
Abstract
Scaffold-free cultures provide promising potential in chondrogenic differentiation of human mesenchymal stem cells (hMSCs). In this study, a novel scaffold-free membrane-based culture system, in which hMSCs were cultivated on a cellulose acetate membrane filter at medium–gas interface, was evaluated for chondrogenesis under the addition of growth factors. Chondrogenic differentiation of hMSCs has been described in scaffold-free pellet cultures with good results. In our study membrane-based cultures (1 × 106 hMSCs) were produced, maintained at the medium–gas interface and cultured for 21 days. Results were compared with findings from standard pellet cultures (2.5 × 105 hMSCs). The effects of the following growth factors were examined: human transforming growth factor-β3 (TGF-β3) ± insulin-like growth factor-1 or ± human fibroblast growth factor 2. After 3 weeks of culture, chondrogenesis was assessed by Safranin-O staining, immunohistochemistry, a dimethylmethylene blue dye binding assay for glycosaminoglycans, and quantitative real-time polymerase chain reaction for cartilage-specific proteins. Membrane-based cultures containing growth factors formed hemispherical structures with a large surface area (65 mm2). When removed from the membrane they showed a histologically smooth cartilage-like surface. Membrane-based cultures stained positive for Safranin-O and collagen type II and contained a high content of glycosaminoglycans. Expression of cartilage-specific markers like collagen type II, aggrecan, and SOX9 was observed under the addition of TGF-β3, whereas combinations of growth factors let to a significant increase of collagen type II expression. A markedly reduced expression of collagen type X was found in membrane-based cultures when only TGF-β3 was added. Pellet cultures showed similar results besides an increased expression of collagen type X and type II that were observed. Membrane-based cultures provide a differentiation system, comparable in chondrogenesis to pellet cultures, which is able to generate scaffold-free neocartilage. The key benefit factors of membrane-based cultures are a histologically smooth cartilage-like surface and reduced expression of collagen type X, both of which are suitable features for its future application in cartilage regeneration.
Introduction
Materials and Methods
Monolayer culture
Adult human bone marrow MSCs were derived from Lonza (Walkersville, MD), seeded in monolayer triple flasks (Nunc, Roskilde, Denmark) at a density of 2 × 103 cells/cm2 and grown at 37°C in a humidified atmosphere containing 95% air and 5% CO2 in a growth medium (alpha-medium [Biochrom, Berlin, Germany] containing 10% fetal calf serum [PAA, Pasching, Austria], 50 U/mL penicillin/streptomycin [Biochrom], and 5 ng/mL recombinant human fibroblast growth factor-2 [FGF-2; Pepro Tech, Rocky Hill, NJ]), which was shown to promote the chondrogenic potential. 9 Medium was replaced every second day. Cells were passaged at 70% confluence using 0.05% trypsin containing 0.02% ethylenediaminetetraacetic acid (Biochrom).
Pellet culture
About 2.5 × 105 hMSCs (passage 5) were centrifuged (150 g, 5 min) in a 15 mL polypropylene tube (TPP, Trasadingen, Switzerland) to form pellet cultures. They were cultured for 21 days in 1 mL differentiation medium consisting of high-glucose Dulbecco's modified Eagle's medium (Biochrom) supplemented with 10 mg/L insulin, 5.5 mg/L transferrin, 5 μg/L selenium, 0.5 mg/mL bovine serum albumin, 4.7 μg/mL linoleic acid, 0.1 μM dexamethasone, 0.2 mM L-ascorbic acid-2-phosphate, 0.35 mM L-proline (all Sigma-Aldrich, Steinheim, Germany), and 30 U/mL penicillin/streptomycin (Biochrom) under standard conditions as indicated above.
Additional growth factors were added to the differentiation medium in four groups: (1) 10 ng/mL recombinant human TGF-β3 (R&D Systems, Wiesbaden, Germany) and 100 ng/mL insulin-like growth factor-1 (IGF-1; PeproTech, Rocky Hill, NJ); (2) 10 ng/mL TGF-β3 and 5 ng/mL FGF-2; (3) 10 ng/mL TGF-β3; and (4) no additional growth factor (control).
Membrane-based culture
About 1 × 106 hMSCs (passage 5) were centrifuged (450 g, 5 min) in a 15 mL polypropylene tube, pipetted as a pellet onto a 8 × 20 mm piece of cellulose acetate membrane filter (Sartorius, Göttingen, Germany; pore size 0.2 μm), maintained at the medium–gas interface by a stainless steel grid in a Petri dish (Nunc) and cultured for 21 days. Differentiation medium with additional growth factors according to the four subsets above was filled up to the level of the grid, and medium change was carried out at 2-day intervals.
Histology
Cultures were rinsed in phosphate-buffered saline (pH 7.4), incubated in 5% sucrose (4°C, 30 min), embedded in Tissue-Tek (Sakura, Zoeterwoude, The Netherlands), and frozen in liquid nitrogen. Serial cryosections (10 μm) were cut, mounted on SuperFrost glass slides (Menzel-Gläser, Braunschweig, Germany), fixed in acetone, and dried at room temperature. Serial sections were stained with 1% Toluidine Blue (Chroma, Münster, Germany) or with Safranin-O (Fluka, Buchs, Switzerland) and fast green (Chroma) as described 12 to estimate the distribution of proteoglycans.
The morphology of pellet and membrane-based cultures was graded according to a visual histological grading system for generated neocartilage 13 by two independent and blinded observers (Table 1).
Three categories were evaluated: (A) intensity of Safranin-O staining, (B) distance between cells/amount of matrix that was accumulated, and (C) cell morphologies (rounded/spindle shaped) represented.
TGF-β3, transforming growth factor-β3; FGF-2, fibroblast growth factor 2; IGF-1, insulin-like growth factor-1.
Immunohistochemistry
Endogenous tissue peroxidase activity in sections was blocked (10 min, 0.3% H2O2 in methanol). To remove glycosaminoglycans (GAGs) and thereby permeabilize the tissue to antibodies, sections were then treated with Chondroitinase ABC from Proteus vulgaris (Sigma-Aldrich) (0.25 U/mL) in Tris pH 7.0 for 15 min at 37°C followed by washing in phosphate-buffered saline. Nonspecific binding sites were blocked with 5% sheep serum. Afterward tissue sections were incubated at 4°C overnight with a monoclonal mouse antibody (AB) against collagen type II (CIIC1, Developmental Studies Hybridoma Bank, University of Iowa, IA) diluted 1:6 or a monoclonal mouse AB against collagen type I (Sigma, St. Louis, MO), both diluted in Tris-buffered saline (TBS), always containing 0.1% Tween 20 (Merck, Darmstadt, Germany) and 1% bovine serum albumin. For negative controls, the first AB was omitted. Sections were washed in TBS and incubated with a biotinylated anti-mouse IgG (Vector Laboratories, Burlingame, CA) diluted 1:200 in TBS for 1 h at room temperature. After repeated washing, the section-bound AB was viewed with 3-amino-9-ethylcarbazole, counterstained with hematoxylin and embedded in Aquamount (BDH Laboratory Supplies, Poole, United Kingdom). Images were obtained using a Zeiss Axioskop 40 (Carl Zeiss, Jena, Germany) equipped with an AxioCam MRc 5 and AxioVision Software Rel. 4.5.
Determination of DNA and GAG content
Cultures were digested at 65°C for 16 h using 10 μg/mL of proteinase K (Sigma-Aldrich) in 30 mM Tris (pH 7.8), 50 mM NaCl, and 10 mM MgCl2. Culture digests were carried through three freeze–thaw cycles. DNA content of digests was determined by the fluorescence-based Hoechst Dye 33258 method 14 using the QuantiFluo™ DNA Assay Kit (BioAssay Systems, Hayward, CA) with a standard curve established with calf thymus DNA. Fluorescence was evaluated with FLUOstar OPTIMA (BMG Labtech, Offenburg, Germany), including Optima software 2.10. GAG content of digests was determined using the dimethylmethylene blue dye binding assay. 15 Briefly, dimethylmethylene blue solution was added to aliquots of digests followed by determination of absorbance at 595 nm. A standard curve was established with chondroitin-6-sulfate from shark cartilage (Sigma-Aldrich). Results were normalized to the DNA content of each sample.
Quantitative real-time polymerase chain reaction
In a LightCycler® (Roche Applied Science, Mannheim, Germany), the following primers were used: cyclophylin B, collagen type II α1 chain (COL2A1), type I α1 chain (COL1A1) and type X α1 chain (COL10A1), aggrecan, SRY (sex-determining region Y)-box 9 (SOX9), and cartilage oligomeric matrix protein (COMP), all purchased from Search-LC (Heidelberg, Germany); the sequences are not provided by the company. Cultures were disrupted under frozen conditions at 100 rpm using a Micro-Dismembrator S (Sartorius). Total RNA was directly isolated from freeze-milled preparations using the RNeasy Lipid Tissue Kit (Qiagen, Hilden, Germany) including RNase-free DNase treatment. For cDNA synthesis, 1 μg of total RNA was reverse transcribed using the Transcriptor High Fidelity cDNA Synthesis Kit (Roche Applied Science). Amplification reactions were performed with a LightCycler-Fast Start DNA Master SYBR Green I kit (Roche Applied Science). Two microliters of diluted sample was used for a quantitative two-step polymerase chain reaction (PCR) (a 10-min step at 95°C, followed by 35 cycles of 10 s at 95°C, 10 s at 68°C, and 16 s at 62°C). Reactions were performed in triplicates; relative quantification of the target gene expression was generated normalizing to cyclophylin B.
Statistical analysis
Results were represented as the mean ± standard deviation. All experiments were repeated at least twice, unless otherwise specified. Statistical analysis was performed using Mann–Whitney U test.
Results
Histology and immunohistochemistry
Pellet cultures
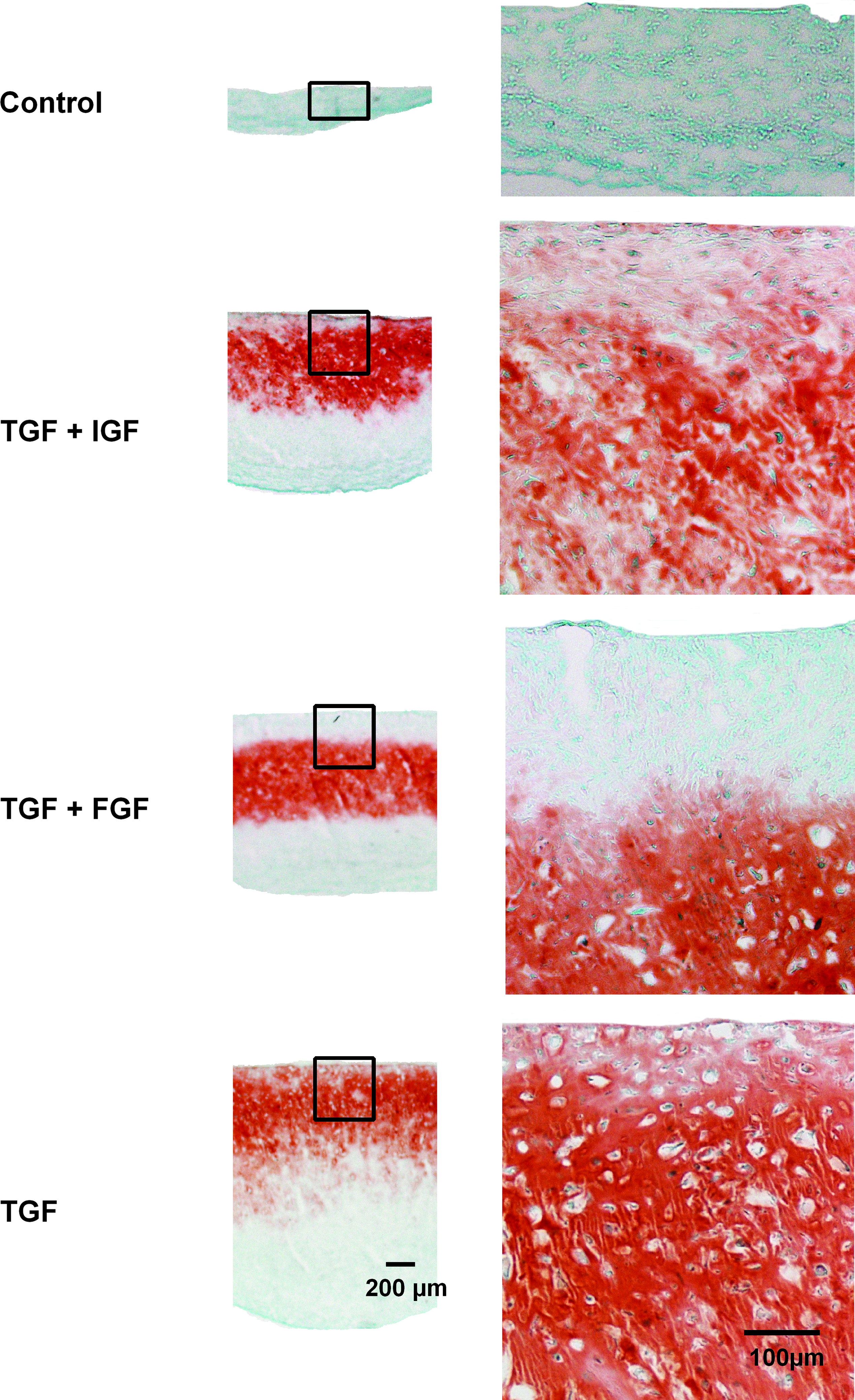
At the end of the differentiation period, spheroid pellet cultures had formed with a maximum diameter of 1.8 mm, control cultures reaching a diameter of 0.5 mm. Positive staining for Safranin-O and collagen type II was found in the central part of all cultures treated with growth factors. Cultures treated with combinations of growth factors revealed reduced staining of Safranin-O and collagen type II up to 150 μm from their final margins. When growth factors were omitted, there was no staining for Safranin-O and collagen type II detected. Pellet cultures did show a uniform staining for collagen type I for all groups of growth factors (Fig. 1).

Safranin-O staining and immunohistochemistry for collagen type II and type I of pellet cultures treated with no additional growth factor, TGF-β3 and IGF-1, TGF-β3 and FGF-2, or TGF-β3. TGF-β3, transforming growth factor-β3; FGF-2, fibroblast growth factor 2; IGF-1, insulin-like growth factor-1. Color images available online at www.liebertonline.com/ten.
Membrane-based cultures
At 21 days membrane-based cultures treated with growth factors had formed hemispherical constructs that were similar to hyaline cartilage with respect to palpation and texture (Fig. 2B). The cultures were easily removed intact from the membrane filter. A diameter of 5.5 mm and a surface of 65 mm2 were reached for all membrane-based cultures treated with growth factors (Fig. 2C, D). Membrane-based cultures grew to a thickness of 1.8 mm, when only TGF-β3 was added (Fig. 3). For combinations of growth factors, a thickness of 1.3 mm was reached (Fig. 3). Control cultures reached a thickness of 0.3 mm and 2.6 mm diameter (Fig. 3).
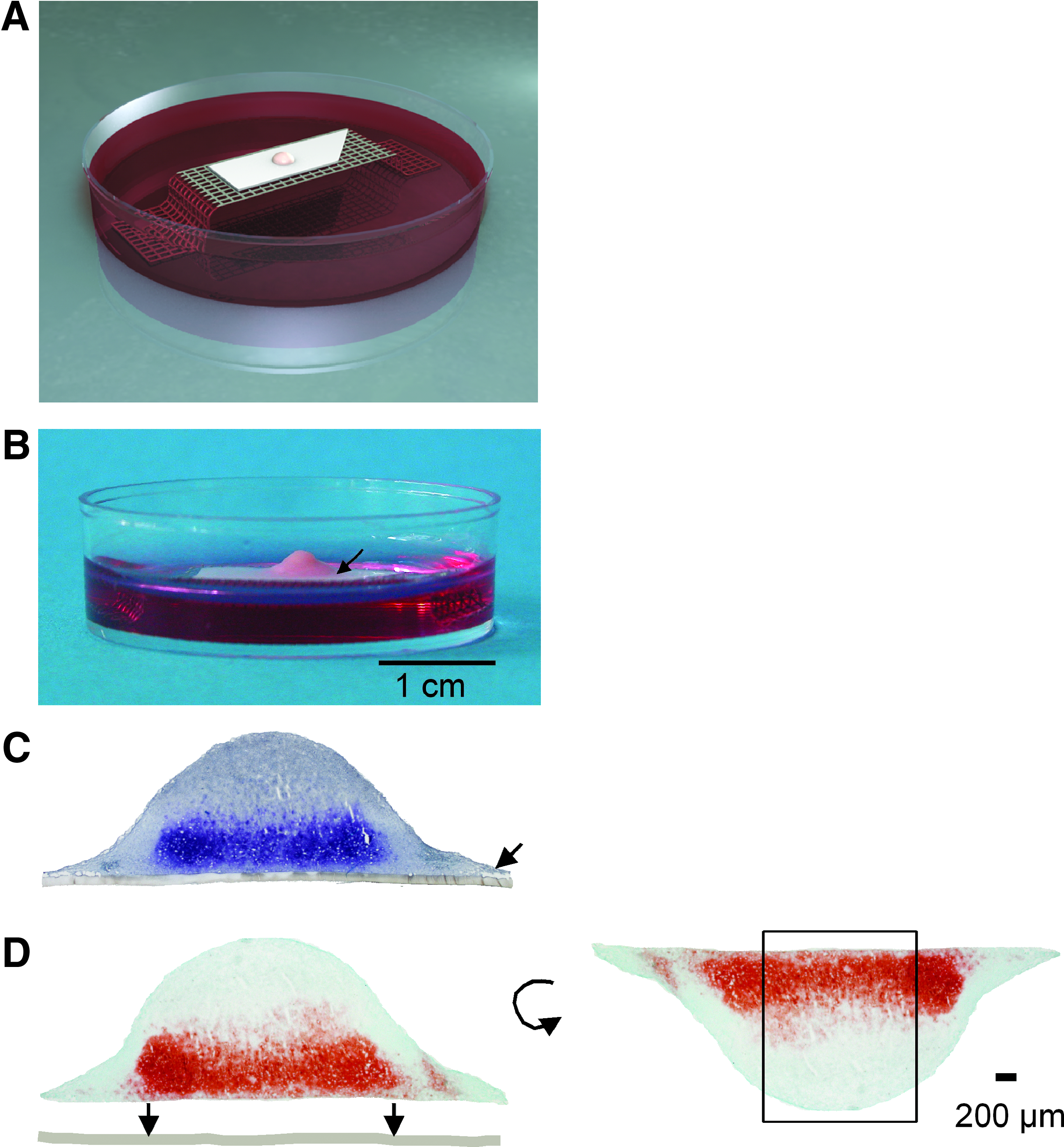
Schematic drawing (
Safranin-O staining of membrane-based cultures treated with no additional growth factor, TGF-β3 and IGF-1, TGF-β3 and FGF-2, or TGF-β3. Color images available online at www.liebertonline.com/ten.
All membrane-based cultures treated with growth factors showed a histologically smooth surface and a zonal distribution of Safranin-O and collagen type II staining: a positive staining was found starting at the membrane up to a distance of 700 μm when cultures were treated with TGF-β3 ± IGF-1 (Figs. 3 and 4). Cultures treated with TGF-β3 + FGF-2 showed a reduced signal for collagen type II and no signal for Safranin-O adjacent to the membrane, and a positive signal was found at a distance of 200–700 μm from the membrane (Figs. 3 and 4).
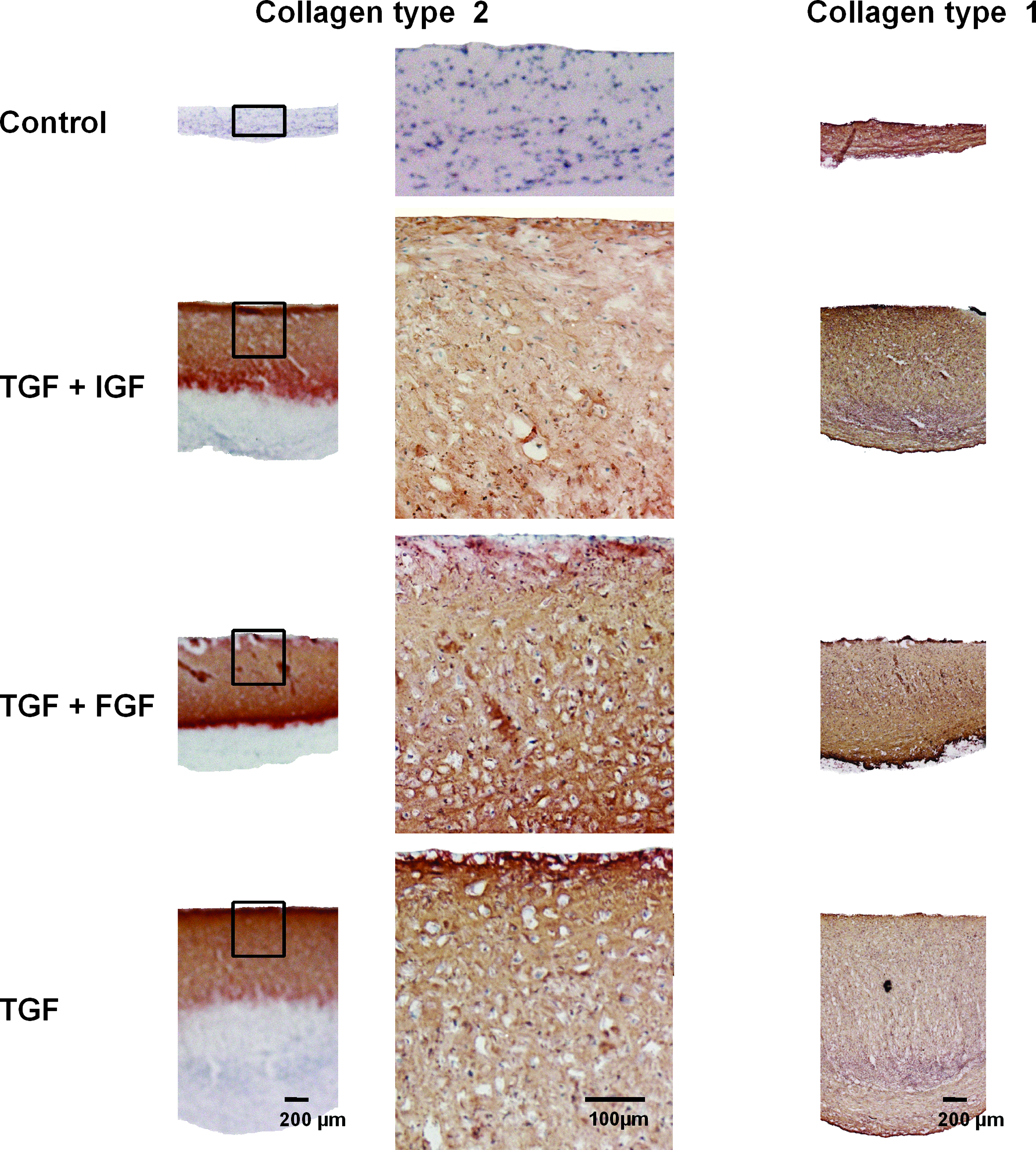
Immunohistochemistry for collagen type II and type I of membrane-based cultures treated with no additional growth factor, TGF-β3 and IGF-1, TGF-β3 and FGF-2, or TGF-β3. Color images available online at www.liebertonline.com/ten.
All membrane-based cultures showed a loss of staining for Safranin-O and collagen type II up to 150 μm from the lateral culture edges (Fig. 2D). Control cultures showed no staining for Safranin-O or collagen type II (Figs. 3 and 4). A uniform distribution of staining for collagen type I could be detected throughout all membrane-based cultures (Fig. 4).
Visual histological grading system
According to the histological grading system, the cultures showed the most cartilage-like morphology with strong staining for collagen type II, rounded chondrogenic cells, and excessive extracellular matrix around cells when only TGF-β3 was added (Table 1).
GAG/DNA content and quantitative real-time PCR pellet cultures that had been added TGF-β3 provided the highest GAG/DNA content. For pellet cultures, a significantly reduced GAG/DNA content was detected in cultures treated with TGF-β3 and IGF-1 compared with membrane-based cultures. The GAG/DNA content of membrane-based cultures showed similar results for the different groups of growth factors (Fig. 5).
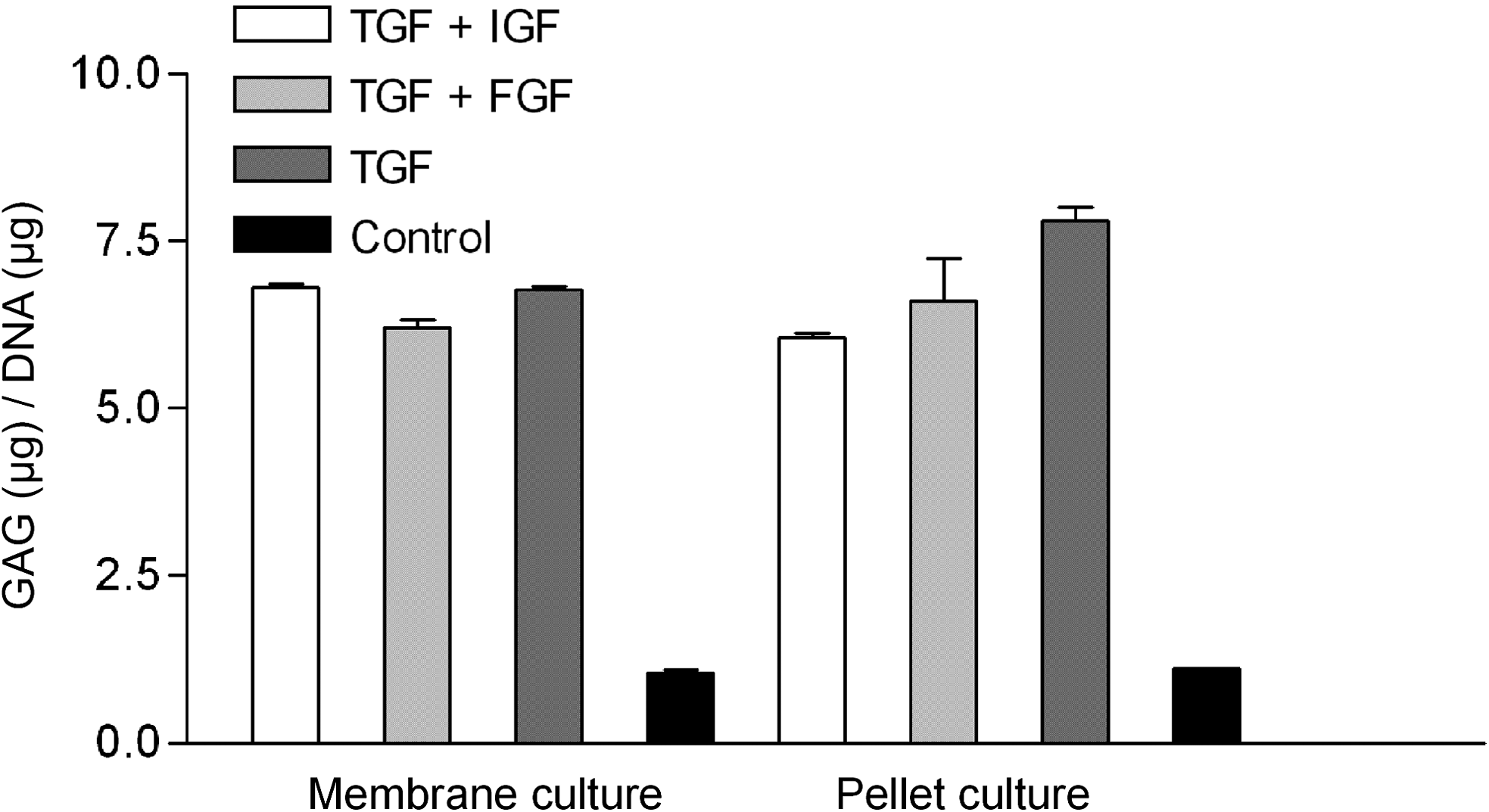
Relative amount of glycosaminoglycan/DNA content in membrane-based and pellet cultures treated with TGF-β3 and IGF-1, TGF-β3 and FGF-2, TGF-β3, or no additional growth factor (control).
Detection of aggrecan, COL2A1, COMP, and SOX9 mRNA by quantitative real-time PCR confirmed a chondrogenic phenotype of pellet and membrane-based cultures, which had been added growth factors. Control pellet cultures did not express any cartilage markers, whereas membrane-based cultures expressed COL2A1, aggrecan, COMP, and COL10A1 mRNA at basal levels. Aggrecan, COL2A1, and COMP mRNA expressions were found to significantly preponderate in pellet cultures under the addition of growth factors compared with equally treated membrane-based cultures. Combinations of TGF-β3 with IGF-1 or FGF-2 significantly enhanced the expression of COL2A1 in both culture systems. The combination of TGF-β3 and FGF-2 significantly increased the expression of SOX9 in pellet cultures. For membrane-based cultures, the highest expression of SOX9 was found in cultures treated with TGF-β3. COL10A1 showed the significantly lowest level of mRNA expression in membrane-based cultures treated with TGF-β3. For membrane-based cultures treated with TGF-β3 and IGF-1, the COL10A1 expression was found to be significantly decreased compared with pellet cultures. The combination of TGF-β3 and FGF-2, however, caused a higher expression of COL10A1 in membrane-based than in pellet cultures (Fig. 6). The lowest expression of COL1A1 mRNA was found in control cultures, followed by membrane-based cultures, which had been added TGF-β3 also in combination with IGF-1 (Fig. 6).

Expression of mRNA in membrane-based and pellet cultures treated with TGF-β3 and IGF-1, TGF-β3 and FGF-2, TGF-β3, or no additional growth factor (control).
Discussion
This study demonstrated that hMSCs cultured on cellulose acetate membrane filters have the ability to differentiate into chondrocytes and form an extracellular matrix rich in collagen type II and proteoglycans dependent on the use of appropriate growth factors. Membrane-based cultures starting with 1 million cells formed hemispherical structures with a large and histologically smooth surface, which reached a surface area of 65 mm2 after 3 weeks of culture.
Pellet cultures made from 250,000 cells are an established chondrogenic assay, which has been used to examine the effects of various growth factors on the control of the differentiation process. 3 In this study, the standard type of pellet culture system with a defined cell quantity was used. We refrained from increasing the cell number to 1 × 106 cells, as it is known that pellet cultures made from 5 × 105 cells already show a strong loss of collagen type II production in their central region. 16 This has not been observed for smaller pellet cultures, and therefore the small pellet size was applied to gain optimal results.
Although pellet cultures have shown rather good results concerning chondrogenic differentiation, their round structure limits the clinical application to restore a histologically smooth cartilage-like surface and dictated the need for improvements in the development of scaffold-free cultures. Membrane-based cultures offer the advantage of generating a histologically smooth surface, eligible to cover, for example, cartilage defects of 4 cm2 by about 6 million cells. The smooth surface of membrane-based cultures might be ascribed to the cellulose acetate membrane filter. Cellulose acetate filters, which are also used for impression cytology in ophthalmology, are able to transfer cells without rupture. In impression cytology, cells become firmly attached to the filter. When the filter is removed, there is no damage to cell membranes being detectable in scanning electron microscopy. 17 It can be assumed that cellulose acetate membrane filters provide a surface for hMSCs in membrane-based cultures, causing a primary attachment of cells which is easy to be removed later not resulting in cell rupture. Membrane-based cultures primarily grow on the plane cellulose acetate membrane and keep their shape when the membrane is removed without cell damage. This might be the basic principle for a histologically smooth surface found in membrane-based cultures. Regarding the expression of cartilage markers, our results were consistent with literature as combinations of growth factors significantly enhanced the expression of COL2A1 in pellet cultures.8,18 The same enhancement of COL2A1 expression was observed for combinations of growth factors in membrane-based cultures. However, an interesting finding was observed for membrane-based cultures treated with TGF-β3. Our results demonstrated a markedly reduced expression of collagen type X and a sustained upregulation of Sox9 in membrane-based cultures incubated with TGF-β3. These cultures also obtained the best results regarding the histological grading of neocartilage. TGF-β plays a central role in chondrogenesis, with TGF-β2 and TGF-β3 being more efficient in inducing chondrogenesis compared with TGF-β1. 19 TGF-β3 is known to suppress collagen type X expression of chondrocytes in cartilage explants, 20 which has not been described for scaffold-free stem-cell cultures so far. 16 Collagen type X, which is a marker for hypertrophy, is one of the main problems of in vitro cartilage generation. 21 Cartilage development involves a range of exactly regulated events, encompassing expression of Sox9 that negatively regulates the transition of chondrocyte proliferation into hypertrophy and is essential for matrix protein gene expression and the maintenance of the chondrocyte phenotype. 22 Reduced collagen type X expression in combination with a strong expression of Sox9 in membrane-based cultures treated with TGF-β3 implies a diminished hypertrophy of chondrogenic cells generated from hMSCs in membrane-based cultures. The reduced expression of COL1A1 mRNA in membrane-based cultures further leads to the hypothesis that the expression of unspecific proteins is suppressed in this culture system.
There are several factors that might induce the beneficial effect described for membrane-based cultures. First, the low solubility of oxygen caused a depressed oxygen tension in the center of diffusion cultures, which had already been described to produce central necroses in former organ cultures. 11 Membrane-based cultures made from hMSCs that undergo chondrogenic differentiation, like in this study, react differently to the hypoxic conditions found in the center of cultures. Since chondrogenesis is enhanced by hypoxic conditions in vitro, 23 the chondrogenic differentiation of hMSCs was presumably supported in central membrane-based cultures. Second, it is well known that composition and matrix elasticity of a substratum play an important role in developing differentiation properties of hMSCs. 10 High-density cultures on cellulose acetate membrane filters have been described to facilitate redifferentiation of chondrocytes. 24 Therefore and according to our results, it can be assumed that membrane-based cultures experience a promotion of chondrogenesis which is comparable to that of pellet cultures but can suppress the expression of collagen type X.
Nevertheless, membrane-based cultures showed a lower expression of COL2A1, aggrecan, and COMP mRNA compared with pellet cultures. An explanation for a reduced collagen type II expression at the culture edges of membrane-based cultures might be tensile forces caused by the hemispherical form of cultures. However, this concurs with a reduced signal for collagen type II and Safranin-O at the margin of pellet cultures and does not explain the differences seen in collagen type II expression. Therefore, it is necessary to look at the zonal distribution for collagen type II and Safranin-O found in membrane-based cultures. Regarding this zonal distribution, the relative reduction in COL2A1, aggrecan, and COMP expression might be related to the reduced amount of cells producing this cartilage-specific proteins in the upper part of the cultures. We assume that either the direct oxygen contact at the surface of cultures abolished collagen type II, aggrecan, and COMP expression or that the diffusion capacity of growth factors did not reach the upper part of the membrane-based cultures. Therefore, further studies with lowered oxygen tension (5%) seem necessary to examine the influence on the expression of these proteins.
As pellet cultures of a smaller size were used, it is possible that similar reductions of COL2A1, aggrecan, and COMP expression would have been observed in the center of pellet cultures made of 1 × 106 cells. We therefore are not able to compare the overall pellet culture conditions to membrane-based cultures but merely state that under standard conditions pellet cultures expressed more COL2A1, aggrecan, and COMP than membrane-based cultures.
In conclusion, membrane-based cultures showed comparable results to pellet cultures concerning the chondrogenic differentiation potential, although there was a reduced expression of collagen type II observed. According to our results, cellulose acetate membrane-based cultures enhanced the chondrogenic differentiation of hMSCs under the addition of TGF-β3 and reduced the expression of genes associated with chondrocyte hypertrophy. Membrane-based cultures offer in addition a large and histologically smooth surface area, which would be suitable for the regeneration of cartilage surfaces. Pellet cultures, however, which form classical spherical structures, might be preferred to fill up the depth of cartilage defects.
Membrane-based cultures show the clinical advantage of not being based on scaffolds. Scaffolds might cause problems regarding retention or degradation and the risk of immunological reactions toward the material. Moreover, scaffold-based cultures have to be seeded with cells. This might be a handicap, particularly in the case of large scaffolds the infiltration rate can be very low, resulting in poor loading and an uneven distribution of cells within the scaffold. These disadvantages are not found in scaffold-free membrane-based cultures.
As cartilage repair is a common medical problem, membrane-based and pellet cultures generated from hMSCs under the stimulation of TGF-β3 might be used for clinical applications as scaffold-free neocartilage in the future.
Footnotes
Acknowledgments
We thank Julia Redeker and Bärbel Schmitt for technical support, Lars Schliack for graphical assistance (![]() ), Prof. M. Shakibaei for help with the membrane-based culture system, and Prof. R. A. Poole for important assistance with the manuscript. This research was supported by the “Förderung für Forschung und Lehre” program from the University of Munich. The authors indicate no potential conflict of interests. This work was presented by Mr. Schiergens in partial fulfillment of the requirements for an M.D. degree, Ludwig-Maximilians-University Medical School, Munich, Germany.
), Prof. M. Shakibaei for help with the membrane-based culture system, and Prof. R. A. Poole for important assistance with the manuscript. This research was supported by the “Förderung für Forschung und Lehre” program from the University of Munich. The authors indicate no potential conflict of interests. This work was presented by Mr. Schiergens in partial fulfillment of the requirements for an M.D. degree, Ludwig-Maximilians-University Medical School, Munich, Germany.
Disclosure Statement
No competing financial interests exist.