
Abstract
Fibrin gels are attractive scaffolds useful for neural tissue engineering applications. The objective of this work was to investigate the apoptotic activity, survival, proliferation, and differentiation of a mixed population of primary neural cells composed of neurons and multipotent precursor cells when cultured in fibrin gels prepared with varying concentrations of fibrinogen (5–25 mg/mL fibrinogen) and thrombin (1–125 U/mL thrombin). Within all fibrin gel formulations tested, the level of apoptosis on day 1 was low and cell survival was equivalent to levels in monolayer culture (67%). Proliferation in gels made from 5 to 12.5 mg/mL fibrinogen was also similar to that observed in monolayer culture, though a lower proliferative response was observed in 25 mg/mL fibrinogen formulations. Relative to monolayer culture, cholinergic and dopaminergic neuronal presence was enhanced, whereas glial cell growth was reduced in fibrin gel cultures. The extent to which levels were altered depended on fibrinogen and thrombin concentration. The findings here suggest the importance of fibrinogen and thrombin concentration in differentially regulating the growth and composition of neural cell populations and are of importance for neural tissue engineering strategies focused on the development of implantable scaffolds for treating neurodegenerative disorders.
Introduction
Toward this end, the focus of this work is to characterize the growth and differentiation of fetal rat cortical cells in fibrin gels prepared with different concentrations of fibrinogen and thrombin to facilitate the development of a fibrin-based tissue engineering therapeutic capable of improving cell transplantation therapy for the treatment of neurodegenerative diseases. The fibrin used in this study is a commercially available sealant called Tisseel®, a material that has been shown to be nonimmunogenic when injected into primate brains. 6 The fetal cells used in this study are isolated at a developmental stage (E14–E15) that closely correlates with the human tissue (16–20 weeks) that is typically used for human transplantation procedures. 7 This rat cell population has demonstrated curative potential in animal models of neurodegenerative disorders.8–10 The fetal tissue is composed of a mixed population of neurons and multipotent neural precursor cells (NPCs). Fibrin has previously been shown to be a permissive matrix for neurons and is capable of supporting the repair of peripheral and central nerve injuries,11–14 but the behavior of undifferentiated NPC populations in fibrin gel cultures is less widely studied. Fetal-derived NPCs are of considerable interest because they have been shown to integrate well with host tissue upon implantation within the brain, and they have the potential to differentiate into neuronal phenotypes that die during the progression of Alzheimer's (cholinergic neurons) or Parkinson's (dopaminergic neurons) disease, though they can also differentiate into astrocytes that may contribute to glial scar formation. 15 Although purer sources of NPCs such as immortalized clones are now available, their curative potential is still not concretely established. 15
Identification of 3D culture conditions that can maximize the growth of key neuronal phenotypes from dissociated fetal tissue while minimizing the growth of astrocytes is a priority for the field. Mixed cortical cultures have been used previously in two-dimensional fibrin culture to help elucidate matrix properties that select for neuronal over glial cell growth, 16 and expanding this type of study to three dimensions is useful for neural tissue engineering applications. There is evidence to suggest that 3D fibrin gels would be supportive of neuronal differentiation as demonstrated in recent publications involving murine embryonic stem-cell-derived NPCs.17,18 Here we build on these promising findings and comprehensively explore the influence of fibrinogen and thrombin concentration on the growth and composition of fetal cells in three dimensions. Studies were designed to characterize the effect of fibrinogen and thrombin concentration on gel properties (pore size, compressive modulus, and degradation rate) and on cellular behavior (apoptotic activity, survival, proliferation, and cell composition) using a combination of biochemical assays and confocal microscopy. To identify potential advantages of culturing this cell population in a fibrin gel, cell behavior in gels was also directly compared to that in monolayer culture.
Materials and Methods
Neural cell isolation and culture
Neural tissue used for all experiments was isolated from E14–E15 fetal rat (Charles River) forebrains and dissociated enzymatically as previously described. 19 The single-cell suspension was encapsulated into fibrin gels or plated as monolayer cultures as described in the section titled “Preparation of cell-seeded fibrin gels and monolayer cultures,” and then cultured at 37°C and 5% CO2 in serum-free media consisting of 50:50 Dulbecco's modified Eagle's medium:F12 (MediaTech, Manassas, VA) supplemented with 1 × N2 (Invitrogen, Carlsbad, CA), 100 U/mL penicillin–streptomycin (Hyclone, Logan, UT), 1 mM L-glutamine (Invitrogen), and 10 ng/mL basic fibroblast growth factor (Sigma, St. Louis, MO). The growth medium was changed every other day for 1–7 days depending on the experiment.
Preparation of cell-seeded fibrin gels and monolayer cultures
Fibrin gel culture
Cells were encapsulated within fibrin gels by mixing two solutions in a 96-well plate, (1) 50 μL of a fibrinogen solution and (2) 50 μL of a thrombin-cell solution (Tisseel; Baxter Biosciences, Deerfield, IL), where the concentration of each component was adjusted using 1 × tris buffered saline (TBS) and 30 mM CaCl2, respectively, to achieve the final gel concentrations listed in Table 1. Cells were suspended in the thrombin solution to avoid spontaneous polymerization of the fibrinogen solution caused by the endogenous secretion of thrombin by neural cells. 20 Upon mixing, the fibrinogen and thrombin coagulated to form a fibrin network that encapsulated the cells at a final concentration of 1 × 107 cells/mL. Polymerization was allowed to progress for 1 h in the incubator. One hundred microliters of the medium was then added, and gels were incubated for long-term culture.
Monolayer culture
Twenty-four-well plates were coated overnight in phosphate-buffered saline containing 4 μg/mL poly(ornithine) (Sigma). Wells were rinsed, and cells were seeded at a final cell density of 2 × 105 cells/cm2.
Estimation of pore diameter and compressive modulus of fibrin gels
Pore diameter
Fibrin gels were made as described above (see section titled “Preparation of cell-seeded fibrin gels and monolayer cultures”) except without cells and with the additional incorporation of fibrinogen Alexafluor-488 (Invitrogen; 50:1 unconjugated:conjugated fibrinogen). Fibrin gels were prepared on microscope slides to allow for direct imaging of the network structure. Freshly prepared fibrin gels were imaged using the 40 × water immersion objective of a confocal microscope (Zeiss, Oberkochen, Germany) with a 280 nm lateral resolution limit and a 1.2 μm axial resolution limit. The average pore diameter was estimated by manually measuring n = 100 pores present in three different images acquired from three different gels using NIH ImageJ software (Table 1).
Compressive modulus
The compressive modulus of n = 9 fibrin gels from three different experiments was determined 24 h after fabrication as previously described. 21 Modulus was assessed after at least 24 h had elapsed based on previous reports that the stiffness of fibrin gels can take up to 24 h to reach a maximum. 22 The compressive modulus was measured for only one thrombin concentration (1 U/mL) per fibrinogen concentration based on previous reports showing that increasing thrombin concentrations in the range tested in this study has a minimal effect on compressive modulus.23,24
Quantification of gel degradation
The rate of cell-mediated fibrin gel degradation was quantified as previously described. 18 Briefly, fibrin gels were made as described above (see section titled “Preparation of cell-seeded fibrin gels and monolayer cultures”) except with the additional incorporation of fibrinogen Alexafluor-488 (see section titled “Estimation of pore diameter and compressive modulus of fibrin gels”). The total amount of fluorescently labeled degradation product released into the culture medium was quantified using a spectrofluorometer (Fluostar, Alexandria, VA). A linear standard curve is obtainable as long as fibrinogen concentrations greater than 0.025 mg/mL are being assessed. Twelve fibrin gels from two different experiments were assessed. The extent of gel degradation is expressed as the total amount of fluorescent degradation product released over a given time interval (1, 5, or 9 days) divided by the total amount of fluorescent material initially present in the gel as estimated by direct measurement of freshly made fibrin gels enzymatically digested using 10 × trypsin. Control gels were also prepared in the absence of cells.
Measurement of total DNA, ATP, and caspase activity in fibrin cultures
On the day of harvest, fibrin gel cultures were enzymatically degraded using 100 μL of collagenase (25 U/mL) (Sigma) to dissolve the fibrin gels, and then 100 μL of cell lysis buffer (20 mM Tris [Bio-Rad, Hercules, CA], 2 mM EDTA [Bio-Rad], 150 mM NaCl [Sigma], and 0.5% Triton X-100 [Fisher Biotech, Wembley, Australia] in diH2O) was added to lyse the cells. All assays were carried out according to the manufacturer's instructions. Total DNA, ATP, and caspase levels were quantified via the PicoGreen assay (Invitrogen), the Cell Glo Assay (Promega, Madison, WI), and the Apo-One assay (Promega), respectively. When measuring caspase levels, collagenase was not used to avoid unintentional cleavage of the assay's fluorogenic substrate. Instead, cell lysis buffer was directly applied to intact fibrin gels to lyse cells. Data represent the average values of n = 16 samples collected from four different experiments. As a negative control and to verify assay functionality, an additional polyethylene glycol (PEG) hydrogel culture condition also accompanied each ATP and caspase assay. Some degree of apoptosis is commonly observed during the first 24 h when this cell population is encapsulated within a PEG gel. (For details on PEG hydrogel culture, see Mahoney and Anseth. 25 )
Assessment of cell composition within fibrin gels
Immunocytochemistry
On days 0, 1, and 7, fibrin gel and monolayer samples were fixed in 4% paraformaldehyde (Sigma). Fibrin gels were cryosectioned into 40-mm-thick sections and mounted on slides. Using standard immunocytochemical techniques, samples were processed for staining with antibodies against nestin, β-tubulin, and glial fibrillary acidic protein (GFAP). These cell markers are commonly used for characterization of E14–E15 brain tissue, 26 though later in development GFAP and nestin are known to begin to co-label a population of radial glial cells. 27 Antibodies were obtained from Sigma. Secondary antibodies conjugated to Alexafluor probes were obtained from Invitrogen. 4′,6-diamidino-2-phenylindole (DAPI) was used as a nuclear counter stain. Samples were imaged via confocal microscopy using the 40 × objective, and the percentage of positive nuclei was manually determined in n = 15 images/condition/cell type collected from three different experiments.
Quantitative real-time reverse transcriptase-polymerase chain reaction
Standard quantitative real-time reverse transcriptase-polymerase chain reaction (qRT-PCR) was performed to assess the relative expression level of cell-type-specific genes on days 0 and 7 of culture in n = 12 samples obtained from three different experiments. Primers were developed to detect gene expression levels for nestin, β-tubulin, choline acetyltransferase (ChAT), tyrosine hydroxylase (TH), and GFAP. Primer sequences listed in Table 2 were obtained from Invitrogen. Isolated RNA was DNAse treated and then accurately quantified using RiboGreen Quantification Reagent (Invitrogen). Equal amounts of RNA (100 ng) were placed in each reverse transcription reaction, thus alleviating the need for inclusion of house-keeping genes. 28 cDNA synthesis and SYBRgreen PCR was carried out per the manufacturer's recommendation (Applied Biosystems, Foster City, CA).
NPC, neural precursor cell; TH, tyrosine hydroxylase; ChAT, choline acetyltransferase; GFAP, glial fibrillary acidic protein.
Characterization of cell distribution and morphology within fibrin gels
Cell morphology within the fibrin gels was observed via confocal microscopy after labeling live cells on day 7 of culture with calcein-AM (Invitrogen). Each analyzed image acquired using a 10 × water immersion objective is a projection of 10 optical slices acquired along the z-axis to reflect a 100-μm-thick portion of the fibrin gel.
Statistical analysis
For all data, statistical significance was determined using a two-tailed Student's t-test. p < 0.05 is considered significant and is denoted with an asterisk unless otherwise stated. Data are presented as mean ± standard error of the mean.
Results
Fibrin gel structure
To study how fibrinogen and thrombin concentrations affect the structure of the Tisseel fibrin network, fibrin gel formulations listed in Table 1 were prepared and observed using confocal microscopy. Results demonstrate that the pore size decreases as fibrinogen concentration increases (Table 1). As thrombin concentration increases from 1 to 125 U/mL, the average pore size decreases approximately by half (Table 1). Representative images enable visual confirmation that a tighter overall network results when either fibrinogen (Fig. 1a vs. 1b) or thrombin (Fig. 1b vs. 1c) concentrations are increased.

Structure and degradation rate of fibrin gels as fibrinogen and thrombin concentration varies. (
Compressive modulus of fibrin gels
Varying the fibrinogen concentration also influenced the compressive modulus of fibrin gels. The compressive modulus is an indicator of the mechanical stiffness of the network, a property that is important to consider as it has been shown to influence process extension from neurons 29 and glial cell growth. 16 Our results show that when fibrin gels were made with fibrinogen concentrations that increase from 5 to 25 mg/mL, the compressive modulus of the gels increased approximately threefold (Table 1).
Cell-mediated degradation of fibrin gels
To determine the extent of cell-mediated fibrin gel degradation over the course of the culture period, fibrin gels were made with the additional incorporation of fluorescent fibrinogen. The release of fluorescently labeled degradation products was monitored during the culture period. Results showed that fibrin networks prepared with higher fibrinogen concentrations degraded more slowly (Fig. 1d). Fibrin networks prepared with higher thrombin concentrations degraded more quickly (Fig. 1d). All gels were present for the duration of the culture period (7 days), a timescale slightly shorter than the timescale over which the fastest degrading gel (5 mg/mL fibrinogen and 125 U/mL thrombin) degraded (9 days).
Influence of fibrin formulation on cell viability: Apoptosis and metabolic activity measurements
In effort to determine if fibrin formulation impacts cell viability, apoptotic activity was assessed on days 0, 1, and 7 of culture as measured by an increase in basal activities of caspase-3 and -7. To account for differences in cell number, caspase activity levels were normalized to total DNA content. On day 0, caspase activity normalized to DNA content was the same across all formulations (4.6 ± 0.3 U caspase/mg DNA) and was not statistically different from levels in monolayer culture (4.1 ± 0.5 U caspase/mg DNA) (data not shown). In all of the fibrin gel formulations and monolayer culture, caspase activity on day 1 or on day 7 was not significantly different from levels on day 0 (Fig. 2a). In contrast, when cells are cultured within a negative control synthetic PEG hydrogel, an increase in caspase activity is observed over the first day as many cells die when photo-encapsulated into the synthetic environment before recovery can ensue.
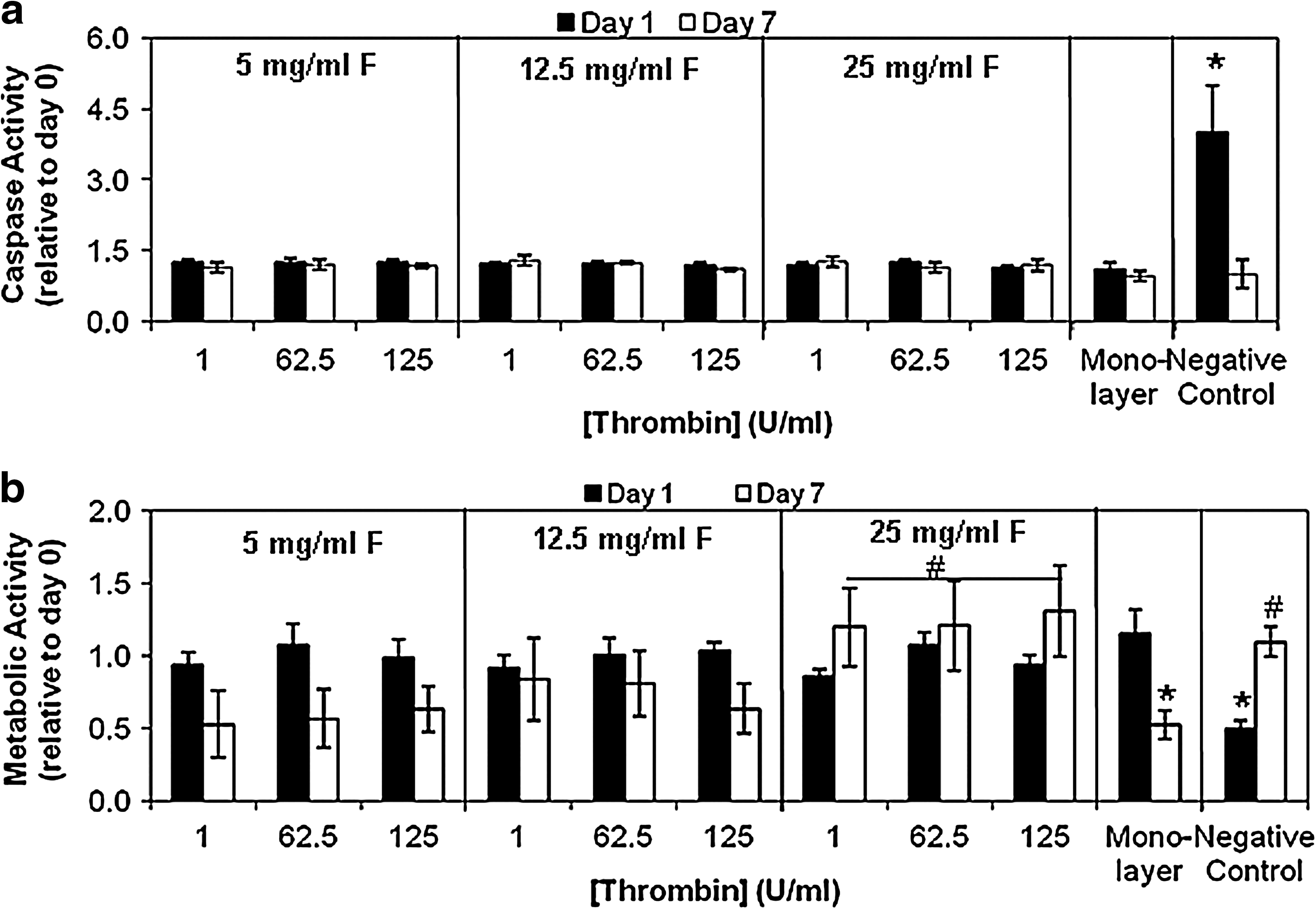
Influence of fibrin formulation on cell apoptosis and viability. (
As another measure to determine if fibrin formulation impacts cell viability, total ATP content normalized with respect to total DNA content was utilized as a quantitative indicator of cell metabolic activity over time. For frame of reference, monolayer cultured cells that are 97% viable (as determined by trypan blue exclusion) contain 201 ± 20 pg of ATP/pg of DNA as previously reported. 25 On day 0 of culture, ATP content normalized to DNA content was the same across all formulations (138 ± 9 pg of ATP/pg of DNA) and was not statistically different from levels in monolayer culture (143 ± 8, ∼67% viability) (data not shown). The change in cell metabolic activity across time is expressed as the ratio of cell metabolic activity on day 1 or 7 to levels on day 0 (Fig. 2b). Results showed that cell metabolic activity on day 1 in either fibrin gel culture or in monolayer culture was not statistically different from that on day 0. Cell metabolic activity on day 7 in fibrin gel culture was not statistically different from that on day 0; however, a significant decrease in metabolic activity was observed in monolayer culture by day 7 (Fig. 2b). Although metabolic activity tends to decrease in fibrin formulations made with lower concentrations of fibrinogen, the decrease is not statistically significant (see Table 3 for p-values). Together, these findings suggest that initial and longer-term cell viability is high in fibrin gel cultures independent of formulation; though by day 7, there is a decrease in the metabolic activity of cells cultured in monolayer culture and, to a lesser degree, in fibrin gels with low fibrinogen concentrations.
Influence of fibrin formulation on cell proliferation and morphology
To determine if fibrin formulation impacts neural cell proliferation, the increase in total DNA content relative to day 0 was measured over time in culture. Between day 0 and 1, there was no significant increase in total DNA content in monolayer culture or any fibrin gel condition (Fig. 3a) (p > 0.05 in all conditions). By day 4 of culture, DNA levels significantly increased in monolayer culture and a similar increase was observed in formulations prepared with either 5 or 12.5 mg/mL fibrinogen. However, cells in fibrin gels made with 25 mg/mL fibrinogen did not proliferate (Fig. 3a). By day 7 of culture, DNA content continued to increase in monolayer culture at a rate only matched by fibrin formulations made with 5 mg/mL fibrinogen and either 62.5 or 125 U/mL thrombin, or those made with 12.5 mg/mL fibrinogen and 125 U/mL thrombin (Fig. 3a). These findings indicate that cells do proliferate well in fibrin gel cultures, although at a slightly reduced level relative to monolayer culture in some formulations—especially fibrin gels prepared with 25 mg/mL fibrinogen.

Influence of fibrin formulation on cell proliferation and morphology. (
Confocal microscopy was used to directly observe the morphology of neural cells encapsulated within the nine fibrin formulations and monolayer culture. Representative images demonstrate that cell morphology was differentially affected among different fibrin formulations (Fig. 3b–d). Although monolayer culture and fibrin gels prepared with 5 and 12.5 mg/mL fibrinogen allowed for extensive cell elongation and process extension (Fig. 3c–e), fibrin gels prepared with 25 mg/mL fibrinogen resulted in the formation of rounded cell clusters (Fig. 3d). Together, these findings indicate that formulations that are least permissive for cell proliferation (25 mg/mL) are also least permissive for cell elongation.
Influence of fibrin formulation on neural cell differentiation
The composition of the freshly isolated cells was consistently determined via immunocytochemistry to be a mixture of multipotent NPCs (50 ± 3% nestin positive) (Fig. 4a, b) and postmitotic neurons (57 ± 4% β-tubulin positive) (Fig. 5a, b). No glial cells are present upon isolation from the E15 rat embryo (0% GFAP positive) (Fig. 6a, b). Immunocytochemistry and qRT-PCR were used to monitor changes in this initial cell population during culture within fibrin gels and monolayer conditions. Due to the high-throughput nature of qRT-PCR, expression of cell-type-specific mRNA transcripts was assessed in all nine fibrin formulations. For immunology studies, cell composition was monitored only in four select fibrin formulations chosen to encompass the highest and lowest fibrinogen and thrombin concentrations.
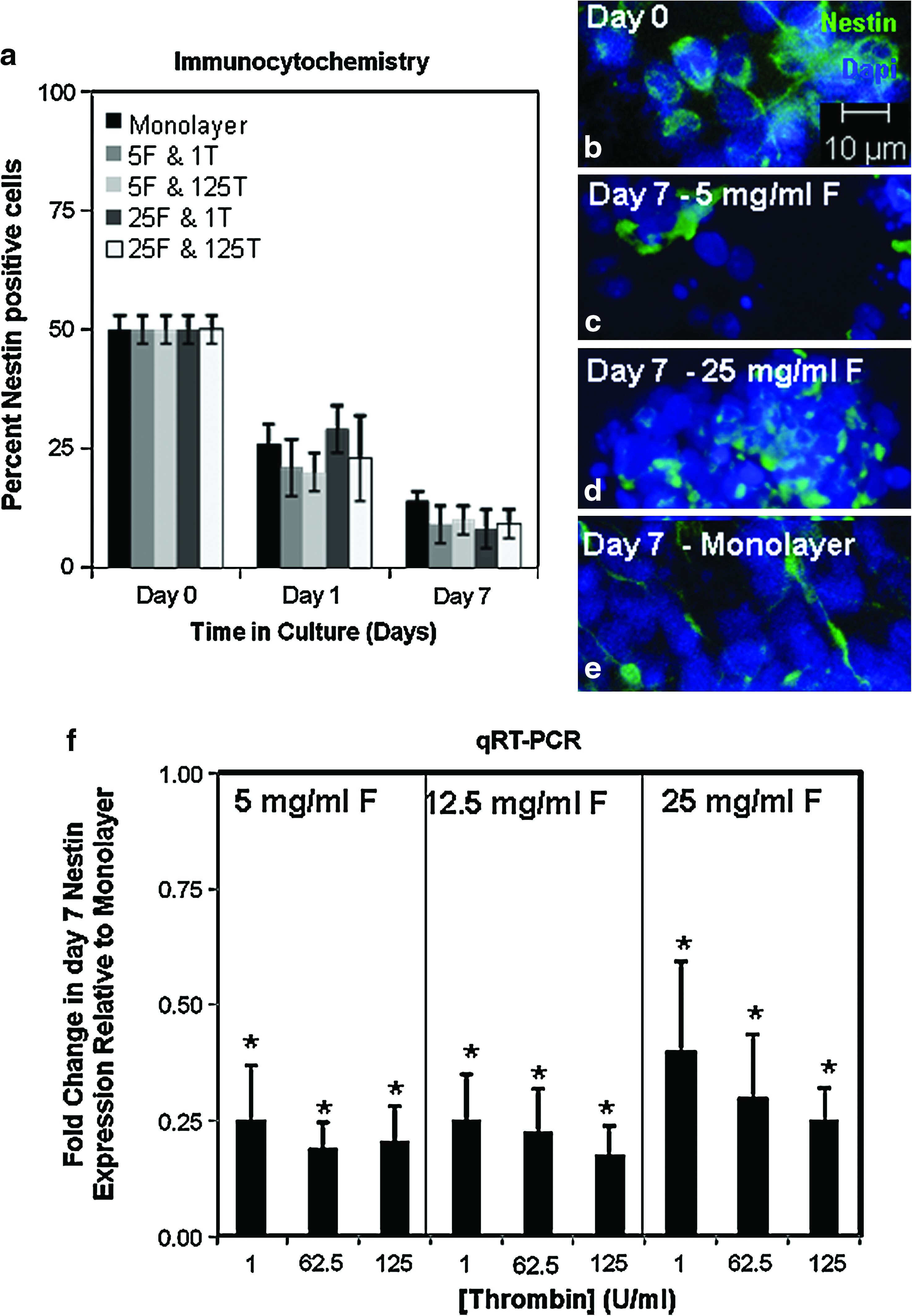
Influence of fibrin formulation on the presence of neural precursor cells. (

Influence of fibrin formulation on the presence of neurons. (

Influence of fibrin formulation on the presence of glial cells. (
Results indicate that when cells are cultured in monolayer and any of the four tested fibrin formulations, the percentage of nestin-positive NPCs that are present upon tissue isolation began to decrease to ∼25% on day 1 and to ∼12% on day 7 (Fig. 4a). qRT-PCR results confirmed a decrease in nestin mRNA expression levels when cells were cultured in monolayer culture, as day 7 nestin levels were only 20% of day 0 levels (data not shown). In all fibrin formulations, an even larger decrease in nestin gene expression was observed (Fig. 4f).
Nestin expression decreases over time presumably as the NPCs differentiate into more mature progeny including neurons and glial cells. Immunocytochemistry was performed to assess expression of neuronal-lineage-specific genes, where β-tubulin expression is pan-neuronal marker for immature neurons. The neurons identified by β-tubulin may have been present upon dissociation or may be newly differentiated progeny. Results demonstrate that the percentage of β-tubulin-positive neurons present in monolayer culture does not significantly decrease over the 7 day culture period (Fig. 5a). Likewise, the growth of neurons is well supported in fibrin gel cultures independent of fibrin formulation as evidenced by the robust staining evident in fibrin gels on day 7 (Fig. 5c, d), though the percentage of β-tubulin-positive cells present on day 7 does decrease to ∼30% of day 0 levels (Fig. 5a). qRT-PCR results confirmed a supportive environment for neurons as β-tubulin mRNA expression levels remain at 80% of day 0 levels when cells were cultured in monolayer culture (data not shown). qRT-PCR results also confirm a significantly decreased level of β-tubulin expression when cells are cultured in fibrin gels relative to monolayer culture independent of the fibrin formulation (Fig. 5f).
As β-tubulin is expressed predominantly in immature neurons, the consistent decrease in β-tubulin-positive cells and mRNA expression observed in culture over the 7 day culture period may be the result of the progression of neurons to a more mature phenotype. To test this possibility, microtubule-associated protein-2 (MAP-2) expression was also assessed as a pan-neuronal marker expressed in mature neurons. Results confirm an increase in the percentage of MAP-2-positive cells over the duration of the culture period (Fig. 5f). Although there was no significant difference in the percentage of MAP-2-positive cells present in monolayer culture and those present in any of the tested fibrin formulations, representative images demonstrate more intense staining in the fibrin gel cultures (Fig. 5h, i) when compared with monolayer culture (Fig. 5j).
The neurons present within fibrin gels may undergo maturation and more quickly transition into specific neurotransmitter-expressing neuronal phenotypes that contain lower β-tubulin levels. To investigate this possibility, the gene expression levels of two neurotransmitter-specific enzymes (ChAT-cholinergic neurons and TH-dopaminergic neurons) were assessed. Results demonstrate that in all fibrin gels tested, ChAT (Fig. 5k, white bars) and TH (Fig. 5k, gray bars) gene expression levels were increased relative to monolayer culture, with this increase dependent on the fibrin gel composition. ChAT gene expression decreased with increasing fibrinogen and thrombin concentration, whereas TH gene expression increased with increasing fibrinogen and thrombin concentration. The differential effect of fibrin formulation on expression of neurotransmitter-specific enzymes was further assessed by confirming that TH expression in fibrin gels made from 25 mg/mL fibrinogen (Fig. 5m) is indeed more intense than TH expression observed on isolated cells present within fibrin gels made from 5 mg/mL fibrinogen (Fig. 5l).
The influence of fibrin formulation on glial differentiation and growth was also determined. In monolayer culture, the first evidence of cells containing brightly labeled GFAP-positive processes did not occur until day 7 when only 1% of the cells stained positively (Fig. 6a, e). GFAP-positive cells never appeared in fibrin gel cultures of any formulation within the 7 day culture period; however, there was an increasing presence of a diffuse, nonspecific stain in the fibrin gels made with 25 mg/mL fibrinogen (Fig. 6a, d) that was absent from the gels made with 5 mg/mL fibrinogen (Fig. 6a, c). qRT-PCR results confirm the increasing presence of GFAP within the cultures, as day 7 GFAP levels were 10,000-fold greater than day 0 levels (data not shown). Interestingly, when cells were cultured within fibrin gels, day 7 GFAP gene expression was significantly reduced relative to levels observed in monolayer culture, though the magnitude of this reduction depended on the fibrin gel composition (Fig. 6). GFAP gene expression was most reduced in formulations prepared with 5 or 12.5 mg/mL fibrinogen, and significantly less reduced when prepared with 25 mg/mL fibrinogen (Fig. 6).
Discussion
This study first characterized the effect that various fibrinogen and thrombin concentrations have on Tisseel fibrin gel stiffness, structure, and degradation rate. Tisseel contains unique salt concentrations and aprotinin that may alter the fibrin material properties relative to other studies. 30 Compressive modulus results were consistent with previous findings in that as fibrinogen concentration increases, the modulus of the resulting gels increases.31–33 Our structural results are also consistent with previous findings in that the porosity of the gel matrix decreases with increasing fibrinogen and thrombin concentrations. 34 Our degradation results demonstrate rapid cell-mediated degradation of fibrin gels prepared using high thrombin concentrations (62.5 or 125 U/mL). Although thrombin concentration greatly influences the degree of degradation when making gels using both high (25 mg/mL) or low (5 mg/mL) fibrinogen, fibrinogen concentration is also a major determining factor in fibrin degradation rates, where formulations with lower fibrinogen concentrations are degraded most quickly by neural cells. These results are consistent with studies showing that fibrin gels made using low fibrinogen and high thrombin concentrations contain thin fibers and that thin fibers are lysed faster than thick fibers.35–37 Interestingly, the degradation timescales observed in the present study are slightly prolonged relative to those achieved in studies performed using purified human fibrinogen, validating the need for a separate study exploring the degradation of Tissel fibrin gels. 38 It is pertinent to note the presence of a recent study that reports results contradictory to those reported here in that increased neurite outgrowth was observed in fibrin gels that contained greater fibrinogen concentrations. 39 By our measurements, greater fibrinogen concentrations create a gel that is less susceptible to proteolysis and should intuitively inhibit neurite outgrowth. This apparent discrepancy may be attributable to the very low thrombin concentrations employed in the previous study (0.25 U/mL), which, we have demonstrated in our lab, leads to very inhomogeneous fibrin gel structures (data not shown) that contain structural attributes (fiber size, fibril bundle diameter, etc.) that are difficult to control and that can substantially influence neurite outgrowth.
Differences in fibrin material properties may variably impact cell function. For example, cells encapsulated within fibrin scaffolds that are very susceptible to proteolytic degradation remain as single cells and extend processes, but when encapsulated within fibrin scaffolds resistant to proteolytic degradation, cells form rounded cell clusters rather than extend processes. This result is consistent with previous findings showing that other cell types remain rounded when gels are prepared with higher fibrinogen, whereas they elongate in gels prepared with lower fibrinogen concentrations.2–4,40
The susceptibility of the fibrin matrix to proteolytic degradation may also impact cell proliferation rates. This study demonstrates that in fibrin formulations prepared with low levels of fibrinogen, cells generally proliferated at a rate comparable to that observed in monolayer culture, suggesting that easily degradable matrices do not impede cell proliferation rates to a high degree. However, when fibrin gels were prepared with 25 mg/mL fibrinogen, proliferation was delayed and was lower in magnitude on day 7, indicating that the matrix imparted a proliferation hindrance. These results are consistent with previous findings using other cell types.3–4
We provide evidence that the decreased proliferation observed in gels prepared with high fibrinogen concentrations is not simply due to variations in the number of surviving cells, as fibrin formulation has no effect on apoptotic activity levels. This result is consistent with previous studies using other cell types.2–4 Likewise, cell metabolic activity levels remain at least as high as those achieved in monolayer culture (67%) for the duration of the experiment. Interestingly, despite good cell viability, there is a reduction in metabolic activity observed in monolayer culture by day 7. A similar trend toward reduced metabolic activity is also observed in fibrin formulations prepared with low fibrinogen concentrations. Incidentally, these are the same culture conditions in which the greatest increases in cell proliferation and process extension are observed. It is possible that the decrease in cell metabolic activity in these conditions is due to confluency, as contact inhibition has been previously reported to result in increased enzyme-mediated hydrolysis of ATP when neural cells are cultured in vitro.41,42 The increased ATP hydrolysis that occurs when cells contact each other in confluent culture conditions would not be associated with an increase in caspase activity levels. 43
The influence of fibrin formulation on cell composition was also assessed in this work. Although the susceptibility of the fibrin matrix to cell-mediated degradation influences how readily entrapped cells extend processes and proliferate, the results presented here demonstrate that the biggest differences in cell composition correlate more with fibrinogen concentration and are more or less independent of degradation rate. Immunological characterization of the initial cell population indicated that ∼50% of the initial cell population was nestin-positive, ∼57% of the initial cell population was β-tubulin-positive, and 0% was GFAP-positive. The number of positive cells present on day 0 exceeds 100%, and this is likely due to some co-labeling of nestin and β-tubulin as NPCs transition into neuronal differentiation. Although double immunostaining was not performed in this work, co-labeling of these two markers during the transition between the precursor cell state and the postmitotic neuronal state has been reported previously. 44 The percentage of β-tubulin, nestin, and GFAP-positive cells is very consistent between cell isolations and agrees well with previously reported studies of this cell population from other groups using other techniques such as fluorescence activated cell sorting (FACs) analysis. 45
Fibrin formulation did not seem to influence the presence of nestin-positive NPCs or β-tubulin-positive neurons, though the presence of these immature cell types were decreased in fibrin gels relative to monolayer culture. In contrast, fibrin gel culture resulted in increased expression intensity of MAP-2, a marker of mature neurons that is present on increasing percentages of neurons in both fibrin gel and monolayer culture throughout the duration of the culture period. The increase in MAP-2 expression over time is consistent with other studies using neural cells isolated from rat embryos at 14–15 days of gestation.46,47 Levels of neurotransmitter-specific enzymes produced by mature, therapeutically relevant dopaminergic and cholinergic neuronal phenotypes were also increased in fibrin gel culture relative to monolayer culture. Interestingly, as fibrinogen concentration increased, the improvement in ChAT expression decreased, whereas the improvement in TH expression increased. We speculate that this may be related to differences in these two neuronal phenotype's need for glial-cell-derived trophic factors and aggregated cell growth, which are beneficial for dopaminergic survival and would increase in fibrin gels made with higher fibrinogen concentrations.48,49 This hypothesis is supported by the increased intensity of TH staining in the cell aggregates that grow within fibrin gels made from 25 mg/mL fibrinogen relative to the isolated cells that grow when fibrin gels are made with 5 mg/mL fibrinogen.
In contrast, GFAP mRNA that is expressed by glial cells which can contribute to glial scars that inhibit nerve regeneration was significantly lower in all fibrin formulations relative to monolayer culture. These robust differences in the neuronal and glial cell mRNA expression may be related to the large difference in the stiffness of fibrin gels compared to monolayer culture. It has been previously demonstrated that soft fibrin substrates do not negatively affect neuronal cell growth, but reduce actin-filament polymerization in glial cells, thereby discouraging glial cell growth. 16 Our study provides additional evidence that substrate stiffness influences the extent of glial cell growth, in that fibrin formulations with increased compressive moduli (higher fibrinogen concentration) resulted in higher GFAP gene expression than that observed in softer fibrin gels.
Although this work focused on the potential impact of mechanical properties of various fibrin formulations on neural cell behavior, another possible reason for the differences in neural cell behavior observed when cultured in various fibrin formulations and/or monolayer culture is the varying levels of fibrin ligand densities. Bioactive or adhesive sites present on fibrin can bind to receptors on cell surfaces, so the instructive signaling that the matrix provides could be ligand-receptor based rather than mechanical. In addition, the fibrinogen and thrombin solutions in the Tisseel kit are prepared by cryoprecipitation from blood plasma, an thus contain other proteins including transforming growth factor β-1, basic fibroblast growth factor, epidermal growth factor, and vascular endothelial growth factor. 48 Each of these proteins has been shown to impact the function of multipotent precursor cells or postmitotic neurons,30,50–58 and may be at least partially responsible for the observed changes in cell behavior in the different fibrin formulations and in monolayer culture.
In conclusion, the studies described here characterize the effect of fibrinogen and thrombin on the growth and composition of a mixed population of postmitotic neurons and multipotent NPCs in fibrin gels. This cell population is useful for neural cell transplant therapy, and the effect of fibrin formulation on this cell type has not yet been explored. Although the cells survive well in all formulations tested, important differences in proliferation and resulting cell composition were observed. Easily degradable fibrin gels prepared with lower fibrinogen concentrations facilitated greater cell proliferation and process extension. Relative to stiff monolayer culture, glial cell growth was suppressed, and the differentiation of two clinically relevant neurotransmitter-expressing neurons (dopaminergic and cholinergic neurons) was promoted in a manner that was dependent on the concentration of fibrinogen or thrombin. The findings here suggest the importance of fibrinogen and thrombin concentration in differentially regulating the growth and differentiation of neural cell populations containing NPCs and are of importance for neural tissue engineering strategies focused on developing implantable cell scaffolds enriched in neurons that tend to be lost during the progression of Parkinson's disease and Alzheimer's disease.
Footnotes
Disclosure Statement
No competing financial interests exist.