
Abstract
Stromal cells residing in murine fetal livers have the ability to promote the hepatic maturation of murine embryonic stem cells (ESCs) and hepatic progenitor cells (HPCs) in vitro. These stromal cells were isolated as the CD49f(+/−)CD45(−)Thy1(+)gp38(+) cell fraction. The present study established a murine fetal liver stromal cell line that induced hepatic maturation in mouse ESCs and HPCs. A transgene containing a temperature-sensitive SV40 large T antigen was transfected into the primary fetal liver stromal cells. These immortalized cells, which were named as the gp38-positive and Thy1-positive murine liver stromal (MLSgt) cells, induced both mouse ESCs and HPCs to differentiate into mature hepatocyte-like cells using a coculture method. Since MLSgt is not a cloned cell line, one clone, MLSgt20, was selected as a line with the characteristic to induce hepatic differentiation, which was comparable to its parental stromal cells. The ESC-derived endoderm cells cocultured with the MLSgt20 cells expressed mature hepatocyte-specific gene markers, including glucose-6-phosphatase, tyrosine aminotransferase, tryptophan 2,3-dioxgenase, and cytochrome P450 (CYP1a1, Cyp1b1, Cyp1a2, and Cyp3a11). In addition, these cells also exhibited hepatic functions, such as glycogen storage and ammonia metabolism. Transmission electron microscopy showed that the cocultured ESCs expressed the morphologic features of mature hepatocytes. In conclusion, a cell line was established that has the characteristic to promote the hepatic maturation of mouse ESCs and HPCs by a coculture method.
Introduction
Our laboratory previously reported that mouse primary CD49f(+/−)CD45(−)Thy1(+) stromal cells from a fetal liver promoted the maturation of hepatic progenitor cells (HPCs) in vitro. 21 Further investigation revealed that this stromal cell fraction was divided into two populations. The gp38-positive cell fraction (Thy1(+)gp38(+) stromal cell) had the ability to promote hepatic maturation of HPCs, whereas the gp38-negative cell fraction did not. 22 Further, mouse ESCs differentiate into mature hepatocyte-like cell after coculture with the CD49f(+/−)CD45(−)Thy1(+) stromal cells. 23
From a practical point of view, cell transplantation therapy or a bioartificial liver would require a large number of hepatocytes. Therefore, large-scale coculture of ESC and Thy1(+)gp38(+) stromal cells would be necessary. To establish a stable coculture system for obtaining a sufficient number of ESC-derived hepatocytes, it would be desirable to establish a cell line that stimulated the differentiation of ESCs into mature hepatocyte-like cells. The present study was designed to establish a cell line utilizing the Thy1(+)gp38(+) stroma cells, and to establish a culture method that differentiated mouse ESCs into functional hepatocyte-like cells. Our culture method is partially based on the knowledge of the physiological liver development. This cell line would also help elucidate the novel molecular mechanism of the coculture system. The present study was designed to establish a cell line that had the characteristic to promote hepatic maturation.
Materials and Methods
Animals
C57BL/6J mice were purchased from SLC (Hamamatsu, Japan). All animal experimental procedures were performed in accordance with Animal Protection Guidance of Kyoto University and Stem Cell and Drug Discovery Institute.
Flow cytometry and cell sorting
Fetal HPCs were obtained from E13.5 fetal livers and enriched using the cell aggregate formation method previously described. 24 The stromal cells were sorted out as the CD49f(+/−)CD45(−)Thy1(+)gp38(+) cell population using FACSAria (BD Biosciences Immunocytometry Systems, San Jose, CA), and these cells were cultured in a hepatocyte differentiation medium (HD medium; Dulbecco's modified Eagle's medium with 10% fetal calf serum [JRH Biosciences, Lenexa, Kansas], 1 mM sodium pyruvate [Sigma-Aldrich, St. Louis, MO], 10 mM nicotinamide [Sigma-Aldrich], 2 mM L-ascorbic acid phosphate [Wako Pure Chemical, Osaka, Japan], insulin–transferrin–selenium supplement [Gibco, Grand Island, NY], 1 × 10−7 M dexamethasone [Sigma-Aldrich], 20 ng/mL hepatocyte growth factor [R&D Systems Inc., Minneapolis, MN], and penicillin/streptomycin) on collagen type I–coated dishes (AGC Techno Glass, Chiba, Japan). The following antibodies were used: anti-CD49f-phycoerythrin, CD45-phycoerythrin, and Thy1-fluorescein isothiocyanate (BD Biosciences). Anti-mouse gp38 antibody (Medical and Biological Laboratories, Nagoya, Japan) was conjugated with allophycocyanin by the conjugation kit (Prozyme, San Leandro, CA).
Generation of immortalized stromal cells derived from mouse liver stroma
A plasmid vector containing a temperature-sensitive SV40 large T antigen (pSVtsA58 ori(−); RIKEN BioResource Center, Tsukuba, Japan) as an immortalizing gene was transfected into the primary mouse CD49f(+/−)CD45(−)Thy1(+)gp38(+) stromal cells derived from fetal liver tissues by Lipofectamine 2000(Invitrogen, Carlsbad, CA) according to the manufacture's protocol. The cells were cultured in the presence of 200 μg/mL G418 (Sigma-Aldrich) 3 days after transfection for 5 days. Surviving colonies were picked up and expanded in HD medium. The immortalized gp38-positive and Thy1-positive murine liver stromal cells (MLSgt) were cultured on collagen type I–coated dishes (BD Bioscience Discovery Labware, Bedford, MA) at 33°C and 5% CO2 in HD medium with 200 μg/mL G418.
Cell culture of mouse ESC–derived endoderm cells with CD49f(+/−)CD45(−)Thy1(+)gp38(+) stromal cell lines
A transgenic mouse ESC line that expressed enhanced green fluorescent protein and a hygromycin resistant gene under the control of alpha-fetoprotein (AFP) promoter was used. 25 The mouse ESCs were differentiated into the AFP-producing cells as previously described. 23 In brief, to induce endodermal differentiation, the ESCs were transferred to a serum-free and feeder layer–free culture on collagen type I–coated dishes (Becton Dickinson Labware, Bedford, MA) with 10 μM all-trans retinoic acid and 1 × 103 U/mL leukemia inhibitory factor (LIF) for 2 days, followed by the sequential addition of 20 ng/mL hepatocyte growth factor and 20 ng/mL basic fibroblast growth factor (Upstate, Lake Placid, NY) for 5 days. To obtain the AFP-producing cells, they were cultured in HD medium containing 50 μg/mL hygromycin for 3 days. In this protocol, approximately 90% of the residual cells were the AFP-producing cells after the drug selection as previously described. 12
The MLSgt cells were disseminated at a density of 3.0 × 104 cells/cm2 to utilize as feeder layer cells. The ESC-derived AFP-producing cells were disseminated at 5 × 104 cells/cm2 on the cell layer of the MLSgt cells. To compare and investigate the suitable culture conditions for the hepatic maturation, the ESC-derived AFP-producing cells were also cultured under various conditions. They were cultured on (1) collagen type I–coated dishes (BD Biosciences), (2) dishes coated with growth factor–reduced Matrigel (BD Biosciences) that was diluted at 1:80, (3) MLSgt cells that were fixed by 0.05% glutaraldehyde (Sigma-Aldrich) for 5 min and washed with phosphate-buffered saline three times and incubated overnight and then washed by 2 mg/mL glycine (Nakalai tesque, Kyoto, Japan) for 2 h, and (4) the cell layer of the SL-10 (subline of STO, mouse fibroblast cell line that we established previously 26 ) that was treated with 10 μg/mL mitomycin C for 2 h and then disseminated 3.0 × 104 cells/cm2. They were incubated at 37°C in 5% CO2 for 7 days in fresh HD medium or in conditioned medium from the MLSgt cells.
Reverse transcription–polymerase chain reaction
Total RNA was extracted using an RNeasy Mini Kit (Qiagen, Chatsworth, CA) and treated with RNase-free DNase (Qiagen). Total RNA (2 μg) was reverse transcribed into cDNA with oligo (dT) 12–18 primer (Invitrogen) using an Omniscript RT Kit (Qiagen). Polymerase chain reaction (PCR) was performed using Ex-Taq polymerase (Takara Bio Inc., Otsu, Japan). Primers were generated for the following mouse genes (oligonucleotide sequences are given in parentheses, followed by the annealing temperature): AFP (5′-GAAGATGGTGAGCATTGC C, 5′-AACAGACTTCCTGGTCCTGG, 58°C), glucose-6-phosphatase (G6P) (5′-TGCATTCCTGTATGGTAGTGG, 5′-GAATGAGAGCTCTTGGCTGG, 58°C), tyrosine aminotransferase (TAT) (5′-TCCAGGAGTTCTGTGAACAGC, 5′-AGTATATGGTGCCTGCCTGC, 60°C), tryptophan 2,3-deoxygenase (TO) (5′-GCTCAAGGTGATAGCTCGGA, 5′-GGA ACTCTGCCATCTGTTCC, 58°C), transthyretin (TTR) (5′-ATGAATTCGCGGATGTGG, 5′-TTCCTGAGCTGCTAACACGG, 60°C), alpha-1 antitrypsin (AAT) (5′-AGATAGCTGAGGCGGTGAAA, 5′-ATGGACAGTCTGGGGAAGTG, 60°C), and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (5′-ATTCAAGGGCACAGTCAAGG, 5′-ATCATAAACATGGGGGCATC, 60°C), α-smooth muscle actin (5′-CTATTCAGGCTGTGCTGTCC, 5′-AGTTCGTAGCTCTTCTCCAGG, 60°C), desmin (5′-GCTATCAGGACAACATTGCG, 5′-GTTGTTGCTGTGTAGCCTCG, 60°C), vimentin (5′-TACCAGGACACTATTGGCCG, 5′-CTGTTGCACCAAGTGTGTGC, 60°C), glial fibrillary acidic protein (5′-AGACAGGAGGCAGATGAAGC, 5′-AAGTGCCTCCTGGTAACTGG, 60°C), platelet endothelial cell adhesion molecule-1 (5′-CCACTTCTGAACTCCAACAGC, 5′-CCAACATGAACAAGGCAGC, 60°C), vascular endothelial cadherin (5′-AGGCTGAATACAAGATCGTGG, 5′-GGTCTGTCTCAATGGTGAAGG, 60°C), CD34 (5′-ACACATCATCTTCTGCTCCG, 5′-CTGTGCTATTGGCCAAGACC, 60°C), CD16 (5′-CCACAACTGGAGTTCCATCC, 5′-TTGTTCCTCCAGCTATGGCACC, 60°C), and cytochrome P450 3a11 (Cyp3a11) (5′-TGAGGCAGAAGGCAAAGAAA, 5′-GGTATTCCATCTCCATCACA, 60°C).
Quantitative PCR
For quantitative PCR, total RNA was reverse transcribed into cDNA using random primers (Invitrogen). Quantitative PCR was performed with a 7500 Fast Real-Time PCR System (Applied Biosystems, Foster City, CA) according to the manufacturer's protocol, using SYBR Green PCR Master Mix (Applied Biosystems). The program was 10 min at 95°C, 40 cycles of 15 s at 58°C, and 35 s at 72°C, followed by a dissociation curve step. The primers were generated for the following mouse genes: AFP (5′-CACACCCGCTTCCCTCAT, 5′-TTTTCGTGCAATGCTTTGGA, 58°C), G6P (5′-CAACCGCCATGCAAAGG, 5′-GGCCTCACAATGGGTTTCAG, 58°C), TO (5′-CCAGGGAGCACTGATGATATATTTT, 5′-GCTGGAAAGGGACCTGGAAT, 58°C), TTR (5′-TGGACACCAAATCGTACTGGAA, 5′-CATCCGCGAATTCATGGAA, 58°C), and GAPDH (5′-CGGCCGCATCTTCTTGTG, 5′-ACCGACCTTCACCATTTTGTCT, 58°C). The quantified values of each gene were normalized against GAPDH using the comparative threshold cycle method.
Functional analyses of the cultured cells
Functional analyses were performed to compare the ESC-derived AFP-producing cells cultured alone and cocultured with MLSgt cells. The MLSgt cells cultured alone and adult murine hepatocyte obtained from 10-week-old C57BL/6J mice were subject to be analyzed as a negative and a positive control, respectively. To examine the glycogen production and storage, periodic acid-Schiff (PAS) staining was performed as previously described.22,23 To investigate ammonia metabolism, cells were cultured on 12-well collagen I–coated dishes (BD Bioscience Discovery Labware) for 24 h in phenol red–free Dulbecco's modified Eagle's medium (Sigma-Aldrich) supplemented with 1.5 mM ammonium chloride (Wako Pure Chemical). After 7 days of culture, the ammonia concentrations in the culture media were measured using an Ammonia-Test Wako kit (Wako Pure Chemical), according to the manufacturer's protocol. Albumin concentrations in culture media were measured by sandwich ELISA method using Mouse Albumin ELISA KIT (Shibayagi, Gunma, Japan). The media were obtained from 24-h cultures of the cocultured ESC-derived AFP-producing cells, the ESC-derived AFP-producing cells cultured alone, MLSgt20 cells cultured alone, and primary murine hepatocytes. The cytochrome P450 1a1, 1b1, and 1a2 activities were assayed with the P450-Glo Assays Kit (Promega, Madison, WI), using luminogenic cytochrome P450 substrates, according to the manufacturer's protocol. To induce these three cytochrome P450s, 10 μM Omeprazole (Wako Pure Chemical) was added to the medium for 48 h before analysis. Luminescence was read by Lumat LB9507 (Belthold Technologies, Bad Wildbad, Germany), and the enzyme activities of cytochrome P450 were presented by relative light unit (RLU).
Transmission electron microscopy
The mouse ESC–derived AFP-producing cells were cocultured with the MLSgt cells, or cultured on collagen type I–coated dish as control, for 28 days. The cells were harvested, and a transmission electron microscopic analysis was performed as previously described. 21
Statistical analysis
All experiments were performed in triplicate. The results are presented as the means ± standard deviation (SD). Statistical analyses were performed by either Student's t-test with Bonferroni correction or Tukey's test. A p-value of <0.05 was considered to be statistically significant.
Results
Generation of stromal cell line–transfected SV40 large T antigen
The SV40 large T antigen was transfected to the primary mouse fetal stromal cells that were isolated as the CD49f(+/−)CD45(−)Thy1(+)gp38(+) cells. The G418-resistant cells were obtained, and the proper gene insertion was confirmed by PCR. The cells were able to proliferate and passaged more than 50 times (data not shown). These cells were assumed to proliferate at 33°C and be quiescent at 37°C because of their temperature-sensitive transgene. However, they could proliferate even at 37°C. Despite the confirmation of the proper gene insertion by reverse transcription (RT)-PCR (date is not shown), the SV40 large T antigen was not inactivated at 37°C.
Coculture of the mouse ESCs with the stromal cell lines
To evaluate the ability of the MLSgt cells to induce mouse ESCs to differentiate into mature hepatocyte-like cells, the mouse ESC–derived AFP-producing cells were cocultured on the feeder layer of the MLSgt cells for 7 days. The cells cocultured with the MLSgt cells expressed mature hepatocyte markers, including G6P, TAT, and TO, whereas the cells cultured alone on collagen type I–coated dishes did not express these maturation markers (Fig. 1A). On the other hand, the AFP expression of the cocultured ESCs with the MLSgt cells was weaker than that of the cells cultured on collagen type I. PAS staining showed the mouse ESC–derived piled up colonies to be stained well (Fig. 1B, E). On the other hand, the ESC-derived AFP-producing cells cultured alone on collagen type I–coated dishes were not stained (Fig. 1D, G). These findings indicated that the mouse ESCs were stimulated to differentiate into mature hepatocyte-like cells by the coculture with the MLSgt cells.
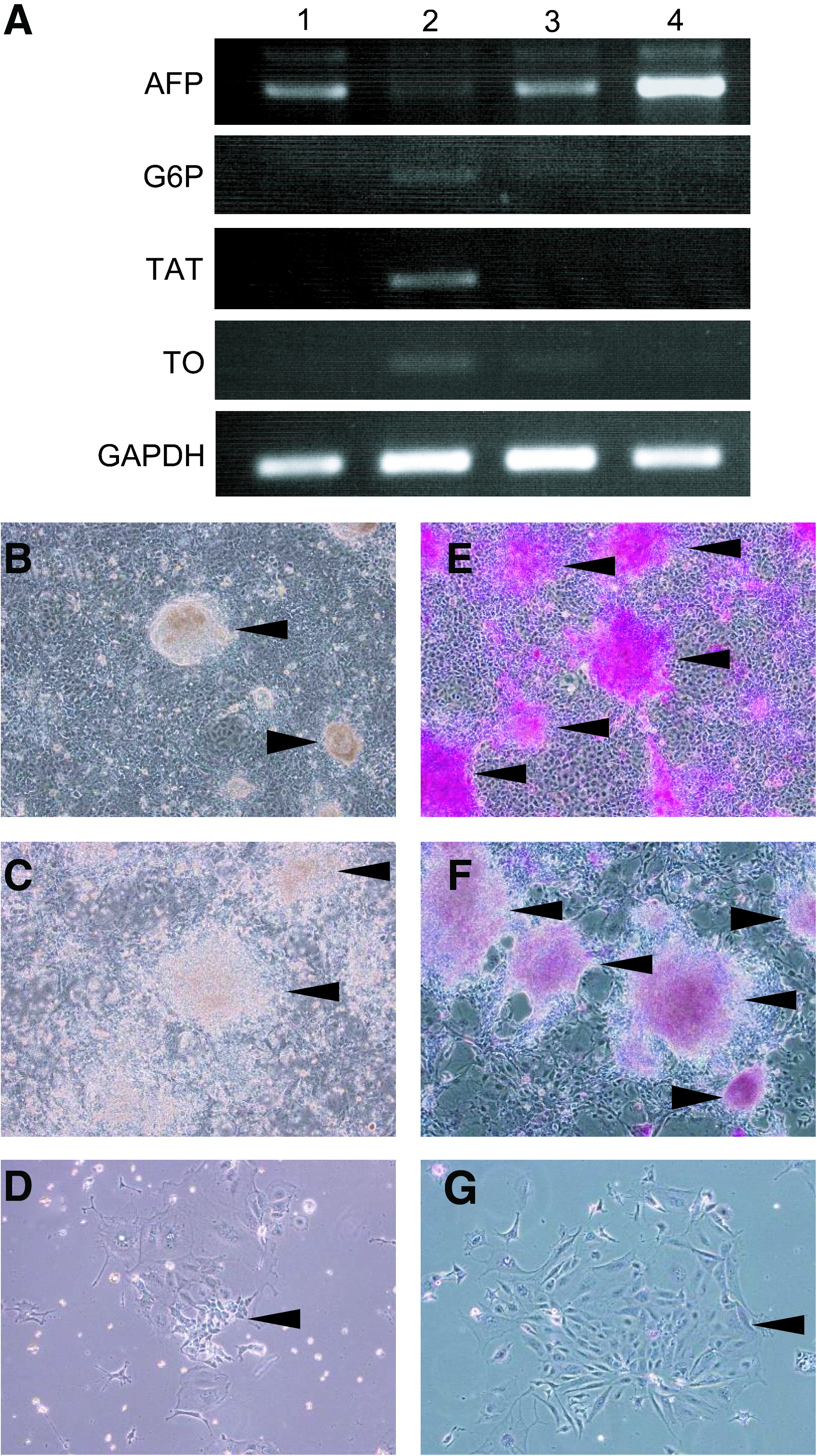
The effect of the MLSgt cells on hepatic maturation. (
To utilize the MLSgt cells as a feeder layer, they were treated by 5 μg/mL mitomycin C for 2 h. However, RT-PCR showed that the mouse ESC–derived AFP-producing cells cocultured with the mitomycin C–treated MLSgt cells did not express the mature hepatocyte markers (Fig. 1A) and that the PAS staining intensity decreased (Fig. 1C, F).
Selection of the most suitable MLSgt cell for hepatic maturation
Because the MLSgt cells were not cloned, they were thought to include various types of cells with different abilities to induce the hepatic maturation. Therefore, to select the most appropriate clone for hepatic maturation, 35 clones were obtained using the limiting dilution method. The ESC-derived AFP-producing cells cocultured with 14 of the 35 clones were hardly stained by PAS stain (data not shown). Therefore, these 14 clones were eliminated from further investigations. The gene expression of the remaining 21 clones was examined by RT-PCR (Fig. 2A). An RT-PCR analysis revealed that no clone expressed G6P, TO, TTR, or AAT. However, 16 of the 21 clones expressed TAT, which is one of the hepatocyte marker genes. To eliminate the possibility of contamination by immortalized HPCs, further examinations were performed using the five clones that did not express TAT: No. 10, 13, 14, 15, and 17 clone. To evaluate the functional properties of the cocultured ESC-derived AFP-producing cells with these five clones, quantitative PCR analyses were performed. AFP was used as an immature hepatoblast marker, and G6P, TO, and TTR were used as mature hepatocyte markers (Fig. 2B–E). The relative expression level was compared with the ESC-derived AFP-producing cells cocultured with the primary mouse CD49f(+/−)CD45(−)Thy1(+)gp38(+) stromal cells. The AFP expression level of cocultured ESCs with No. 10 MLSgt cells decreased to approximately 36% of that of coculture with the primary cells (Fig. 2B). On the other hand, the expression level of G6P was 0.7-fold (Fig. 2C), that of TO showed a 17.6-fold higher expression (Fig. 2D), and that of TTR was 0.13-fold, in comparison with the expression of each of the cocultured cells with the primary cells. The results, therefore, suggested that the No. 10 MLSgt cell was the most appropriate line of five clones for hepatic maturation. This clone was named as MLSgt20.
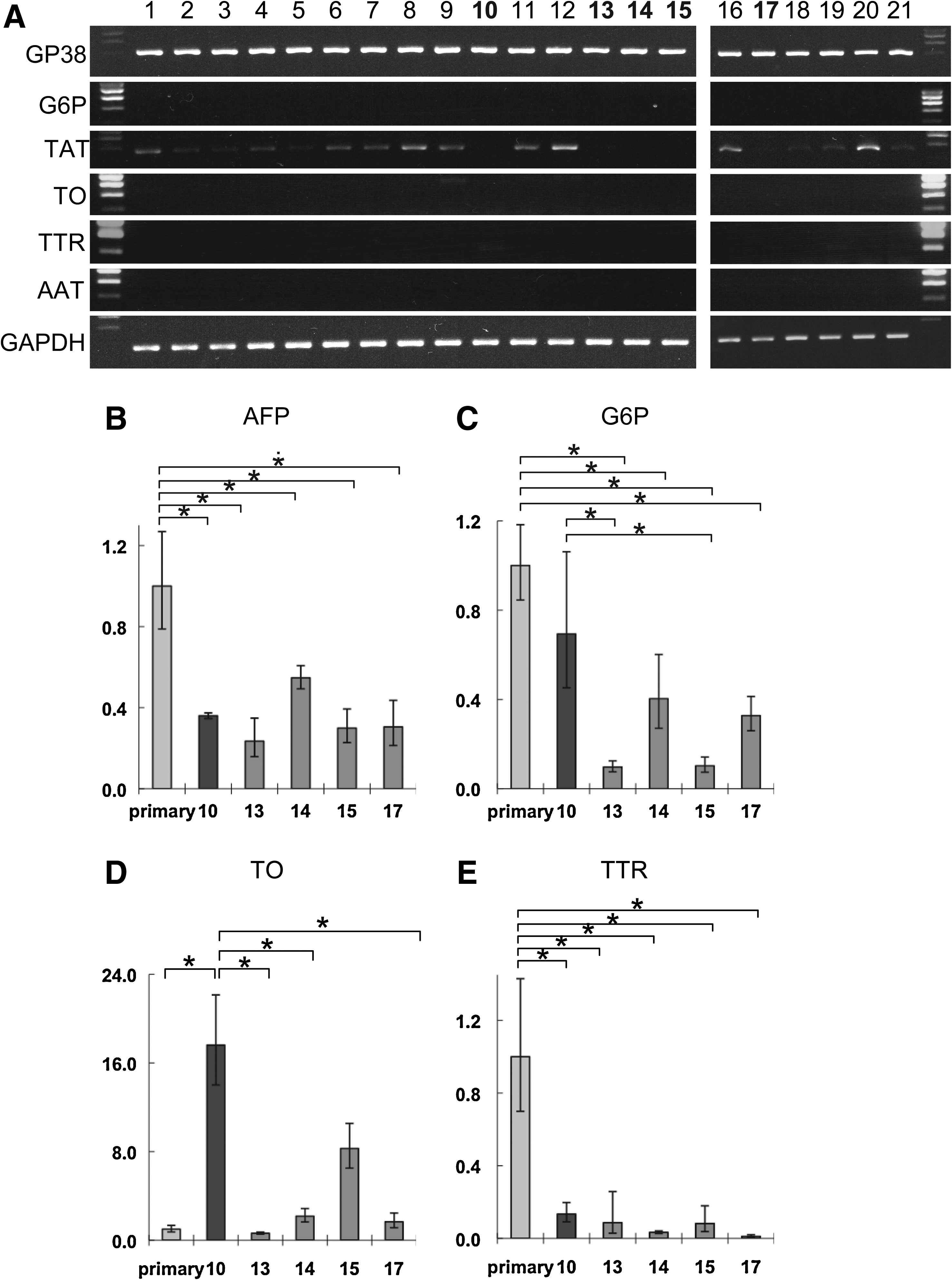
Selection of one MLSgt cell with the most advanced ability to induce hepatic maturation. (
Characterization of the selected MLSgt cell
Phase-contrast microscopy demonstrated that the MLSgt20 cells exhibited cuboidal shapes, which were similar to the original fetal liver stromal cells (date not shown). A flow cytometric analysis showed this cell line to express gp38, but not Thy1 (Fig. 3A). The expression of Thy1 disappeared during the process of immortalization. An RT-PCR analysis was performed to compare the gene expression patterns between the primary gp38-positive stromal cells and MLSgt20 cells (Fig. 3B). Both expressed α-smooth muscle actin, desmin, and vimentin, which are stromal markers. Neither of them expressed an endoderm marker (AFP), a hepatic stellate cell marker (glial fibrillary acidic protein), or endothelial cell markers (platelet endothelial cell adhesion molecule and vascular endothelial cadherin). On the other hand, the expression pattern of two genes was different. The expression of CD34 (one of the stromal markers) disappeared, and CD16 (Kupffer cell marker) was expressed weakly in the MLSgt20 cells.
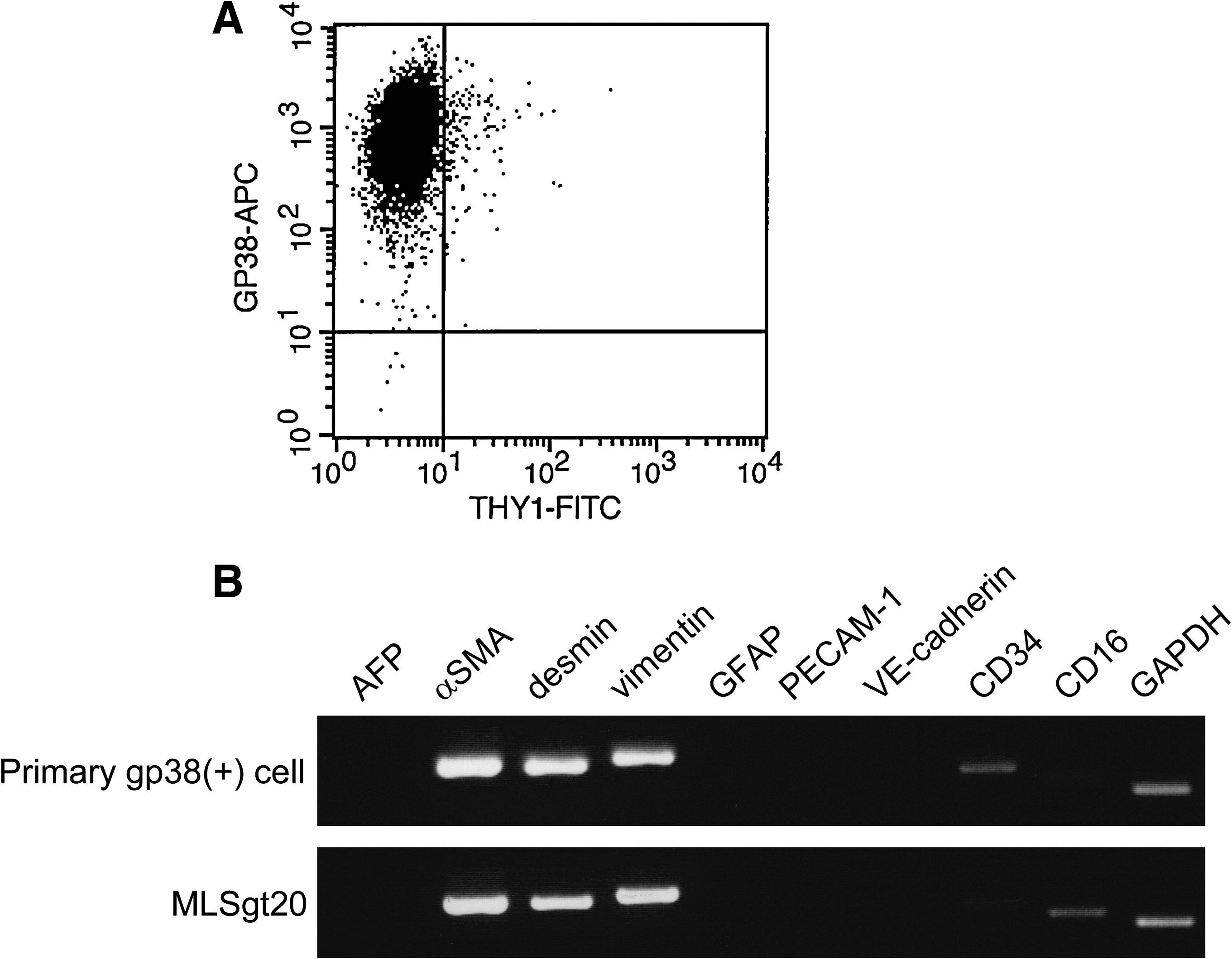
Characterization of the MLSgt20 cells. (
PCR analysis of hepatic gene expression in cells cocultured cells MLSgt20 cell
The ESC-derived AFP-producing cells were cultured under various culture conditions (Fig. 4). The mouse ESC–derived AFP-producing cells were cocultured on a layer of the MLSgt20 cells, cultured alone on collagen type I–coated dishes, on Matrigel-coated dishes, or on an MLSgt20 cell layer fixed by glutaraldehyde. To compare another type of cell line, the ESC-derived AFP-producing cells were cocultured with SL-10. These cells were cultured in two types of medium, fresh HD medium and HD medium conditioned by the MLSgt20 cells. The mouse ESC–derived AFP-producing cells cocultured with the MLSgt20 cells expressed mature hepatocyte genes. Regarding G6P, the expression level was the same as that cocultured with SL-10; however, the expression levels of TAT, TO, and cytochrome P450 3a11(Cyp3a11) were detected when cocultured with MLSgt20. On the other hand, the mouse ESC–derived AFP-producing cells cultured alone, even if they were cultured in the MLSgt20-conditioned media, did not express these genes. These findings suggested that the hepatic maturation required the cell contact with the viable MLSgt20 cells. Humoral factors secreted by the MLSgt cells or cell contacts with other fibroblasts might not be sufficient to stimulate the differentiation of ESCs into hepatocyte-like cells.
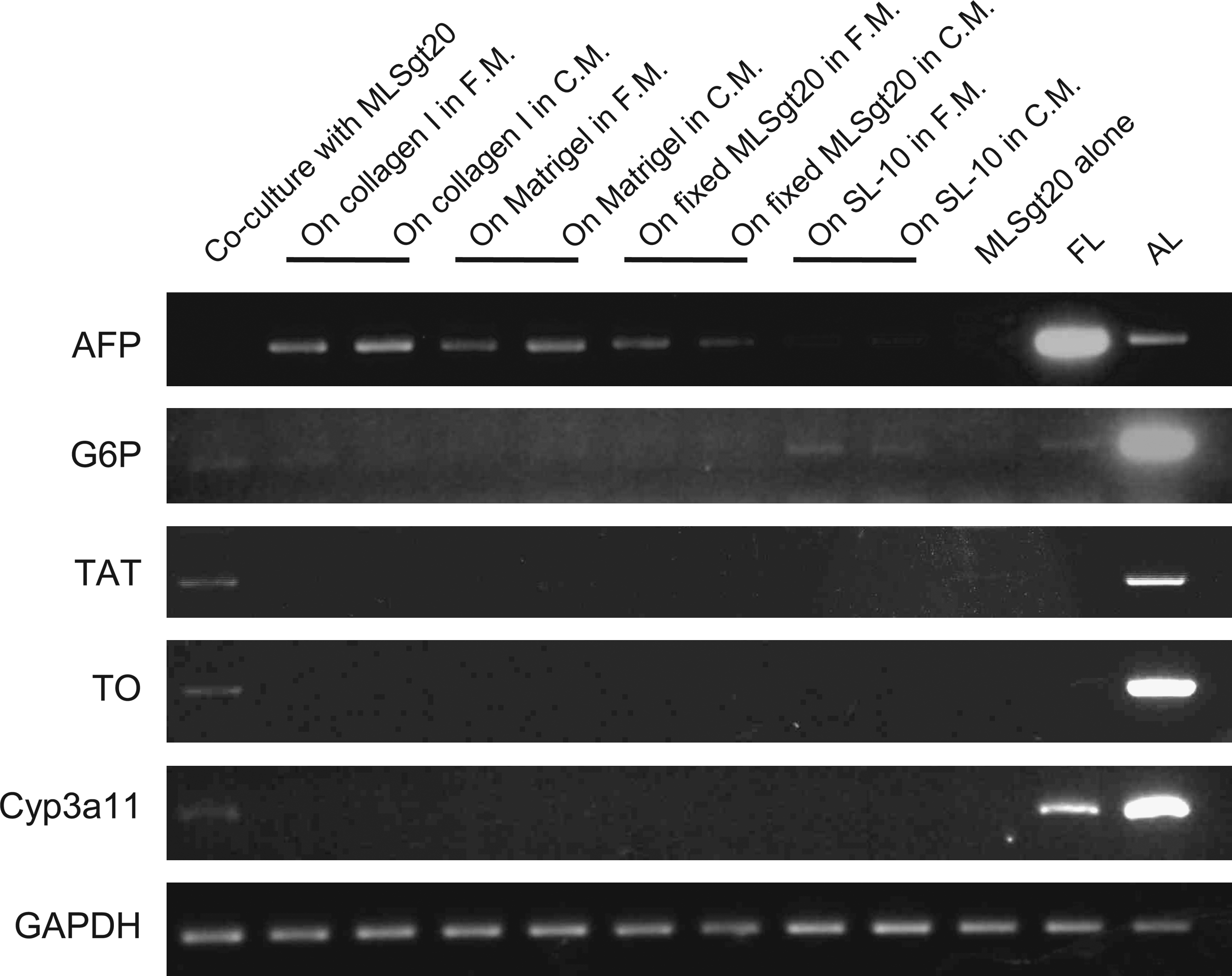
An RT-PCR analysis of cultured cells under various conditions. Mouse ESC–derived AFP-producing cells were cultured under nine conditions. They were cocultured with the MLSgt20, cultured on collagen type I–coated dishes, on Matrigel-coated dishes, on the MLSgt20 cell layer fixed by glutaraldehyde, and cocultured with the SL-10 layer. They were cultured in fresh media (F.M.) or conditioned media from the MLSgt cells (C.M.). FL, E13.5 murine fetal liver; AL, adult liver.
Hepatic maturation of ESCs by coculture with MLSgt20
The ESC-derived AFP-producing cells that were cocultured with MLSgt20 cells were positive for PAS staining in piled up regions (Fig. 5A). The ESC-derived cells formed these piled up colonies surrounded by the feeder layer of the MLSgt20 cells. All cells in the piled up colonies were positive for PAS staining. The HPCs cocultured with MLSgt20 cells were also positive (Fig. 5B) and expressed the mature hepatocyte markers in RT-PCR analysis (Fig. 5C). The ammonia clearance activity is an important hepatocyte function (Fig. 5D). The amount of ammonia removed by the ESC-derived AFP-producing cells alone and the cell line MLSgt20 alone was 244 μM/24 h/well and 255 μM/24 h/well, respectively. The primary hepatocytes removed about 722 μM/24 h/well of ammonia. The cocultured cells removed 459 μM ammonium chloride, which was about twofold greater ability than that of the ESC-derived AFP-producing cells alone (p = 0.01). The concentrations of albumin in culture media were measured after 7 days of culture (Fig. 5D). The cocultured cells and primary hepatocyte secreted albumin 30.7 ng/mL/24 h and 206.3 ng/mL/24 h, respectively. On the other hand, albumin in the culture medium of ESC-derived AFP-producing cells alone or MLSgt20 alone was not detected. To further examine the hepatocyte-specific function, cytochrome P450 detoxification enzymatic assay was performed (Fig. 5E). The cocultured cells showed significantly high Cyp1a1 and Cyp1b1 activities (688.0 ± 37.5 RLU and 718 ± 35.6 RLU, respectively), in comparison to the ESC-derived AFP-producing cells alone (412.3 ± 62.1 RLU and 553 ± 21.2 RLU, respectively) or MLSgt20 alone (4.3 ± 3.1 RLU and 9.0 ± 4.3 RLU, respectively) (p < 0.01). The cocultured cell showed also a higher activity of Cyp1a2 (275.0 ± 51.6 RLU) than the ESC-derived AFP-producing cells alone (190.3 ± 30.1 RLU) or the MLSgt20 cells (65.7 ± 33.5 RLU) did, whereas it did not show a statistically significant difference. We examined these cytochrome activities of primary hepatocytes. They showed about a 14 times greater activity in Cyp1a1 (9751.7 ± 220.4 RLU), 17 times greater Cyp1b1 (12164.0 ± 351.0 RLU), and 2000 times in Cyp1a2 (549430.0 ± 15095.6 RLU) compared to the cocultured cells, respectively. These findings indicated that the ESC-derived endoderm cells cocultured with MLSgt20 cells had cytochrome enzyme activities, even if their activities were weaker than those of the primary hepatocytes. To evaluate the morphological characteristics of the ESC-derived cells cocultured with the MLSgt20 cells, the cocultured cells were observed by transmission electron microscopy (Fig. 5F). The mouse ESC–derived cells cocultured with the MLSgt20 possessed large numbers of glycogen granules and peroxisomes, well-developed rough endoplasmic reticulum, and mitochondria in their cytoplasm. Occasionally, they formed biliary canaliculi–like structures with tight junctions. On the other hand, the ESC-derived AFP-producing cells cultured alone on collagen I–coated dishes did not show these hepatocyte-specific characteristics and did not form any biliary canaliculi either (data not shown).
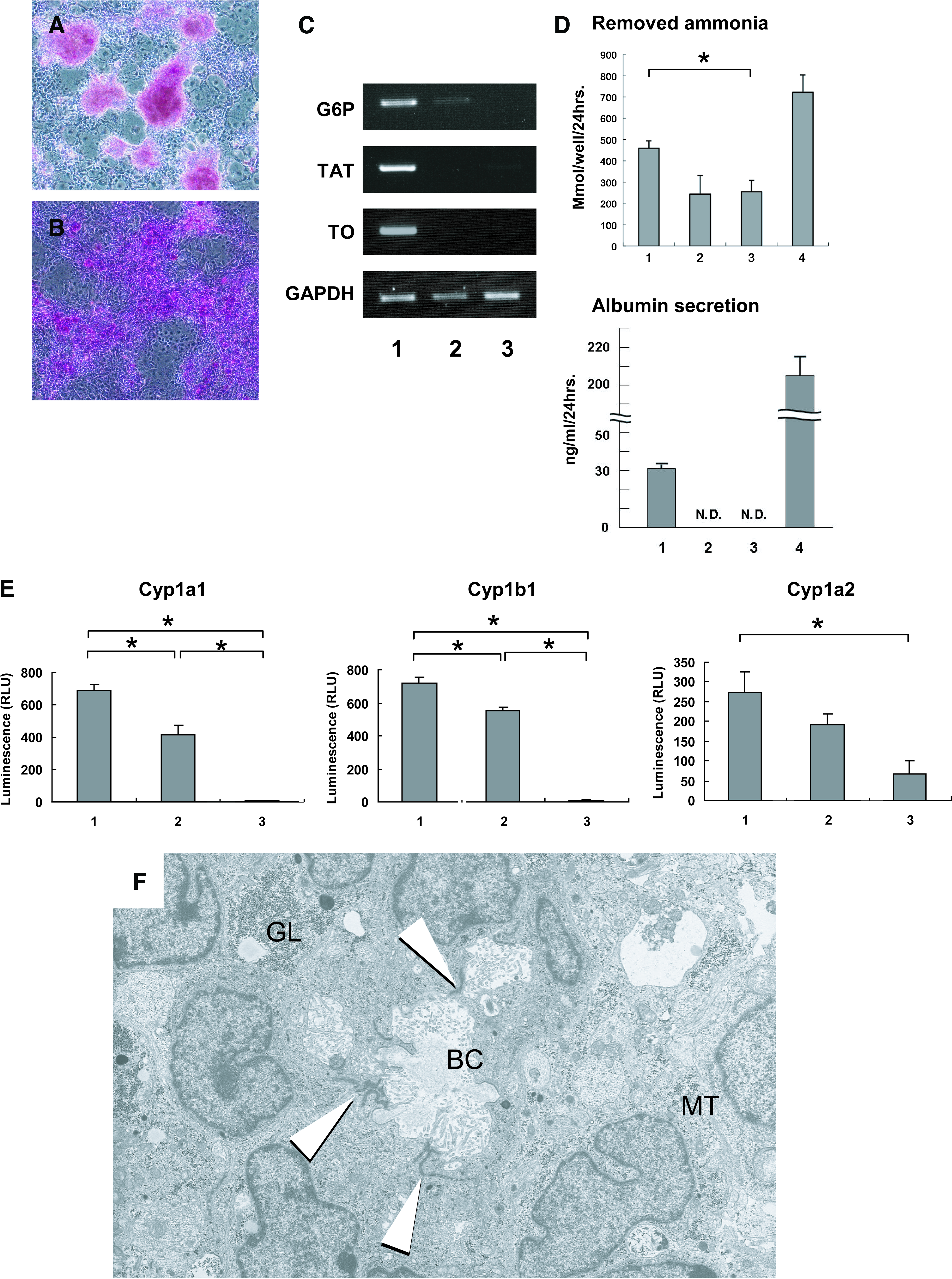
The ESC-derived AFP-producing cells that were cocultured with the MLSgt20 had hepatocytes functions and hepatic morphologies. Periodic acid-Schiff staining of the mouse ESC–derived AFP-producing cells (
Discussion
The functional hepatocytes derived from ESCs may therefore be useful for cell transplantation therapy, bioartificial livers, and drug discovery supporting systems. Although several articles reported that mouse and human ESCs could differentiate into hepatocyte-like cells,12,23,27,28 it has been difficult to differentiate ESCs into functional mature hepatocytes. This is because the process of the hepatic development has not been completely clarified. This laboratory reported that the stromal cells residing in fetal mouse livers have the ability to promote the hepatic maturation of murine ESCs and HPCs by cell–cell contact. 21 This study sought to establish a cell line that had the capability to induce ESCs to differentiate into mature hepatocyte-like cells, based on hepatic developmental biology.
The temperature-sensitive SV40 large T antigen was transfected into the primary CD49f(+/−)CD45(−)Thy1(+)gp38(+) stromal cells from the fetal mouse livers, which were unable to proliferate infinitely. The one clone with the most advanced ability for hepatic maturation, the MLSgt20, was selected from the immortalized cell population. The MLSgt20 was demonstrated to have the stromal features of the parental primary CD49f(+/−)CD45(−)Thy1(+)gp38(+) cells. The ESC-derived AFP-producing cells cocultured with the MLSgt20 cells expressed mature hepatocyte-marker genes, including G6P, TO, AAT, and TTR. The cocultured cells also exhibited the functions and morphologies characteristic of mature hepatocytes, suggesting that the MLSgt20 promoted the ESC differentiation into functional hepatocyte-like cells in vitro. On the other hand, the ESC-derived AFP-producing cells that were cultured alone on collagen type I–coated dishes or cocultured with the other stromal cell line, SL-10, did not exhibit any signs of maturation. Therefore, the hepatic maturation effect was unique to the MLSgt.
Sixteen of the clones expressed the mature hepatocyte gene marker, TAT. There is a possibility that the original stromal cells were contaminated with HPCs. In any case, the selected MLSgt20, which did not express any endodermal markers, was thought to be stromal cells.
The Thy1 expression of the MLSgt20 cells disappeared during the process of immortalization, whereas their parental cells were positive for Thy1. However, the hepatic maturation effect was retained in this cell line. Therefore, Thy1 might simply act as a cell surface marker and not play an important role in hepatic maturation.
The MLSgt20 stimulated hepatic maturation not only on mouse ESCs, but also on mouse HPCs. This maturation process never occurred when these cells were cultured on collagen type I–coated dishes, on Matrigel-coated dishes, on the mitomycinC (MMC)-treated MLSgt cells, or on the MLSgt cell layer fixed by glutaraldehyde, even in the conditioned medium from the MLSgt20. These results suggested that the mechanism of the hepatic maturation by coculture with MLSgt20 was mediated by some specific surface molecules present on the MLSgt20 and/or by very short-acting agents secreted by the MLSgt cells, which was consistent with the previous findings. 22 Liver organogenesis is promoted by cell–cell contact between hepatoblasts and endothelial cells. 29 Although the MLSgt20 cells did not express any endothelial markers, the relationship between the primary CD49f(+/−)CD45(−)Thy1(+)gp38(+) stromal cells and endothelial cells during the hepatic development remains unclear. To elucidate the mechanisms of hepatic maturation, this cell line will be a worthy tool of the investigation.
In this study, the SV40 large T antigen had a temperature-sensitive element. Therefore, the immortalized cells had been expected to grow at 33°C and to cease proliferating at 37°C. However, they continued to proliferate even at 37°C. To apply the coculture technique to cell transplantation, it would be necessary to deplete the MLSgt cells. Therefore, additional procedures such as transfection with a suicide gene, such as herpes simplex virus-thymidine kinase gene, may be necessary.
In conclusion, a cell line was established that possessed the capability to induce mouse ESCs and HPCs to differentiate into mature hepatocyte-like cells, and these findings were based on knowledge of the process of hepatic developmental biology. A large number of the ESC-derived mature hepatocyte-like cells were obtained easily using this cell line. These findings will contribute to the availability of ESC-derived hepatocytes with sufficient capabilities to be applied for practical use in cell therapy, bioartificial liver, and drug discovery research and additionally may apply to the differentiation of both human ESCs and induced pluripotent stem cells into functional hepatocytes.
Footnotes
Acknowledgments
We thank Makio Fujioka for his valuable assistance with transmission electron microscopy. This study was supported in part by grants from the Scientific Research Fund of Ministry of Education, Culture, Sports, Science, and Technology of Japan and by the New Energy and Industrial Technology Department Organization.
Disclosure Statement
No competing financial interests exist.