hUASMCs (Fig. 2F). And as shown in the inset images by phalloidin staining, we could demonstrate that the stress–fiber pattern of hASCs induced with TGF-β1 or BMP4 as well as TGF-β1 plus BMP4 was similar to the one of primary SMCs isolated from human umbilical artery. On the basis of morphological acquisition, it seems that optimal SM differentiation was observed in cells treated with the combination of TGF-β1 and BMP4 compared with cells treated with each of them.
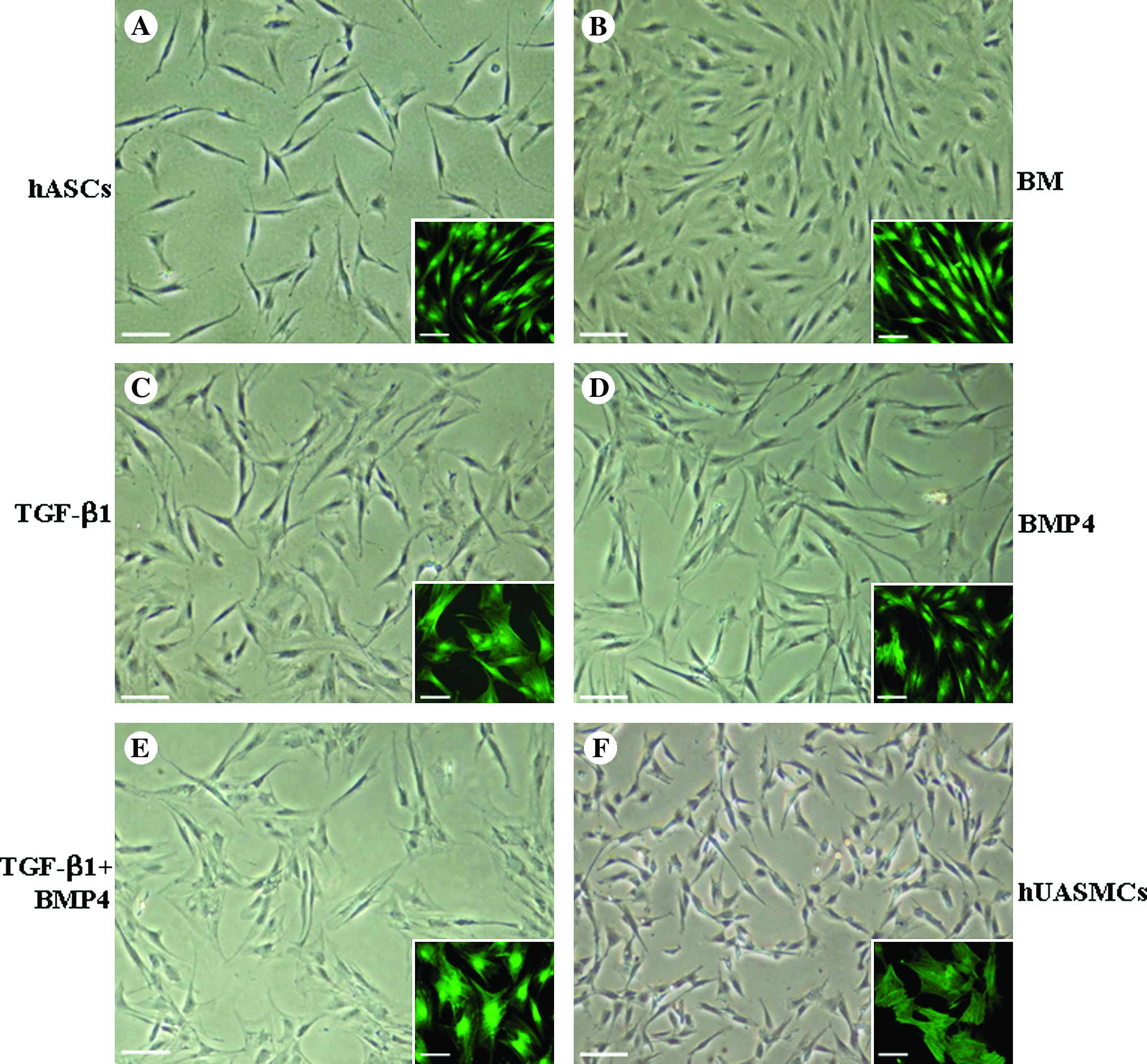
Respective and combinational effects of TGF-β1 and BMP4 on the morphology of hASCs. Phase-contrast images of hASCs of the fifth passage cultured in normal medium (
Expression of SMC-specific markers in hASCs treated with TGF-β1 and BMP4
To determine whether hASCs were able to acquire an SMC phenotype when treated with TGF-β1 and BMP4, SM-specific contractile proteins (α-SMA, SM22α, calponin, and SM-MHC) were detected by immunofluorescent staining. In parallel, expression of these markers in hUASMCs was also examined to serve as a positive control. As shown in Figure 3, there was a baseline expression of α-SMA and SM22α in undifferentiated hASCs (growth medium) and hASCs cultured in BM (1% FBS), respectively. However, expression of calponin could be detected merely in a small fraction of cells cultured in BM and no expression of SM-MHC could be observed in cells of undifferentiated and BM groups. When being exposed to TGF-β1 (5 ng/mL) or BMP4 (2.5 ng/mL) alone, expression of α-SMA and SM22α were found to be enhanced. The expression of calponin was enhanced by the treatment of TGF-β1 but not so obviously by BMP4. Further, expression of SM-MHC which represents a late marker of SMC differentiation was weakly detected in hASCs stimulated with TGF-β1, whereas such an expression was remarkably enhanced in hASCs stimulated with the combination of TGF-β1 and BMP4 for 7 days, reaching a similar level as that of hUASMCs.
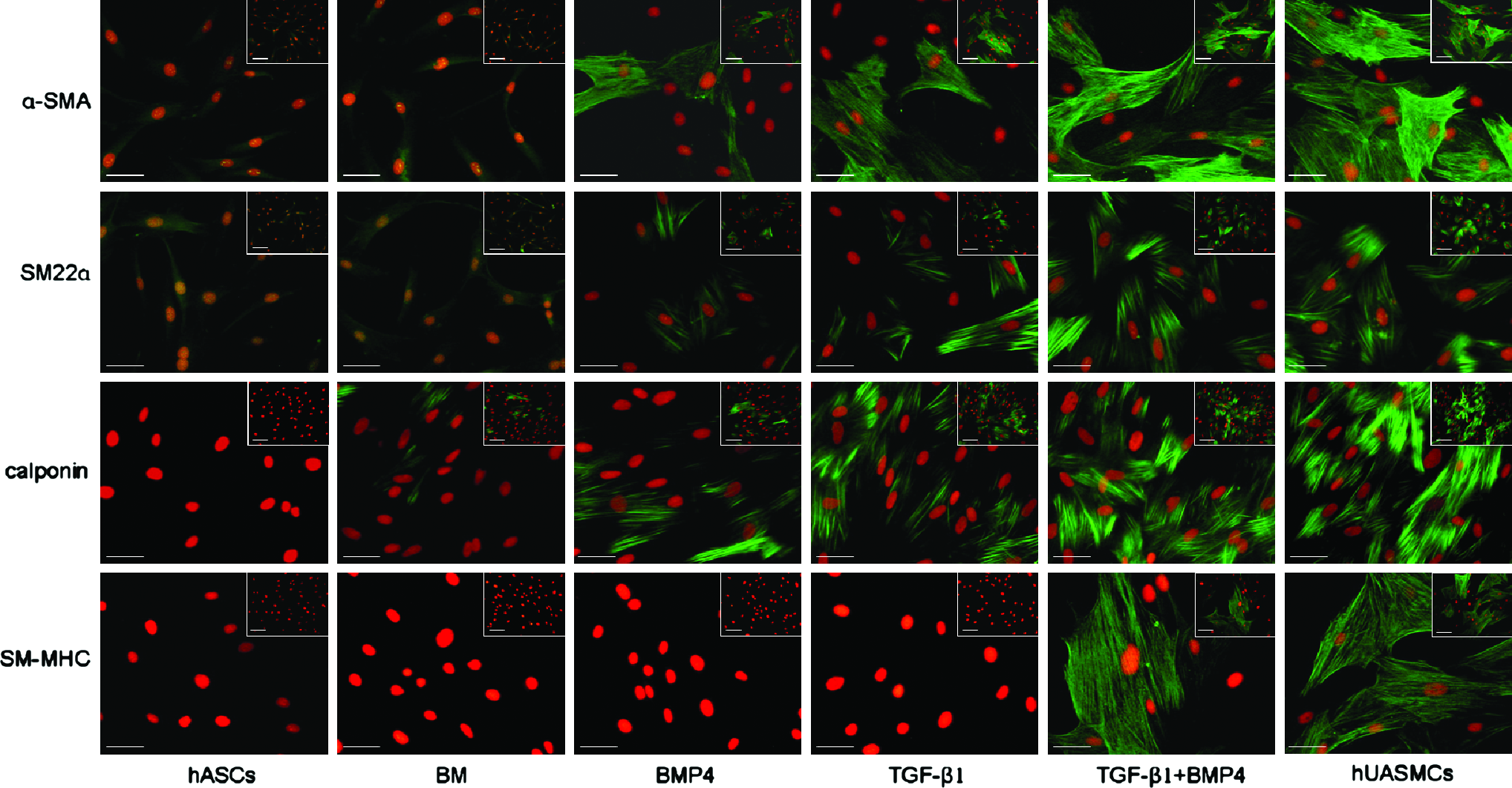
Expression of SMC-specific proteins (α-SMA, SM22α, calponin, SM-MHC) in hASCs under different culture conditions for 7 days. Cells were subjected to immunofluorescence staining with antibodies against α-SMA, SM22α, calponin, and SM-MHC (green) and nuclear staining with propidium iodide (red), respectively. Representative example of more than five experiments. Scale bars: 25 μm for all figures; 50 μm for insets. Color images available online at www.liebertonline.com/ten.
To ascertain the results of immunofluorescent staining, expression of α-SMA, calponin, SM22α, and SM-MHC in the protein level was analyzed by western blot detection (Fig. 4A). In consistence with the immunostaining results, expression of α-SMA was detectable in either undifferentiated or growth factor-treated hASCs. A faint baseline expression of SM22α could be detected in undifferentiated hASCs. Upon stimulation by TGF-β1 or BMP4 alone, the expression of calponin and SM22α was found to be enhanced. More importantly, it was found that only under the circumstance of a combined stimulation with TGF-β1 and BMP4, both early (α-SMA, calponin, and SM22α) and late markers (SM-MHC) of SMC differentiation were detected at a similar level to those in hUASMCs (Fig. 4A and C).
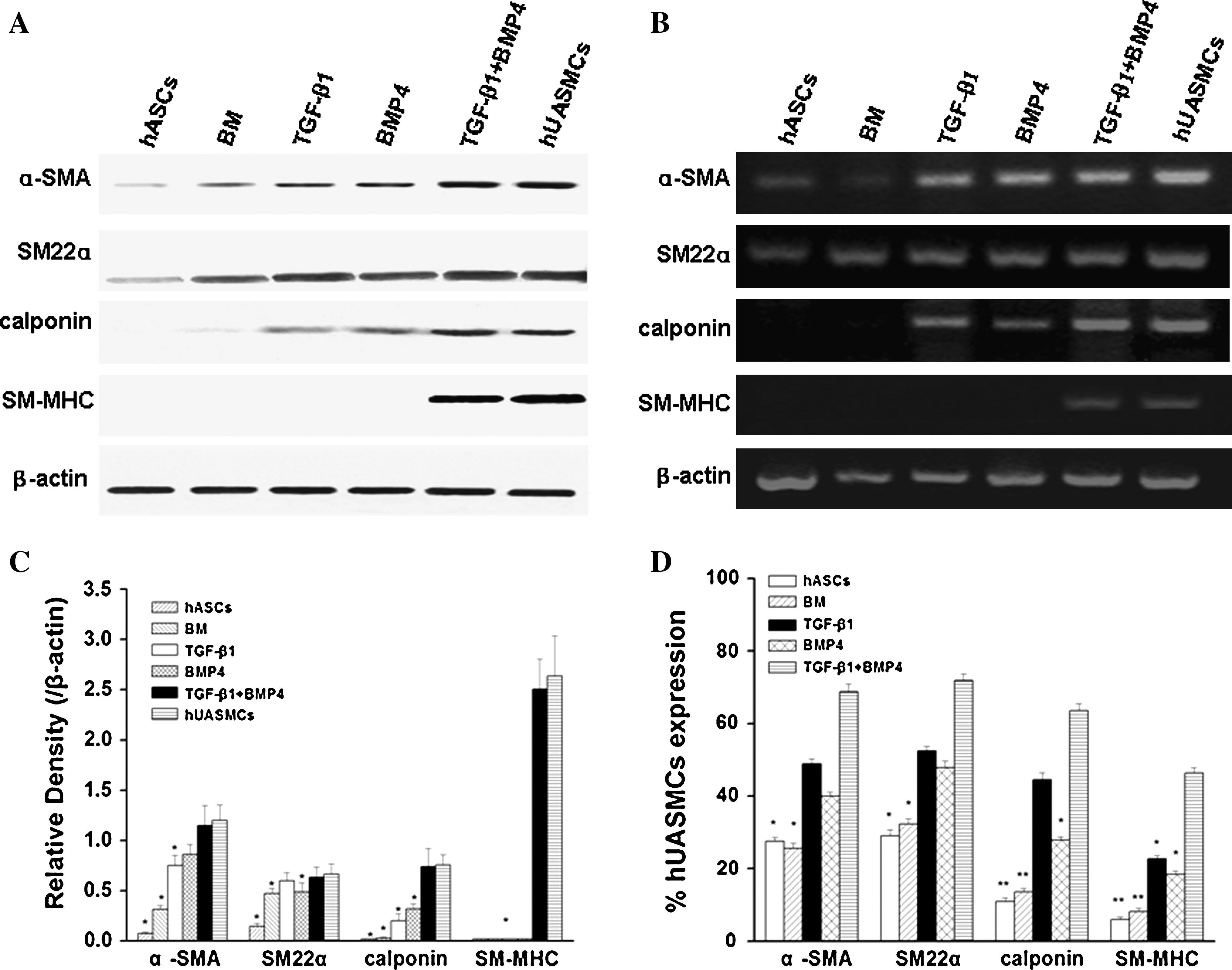
Expression of SMC-specific proteins (α-SMA, SM22α, calponin, SM-MHC) in hASCs at the transcript and protein levels. (
To further substantiate the effect of TGF-β1 and BMP4 on SMC differentiation of hASCs, both RT-PCR and real-time PCR were carried out to examine the transcript levels of the aforementioned SMC-related genes. As shown in Figure 4B, undifferentiated hASCs expressed only low levels of α-SMA and SM22α mRNA. The addition of TGF-β1 or BMP4 alone in the culture medium for 7 days resulted in the expression of calponin in hASCs, respectively. However, only upon the combined stimulation by TGF-β1 and BMP4 for 7 days, expression of SM-MHC was detected at a similar level to that expressed in hUASMCs. As further analyzed by real-time PCR quantitatively, transcript levels of α-SMA, calponin, SM22α, and SM-MHC in TGF-β1 and BMP4-treated group reached 66.3% ± 2.16%, 70.8% ± 3.54%, 61.4% ± 1.92%, and 45.6% ± 1.23% of that in hUASMCs, respectively (Fig. 4D). In addition, when compared with that in undifferentiated hASCs, the expression of α-SMA, calponin, SM22α, and SM-MHC of the same group was increased to 2.6-, 2.4-, 5.8-, and 7.8-fold, respectively. Taken the above results together, the inducing effects of TGF-β1 and BMP4 on SMC differentiation of hASCs in both protein and transcript levels can be confirmed.
As previous studies showed that SMCs express the contractile differentiated phenotype when the cells are not in a proliferated state, 26 we detected cell proliferation of hASCs cultured in different media at indicated time points using Hoechst 33258 dye DNA assay, and the results indicated that hASCs cultured in the growth medium kept on increasing in the duration of 10 days, whereas growth factor-treated hASCs, either by TGF-β1/BMP4 alone or in combination, exhibited a growth arrest as that cultured in BM (Fig. 5A). Further, as reported previously, BMP4 could initiate osteogenic differentiation of ASCs. 27 we therefore examined the expression of osteogenic related genes including ALP, OCN as well as Cbfa-1 in hASCs which were subjected to the above culture conditions by real-time PCR. Compared with hASCs cultured in osteogenic medium, no obvious expression of these osteogenic relevant genes could be detected in hASC-cultured media either containing BMP4 or not (Fig. 5B).
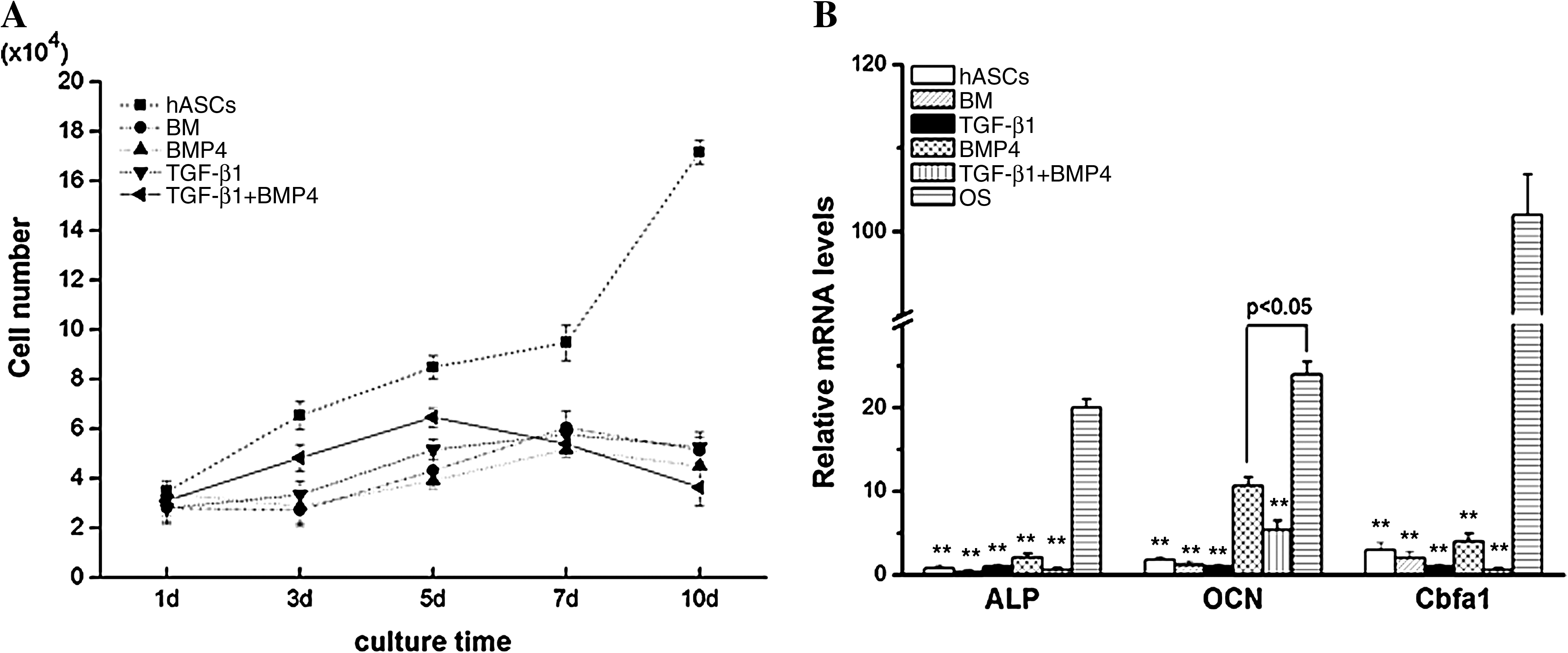
Proliferation and expression of osteogenic differentiation-related genes (alkaline phosphatase [ALP], osteocalcin [OCN], and core binding factor alpha 1 [Cbfa1]) of hASCs cultivated under different culture conditions. (
To quantify the percentage of SM differentiated cells in hASCs treated under different conditions, proportions of positive cells expressing α-SMA, SM22α, calponin, and SM-MHC, respectively, were determined by fluorescence-activated cell sorting. As shown in Figure 6, the α-SMA was detected in 21.6% ± 0.86%, 43.7% ± 1.35%, 36.7% ± 2.06%, and 64.4% ± 1.25% of undifferentiated hASCs, hASCs treated with TGF-β1 alone, hASCs treated with BMP4 alone, and hASCs treated with the combination of TGF-β1 and BMP4, respectively. Similar enhancement in SM22α-positive cells in the TGF-β1 and BMP4-treated group was observed. In comparison with that of undifferentiated hASCs (8.78% ± 2.02%), expression of calponin was significantly increased in the TGF-β1 (40.1% ± 1.86%) and BMP4 (23.6% ± 1.59%) treated alone as well as treated in combination groups (57.2% ± 2.08%). And most importantly, SM-MHC-positive cells reached about 50% of that of hUASMCs.
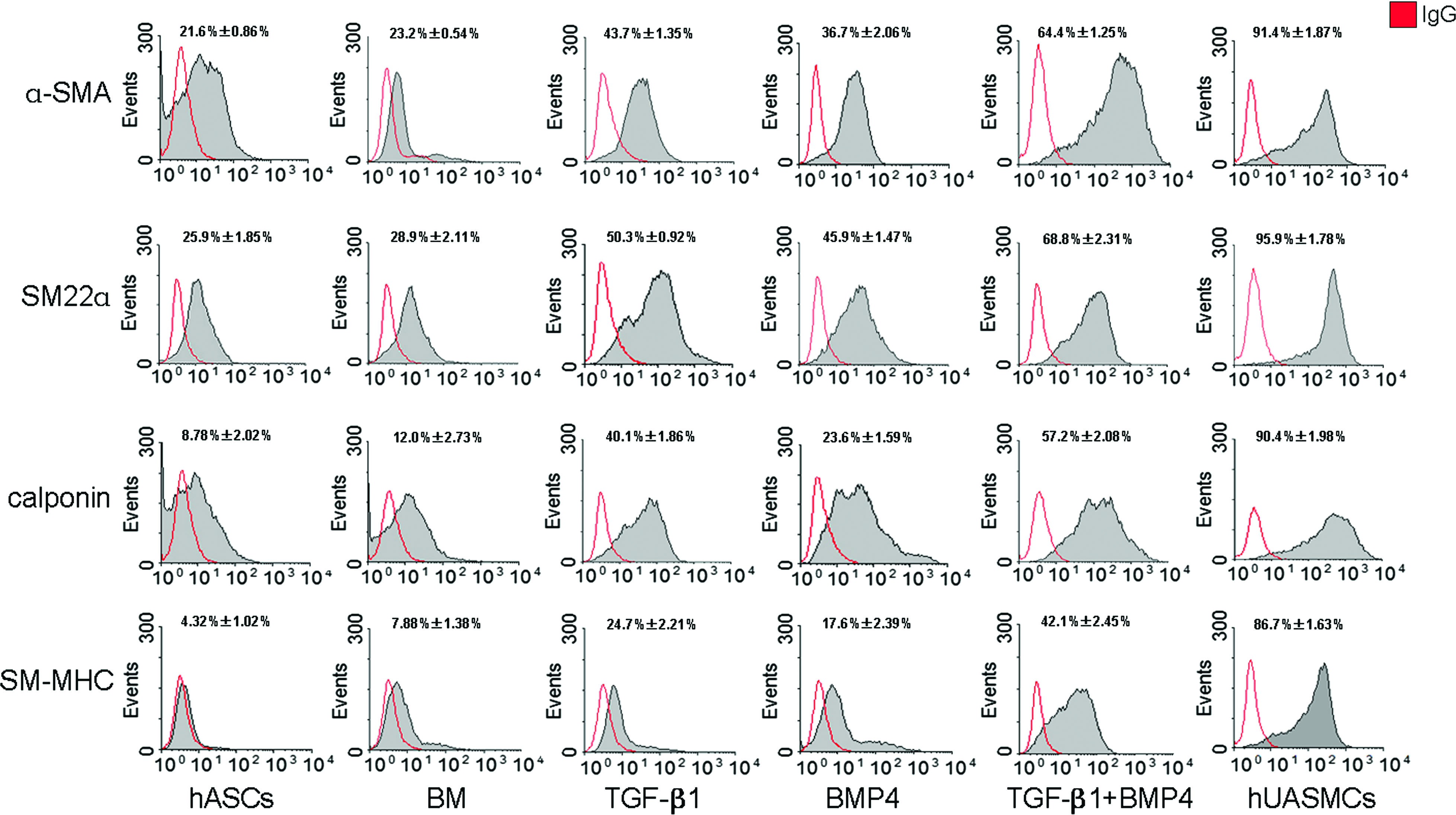
Fluorescence-activated cell sorting analysis of the expression of SMC-specific proteins (α-SMA, SM22α, calponin, SM-MHC) in hASCs cultured under different conditions for 7 days (n = 3). The percentages of positive cells expressing these specific markers increased significantly under the combinational stimulation with TGF-β1 and BMP4 to a similar high level as those corresponding ones in primary hUASMCs. Color images available online at www.liebertonline.com/ten.
Mediation of collagen gel matrix contraction by the SMCs differentiated from hASCs
The primary physiological function of SMCs is to contract and relax in response to various stimuli. Thus, to evaluate whether SMCs differentiated from hASCs that were induced by TGF-β1 and BMP4 together have the ability of contracting and relaxing, we cultured the cells in a collagen gel lattice and observed the size of gels with time. As shown in Figure 7A, the shrinkage of gels embedded with differentiated SMCs was remarkable and displayed a time-dependent manner. After 48 h, the area of gel lattice was decreased by almost 70% (Fig. 7B). However, the same type of collagen gel matrix containing undifferentiated hASCs kept quite stable in size in the duration of examination (around 90% at 48 h) (Fig. 7B). Compared with the gel containing undifferentiated cells, the area of collagen lattice with differentiated hASCs was significantly diminished since 6 h. Thus, these results indicated that with the combined induction by TGF-β1 and BMP4, hASCs differentiated into an SMC phenotype and even acquired the contractile ability as that of hUASMCs.
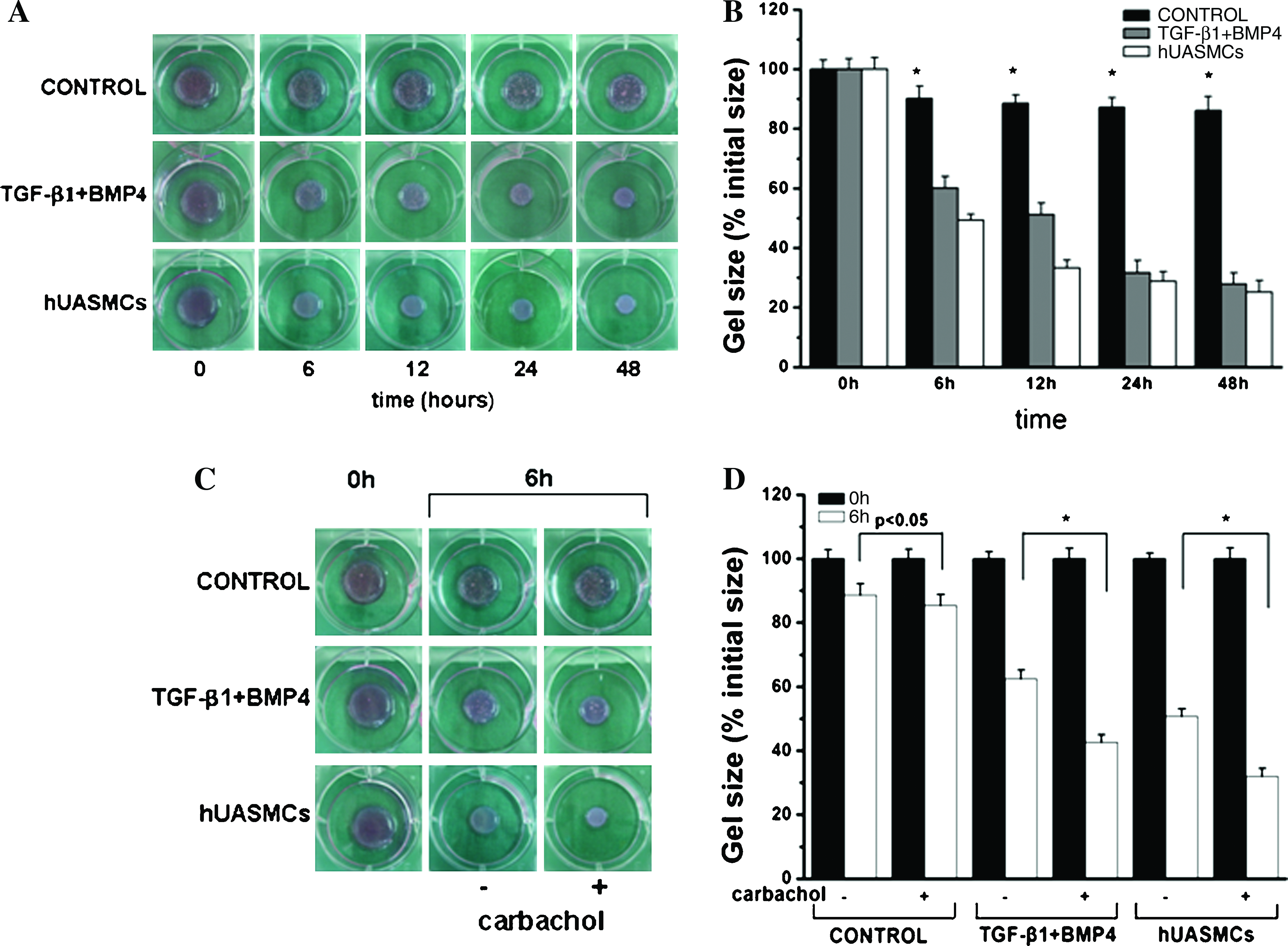
Contractile activity of the smooth muscle-like cells differentiated from hASCs. (
Then, we performed the contractile response of differentiated hASCs by using carbachol, a muscarinic agonist that can induce contraction of cultured SM. 28 As shown in Figure 7C and D, the size of collagen matrix with differentiated hASCs reduced to around 40% when cultured in the presence of carbachol at 6 h, whereas that in the absence of carbachol decreased to around 60%, which is consistent with the results of hUASMC group. In conclusion, the above results confirmed that differentiated hASCs showed contraction in response to the muscarinic contractile agonist carbachol, suggesting that hASCs can be differentiated into phenotypic and functional SMCs by TGF-β1 and BMP4 induction.
Discussion
Despite accumulating evidences showing that development of vascular grafts by tissue engineering approach held great potential in treating vascular diseases, cell source remains a major obstacle in translating the progress of studies to clinical application. The limited replicative capacity of adult mature SMCs made it not an optimal candidate as source of seed cells for constructing tissue-engineered blood vessels as documented in many early reports.29–31 Thereafter, many studies endeavored to explore whether functional SMCs could be generated from various types of adult MSCs. BMSCs were dominantly studied and possess a strong ability to differentiate into SMC lineage in response to a variety of stimuli such as growth factors,5,6 mechanical stress, 32 as well as coculture with endothelial cells. 33 As a major counterpart of MSCs derived from bone marrow, in vitro expanded hASCs has been shown to be capable of forming a microvessel-like structure in vitro which later improved postnatal neovascularization in a mouse model of hindlimb ischemia by incorporating into neo-generated vasculatures. 34 These results highlight the concept that hASCs represent an easily accessible, novel cell source for therapeutic angiogenesis and development of engineered blood vessels.
In this study, we demonstrated that hASCs acquired SMC phenotype as evidenced by their expression of specific structural proteins upon synergistic induction with TGF-β1 and BMP4 in vitro. It was also observed that there existed a baseline expression of α-SMA in the expanded cultures of hASCs. As an early marker of SMC differentiation, α-SMA is often adopted to characterize the preliminary SMC phenotype. 1 However, expression of α-SMA can often be observed in myofibroblasts and endothelial cells under certain conditions. 1 Hence, its expression alone does not provide definitive evidence for an SM lineage. Similar to the expression of α-SMA, it was found that there was a baseline expression of SM22α in undifferentiated hASCs and the expression level was almost unaffected by stimulation with either TGF-β1 or BMP4 alone. The above phenomena had also been observed by Rodríguez et al., 35 as they detected baseline expression of α-SMA and SM22α in clonogenic hASCs derived from primary cultures of processed lipoaspirate cells. Calponin and SM-MHC have been widely accepted as mid and late markers of SMC differentiation with more SMC correlativity, and in particular, SM-MHC is believed to be only expressed in contractile SMCs but not in any other cell types. 26 In this study, we found the initiation of calponin expression in hASCs in the presence of TGF-β1 or BMP4 alone as well as in the presence of both of them for 7 days, respectively. However, SM-MHC could only be detected when hASCs was stimulated by TGF-β1 and BMP4 together. Thus it was demonstrated that TGF-β1 and BMP4 together can induce the expression of all these four contractile proteins in hASCs. Further, we also tried to extend the induction time to 14 days and later, and we found that the expression of α-SMA, SM22α, calponin, and SM-MHC could still be detected in differentiated hASCs by immunofluorescent staining (data not shown). Moreover, to identify that the differentiated hASCs were not myofibroblasts, we performed immunofluorescence staining to detect the expression of vimentin and desmin. It was shown that no expression of vimentin was detected in the differentiated hASCs, whereas the expression of desmin was detected in the differentiated hASCs. (data not shown). All these results suggest that either TGF-β1 or BMP4 alone could initiate partial SMC differentiation of hASCs, whereas the combination effect of TGF-β1 and BMP4 synergistically stimulated full differentiation of hASCs toward SMC phenotype.
The role of TGF-β1 in regulating the proliferation and differentiation of SMCs and thereby the maturation of blood vessels in vasculogenesis is well documented. It has been reported that TGF-β1 promotes vessel maturation by stimulating extracellular matrix production and by inducing differentiation of mesenchymal cells to mural cells in mammalian development. 36 Studies of knockout mice also underscored the importance of TGF-β1, its receptor, and signaling components for proper SM development in the initial phase of angiogenesis. 10 In a recent study by Kim et al., it was found that autocrine TGF-β1-Smad2 cross-talk pathway is involved in the differentiation of hASCs to contractile SMCs induced by angiotensin-2. 25 On the other hand, the role of BMP4 in development has been generally accepted as an important instrumental signal in regulating the induction of cartilage and bone in limb growth rather than a regulator in vasculature development.37,38 It was till recent years that the role of BMP4 in vascular and hematopoietic development was recognized. In vitro, cultured endothelial cells have been shown to express BMP4 when exposed to both cytokines and oscillatory shear stress. 39 In vivo, expression of BMP4 was found to be upregulated in endothelium during arterial remodeling, and further, activating the BMP signaling cascade resulted in the decreased proliferation and migration of SMCs. 40 These studies suggest that BMPs may elicit a protective role in counterbalancing the effects of mitogens and chemoattractants that contribute to pathologic arterial remodeling. Moreover, it was shown that when exposed to BMP4, human embryonic stem cells expressed trophoblast markers and outgrowth of capillary-like structures was observed, indicating the presence of mature endothelial cells and α-SMA-positive SMCs. 41 On the basis of these studies, BMP4 was chosen as a costimulator with TGF-β1 to induce the differentiation of hASCs to SMCs.
The mechanism(s) responsible for SMC differentiation of hASCs induced by TGF-β1 and BMP4 in combination remain unclear. TGF-βs are known to modulate SMC differentiation by directly binding to type-I receptor and thereafter activating downstream signals of Smad proteins.17,42 On the other hand, it seems that BMP4 may initiate SMC differentiation of hASCs in an indirect way which is independent on the TGF-β pathway. According to the report by Lagna et al., 22 the SMC phenotype switch induced by BMP4 from synthetic to contractile ones requires intact RhoA/ROCK signaling which is not blocked by inhibitors of the TGF-β pathways. Therefore, we speculate that a cross-talk between BMP and TGF-β pathways may contribute to the regulation of SMC differentiation from hASCs, but the detailed underlying mechanism needs to be further investigated. Moreover, cell proliferation assay showed that hASCs treated with growth factor exhibited a growth arrest as that cultured in BM.
Previous studies have demonstrated that BMP4 could participate in in vivo endochondral ossification and is one of the main local contributing factors in the early stage of bone repair. 43 Transfer of adenoviral-mediated BMP4 gene to mouse ASCs resulted in their osteogenic differentiation and bone formation was observed when the transduced cells were injected in vivo. 44 Thus, one major concern in stimulating the differentiation of hASCs into SMC lineage with BMP4 is that treated hASCs may undergo osteogenic differentiation in parallel, which would lead to calcification of the engineered vessel constructed using such differentiated hASCs. As a marker indicating early differentiation of osteoblasts, ALP was shown to regulate organic or inorganic phosphate metabolism via the hydrolyzation of phosphate esters and to function as a plasma membrane transporter for inorganic phosphates. OCN is also an established indicator of the osteogenic differentiation and is synthesized only by mature osteoblasts to bind both collagen and calcium in the extracellular matrix of bone tissue, whereas Cbfa1 is known as an osteoblast transcriptional activator which is identified as a key regulator of osteoblast differentiation. The function of Cbfa1 is dominant during osteoblast differentiation, that is, its regulation of the expression of major osteoblast genes and its role in maintaining the functions of differentiated osteoblasts at an early stage. In this study, the expression of osteogenic related genes including ALP, OCN, and Cbfa1 could not be detected in hASCs treated with BMP4 and TGF-β1 in combination.
To ascertain a differentiated SMC phenotype, not only expression of the contractile structural proteins should be evaluated, but also the special functional property of contraction elicited by induced cells in response to various stimuli needs to be demonstrated. Thus, a collagen lattice contraction assay was performed to assess the contractile property of differentiated hASCs induced by TGF-β1 and BMP4 in combination. It was found that induced hASCs elicited a remarkable contraction of collagen gel lattice. In addition, the collagen gel embedded with differentiated hASCs underwent a further shrinkage in response to the muscarinic contractile agonist carbachol. Hence, these data suggest that hASCs can be induced to differentiate into an SMC phenotype with contractile function when stimulated by TGF-β1 and BMP4.
Conclusion
It was demonstrated in this study that expanded hASCs could be induced to differentiate along an SMC pathway by TGF-β1 and BMP4 stimulation for 7 days in vitro, as evidenced by expression of SMC-specific transcripts and proteins including α-SMA, calponin, SM22α, and SM-MHC. Such differentiated hASCs when embedded in collagen lattice displayed contraction which was further strengthened in response to carbachol stimulation. These results substantiate the possibility of using hASCs as a promising candidate for treating cardiovascular diseases and for blood vessel engineering purposes.
Footnotes
Acknowledgments
This work was financially supported by the Doctoral Innovation Foundation of Shanghai Jiao Tong University School of Medicine (grant no. BXJ0824), Doctoral Degree Foundation of China (grant no. 200802480072), and National “973” Project Foundation (grant no. 2005CB522700), Natural Science Foundation of China (grant no. 30970743/C100202) and National “863” Project Foundation (grant no. 2009AA02Z110).
Disclosure Statement
No competing financial interests exist.