
Abstract
Embryonic stem cells (ESCs) can have unlimited proliferation potential and the capability to differentiate into almost all types of tissues. The use of ESCs as a novel source of hepatocytes has therefore recently received much attention. To date, however, the efficient and mass differentiation of ESCs into hepatocytes in vitro has been proven to be difficult. In this study, we investigated a three-dimensional (3D) mouse ESC hepatic differentiation system. Cells derived from 5-day-old embryoid bodies were mixed with Matrigel and immediately seeded in a biodegradable polymer scaffold composed of poly-
Introduction
Embryonic stem cells (ESCs) derived from the inner cell mass of blastocysts have unlimited proliferation potential in vitro, while retaining the potential to differentiate into all types of cells, including hepatocytes. 4 Several recent studies have reported the differentiation of hepatocyte-like cells from human or mouse ESCs.5–12 However, most studies demonstrating hepatic differentiation from ESCs have been based on common laboratory procedures carried out in two-dimensional (2D) cell culture systems such as T-flasks, which are limited by the available surface area and difficulties involved in scaling up the procedures. Further, hepatocyte-like cells in conventional monolayer culture systems require harsh treatment to dislodge them from the culture dish before use or prior to transplantation. This process can injure the cells or destroy tissue structures. 13 The efficient and mass differentiation of ESCs into hepatocytes in vitro has therefore proven difficult. Moreover, these strategies have ignored the fact that extensive cell–cell and cell–matrix interactions in vivo are important for the maintenance of hepatocyte functions and the maturation of hepatic progenitors. 14 Thus, mass and long-term cultivation of healthy and functional ESC-derived hepatocyte-like cells presents a challenge to conventional hepatic differentiation methods.
We hypothesized that porous biodegradable polymer scaffolds containing Matrigel and appropriate, dissolved inducing factors could be used to support and induce the hepatic differentiation of mouse ESCs, because of their resemblance to the cellular environment in developing liver tissue. The scaffold provides physical cues for cell orientation and spreading, while the pores provide space for remodeling of tissue structures. Different cocktails of growth factors and hormones could allow stepwise hepatocyte specification of ESCs in the three-dimensional (3D) scaffold systems. This system could lead to highly efficient and mass hepatic differentiation of ESCs.
To test the hypothesis, embryoid bodies (EBs) derived from mouse ESCs were formed using the hanging drop method. These EBs were trypsinized and cultured in a 3D culture system composed of biodegradable polymer scaffolds and Matrigel, together with exogenous growth factors and hormones, to promote hepatic histogenesis.
Materials and Methods
Mouse ESC culture and EB formation
Mouse ESCs (CCE line; StemCell Technologies, Vancouver, Canada) were maintained in Iscove's modified Dulbecco's medium (IMDM; Gibco/Invitrogen, Carlsbad, CA) supplemented with 15% fetal bovine serum, 0.1 mM nonessential amino acids, 2 mM
Polymer scaffolds
The nonaligned scaffolds were fabricated from small-diameter fibers of uniform length. Using a nonwoven textile process, the fibers were randomly entangled to form a strong, flexible, and porous 3D matrix. Scaffolds comprised biodegradable copolymers composed of 50% poly-
Hepatic differentiation on Matrigel and scaffolds
For seeding of cells on scaffolds, 5-day-old EBs were trypsinized and EB-derived cells were resuspended at a concentration of 1 × 106 cells per mL in a 1:1 mixture of culture medium and growth factor-reduced Matrigel (BD Biosciences, San Jose, CA) and were then seeded onto polymer scaffolds. After incubating for 30 min at 37°C, the scaffolds were transferred into 12-well plates and cultured with IMDM without LIF. Growth factors were added to the medium at the following times to induce hepatic differentiation: 10−7 M dexamethasone and 1% dimethyl sulfoxide were added on days 1–4; 10 ng/mL fibroblast growth factor 4 (Sigma-Aldrich, St. Louis, MO) was added on days 5–10; 20 ng/mL recombinant mouse hepatic growth factor (R&D Systems, Minneapolis, MN) was added on days 8–12; 10−7 M dexamethasone, 1% insulin–transferrin–selenium (Gibco), and 10 ng/mL oncostatin M (R&D Systems) were added on days 10–20.
Scanning electron microscopy examinations
Cell–scaffold constructs were fixed with 0.25% glutaraldehyde solution and dehydrated in a series of graded ethanols and tert-butyl alcohol at 37°C. Dried constructs were sputter-coated with gold/palladium and examined using a Hitachi S-3400N scanning electron microscope (Central Laboratory of Third Military Medical University, Chongqing, P.R. China) operated at 15 kV.
Reverse transcription–polymerase chain reaction analysis
Total RNA was extracted using an RNeasy Mini Kit (Qiagen, Valencia, CA) and treated with RNase-free DNase (Qiagen). Reverse transcription–polymerase chain reaction (RT-PCR) was carried out using a Qiagen OneStep RT-PCR Kit with the addition of 10 U RNase inhibitor (Invitrogen) and 20 ng RNA, according to the manufacturer's instructions. The primer sequences and annealing temperatures used were as follows: albumin (ALB; sense, 5′-GACAAGGAAAGCTGCCTGAC-3′; antisense, 5′-TTCTGCAAAGTCAGCATTGG-3′; 59.7°C), α-fetoprotein (AFP; sense, 5′-AGCAAAGCTGCGCTCTCTAC-3′; antisense, 5′-GAGTTCACAGGGCTTGCTTC-3′, 55°C), transthyretin (TTR; sense, 5′-CTCACCACAGATGAGAAG-3′; antisense, 5′- GGCTGAGTCTCTCAATTC-3′; 52.3°C), glucose-6-phosphatase (G6P; sense, 5′-CAGGACTGGTTC ATCCTT-3′; antisense, 5′-GTTGCTGTAGTAGTCGGT-3′; 58°C), beta-actin (sense, 5′-TGTTACCAACTGGGACGACA-3′; antisense, 5′-GGGGTGTTGAAGGTCTCAAA-3′; 55°C).
Immunostaining
For immunostaining, cultured cell scaffolds obtained on day 20 were embedded in optimal cutting temperature compound and stored at −80°C until use. Optimal cutting temperature-embedded samples were cut using cryostat into 8-mm-thick sections and mounted on poly-
Western blot analysis
Western blot analysis was performed to detect ALB protein in mouse ESCs. In brief, the protein concentration in cell lysates was measured using the Bio-Rad Protein Assay Kit (Bio-Rad Laboratories, Hercules, CA). About 20 μg of protein from each supernatant was subjected to 4–15% Tris-HCl gel electrophoresis (Bio-Rad Laboratories) and then transferred to a nitrocellulose membrane (Pierce Biotechnology, Rockford, IL). The membrane was blocked in 2.5% milk-blocking buffer. The blot was probed with goat anti-rat ALB antibody (1:10,000; MP Biomedicals, Aurora, OH). The blots were then incubated with a horseradish peroxidase-conjugated donkey anti-goat IgG (Santa Cruz Biotechnology, Santa Cruz, CA). The protein bands were visualized by means of enhanced chemiluminescence (Pierce Biotechnology) and the images were captured using medical X-ray film.
Assessment of liver-specific function in vitro
Three function tests were carried out on cells in 3D Matrigel and scaffold cultures on days 5, 10, 15, and 20 to determine if the differentiated cells were functional. The amount of ALB secreted into the medium was quantified using a mouse ALB enzyme-linked immunosorbent assay quantitation kit (Bethyl) under the conditions recommended by the manufacturer. The ability of differentiated cells to take up low-density lipoprotein (LDL) was assessed by DiI-Ac-LDL (acetylated low density lipoprotein, labeled with 1,1′-dioctadecyl-3,3,3′,3′-tetramethyl-indocarbocyanine perchlorate) staining. In brief, the 3D cultured scaffolds were incubated with 20 μg/mL DiI-Ac-LDL (Biomedical Technologies, Stoughton, MA) for 4 h at 37°C. Incorporation of fluorochrome-labeled LDL into cells was visualized using a confocal laser scanning microscope (CLSM; model IX71; Olympus, Tokyo, Japan). Meanwhile, DiI-Ac-LDL–stained scaffolds were placed in a medium containing 0.1 mM calcein-AM (Dojindo Molecular Technologies, Gaithersburg, MD) and incubated for 20 min at 37°C to determine cell viability. Glycogen production was analyzed by periodic acid-Schiff (PAS) staining after inducing hepatic differentiation. PAS staining was performed on tissue sections using a PAS stain kit (American MasterTech Scientific, Lodi, CA). The ratio of hepatic differentiation was determined by counting the PAS-positive cells on day 20 of cell differentiation. 15 Spontaneously differentiated cells cultured with the same 3D scaffold, but without any growth factors or hormones, were used as controls.
Results
Cell growth and morphology of mouse ESCs in Matrigel and scaffold culture
After their formation, 5-day-old EB-derived cells were cultured and differentiated in the Matrigel and scaffold for 20 days (Fig. 1). During 3D culture, mouse ESCs grew and proliferated in two ways. First, mouse ESCs spontaneously formed spherical multicellular aggregates (spheroids) in the pores of the polymer scaffolds. The spheroids grew larger, and some spheroids adhered to each other. Second, mouse ESCs spontaneously formed a mesh between or along the fabric fibers of the scaffolds. At a later stage of culture, the mouse ESCs not only established a 3D network but also formed irregular cell aggregates within the polymer scaffolds, and the density of the cells increased over time (Figs. 2 and 3). Some teratoma-like tissues were observed in some sections after prolonged periods in culture. We attempted to prolong the culture period to 30 days, but increasing amounts of dead tissue accumulated with increasing culture duration. This was especially marked in the center of the cultures.

Experimental scheme used for the differentiation of mouse ESCs into hepatocytes in 3D polymer scaffolds. 3D, three-dimensional; DEX, dexamethasone; DMSO, dimethyl sulfoxide; EBs, embryoid bodies; ESCs or ES cells, embryonic stem cells; FGF4, fibroblast growth factor 4; HGF, hepatocyte growth factor; ITS, insulin–transferrin–selenium; LIF, leukemia inhibitory factor; OSM, oncostatin M.
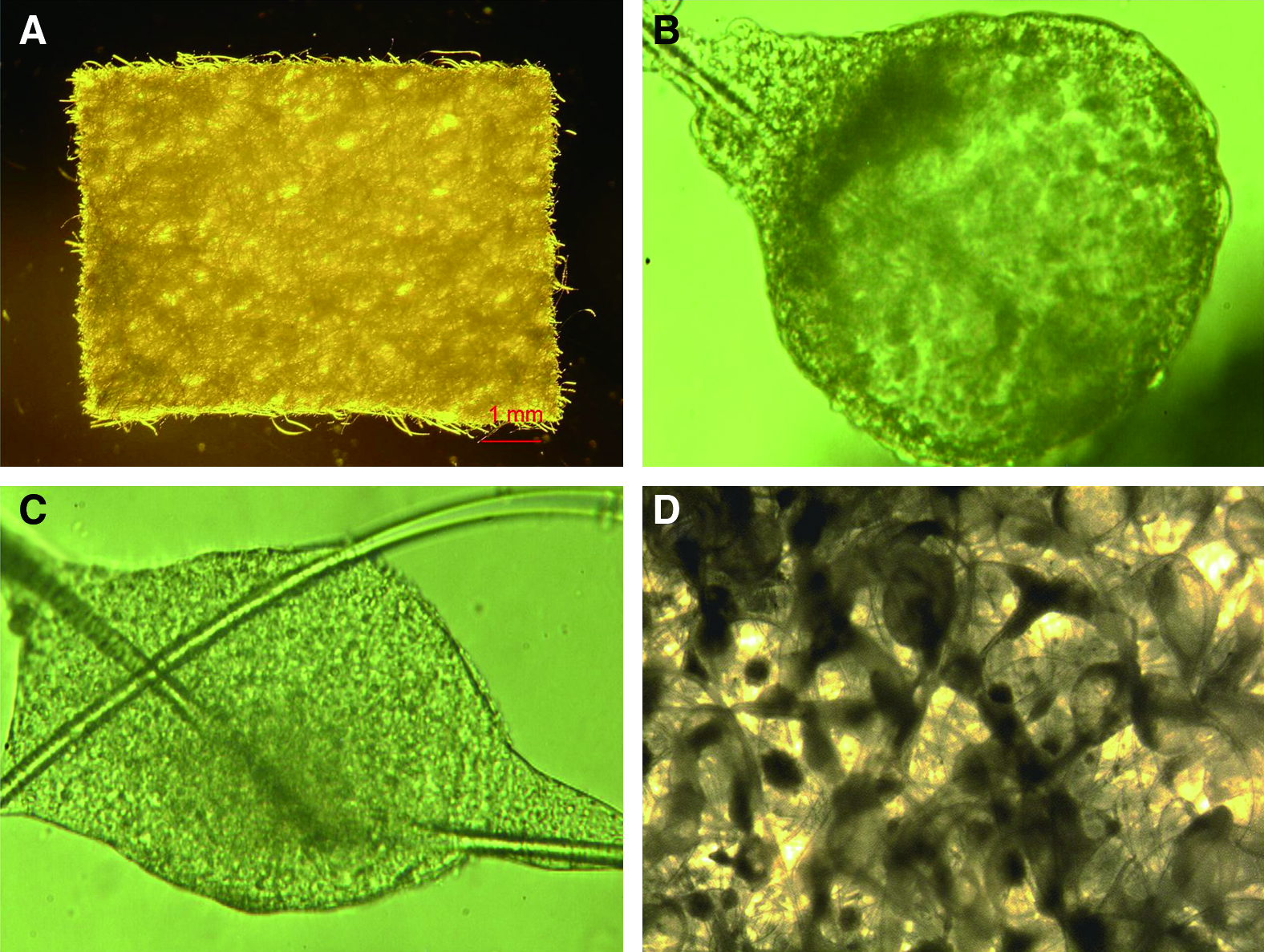
Cell growth and morphology of mouse ESCs in Matrigel and scaffold culture. (
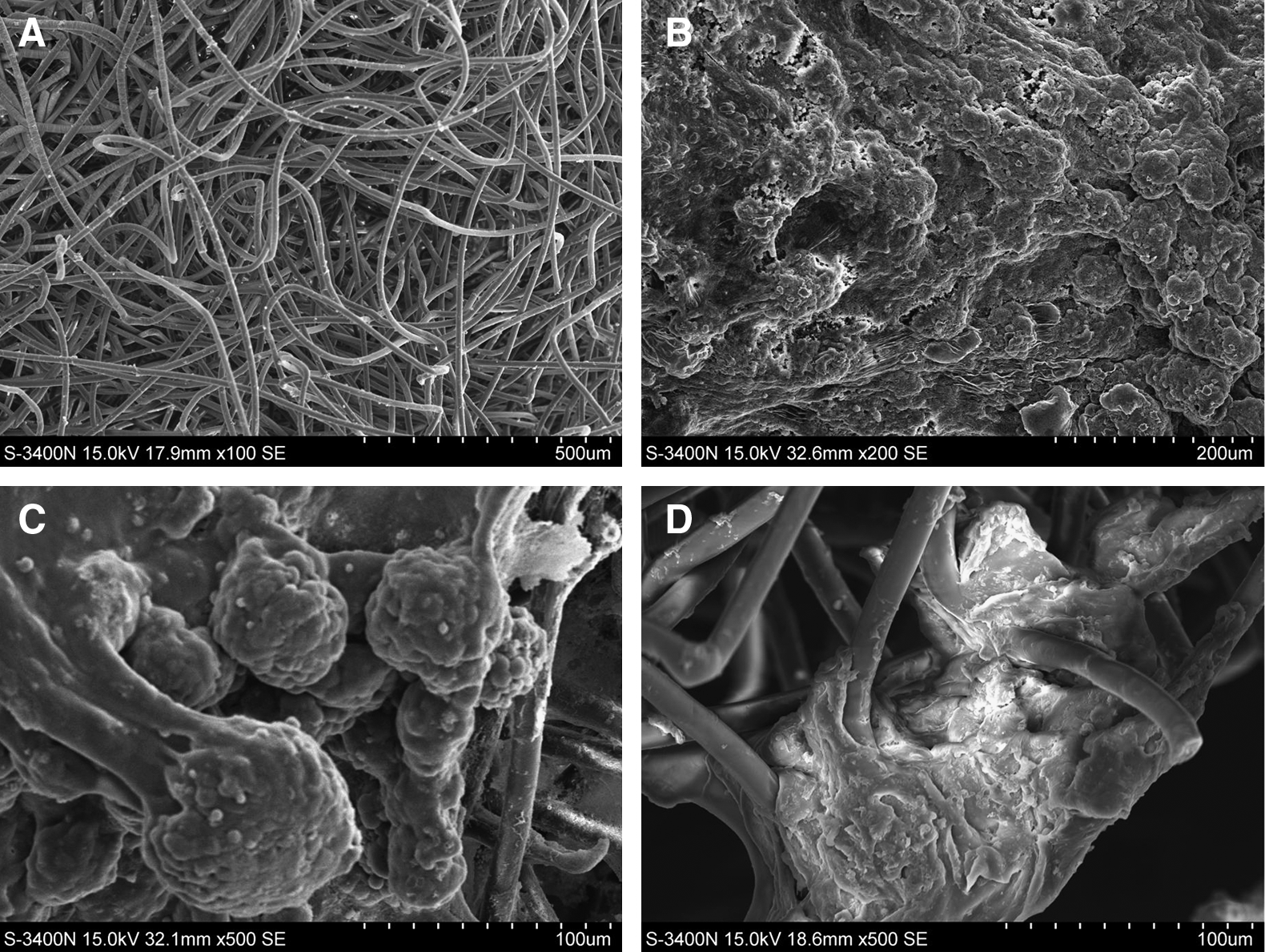
Scanning electron micrographs showing the morphology of mouse ESCs in 3D scaffold cultures. (
Gene expression in mouse ESC-derived hepatocytes
To assess the differentiation of mouse ESCs into hepatocyte-like cells, we used RT-PCR to analyze endodermal- and liver-specific genes (Fig. 4A). Total RNA was isolated on days 0, 5, 10, 15, and 20 of the 3D scaffold cultures. ALB, AFP, G6P, and TTR were used as endodermal- and liver-specific gene markers. TTR represents a marker for endodermal or yolk sac-like differentiation and is expressed throughout liver maturation; AFP is an endodermal differentiation marker and an early fetal hepatic marker; G6P is predominantly expressed in hepatocytes in late gestation; ALB is a hepatocyte-specific gene. Figure 4A shows the expression of endodermal and hepatic differentiation markers in differentiating mouse ESCs. AFP and TTR were expressed from days 5 to 20. ALB and G6P were expressed from days 10 to 20. Both endoderm-specific and hepatocyte-specific gene expression increased during culture of these cells. Undifferentiated mouse ESCs did not express either endodermal- or hepatocyte-specific genes. These data indicate that mouse ESCs can differentiate toward the hepatic lineage and produce mature hepatocytes in 3D Matrigel and scaffold cultures.
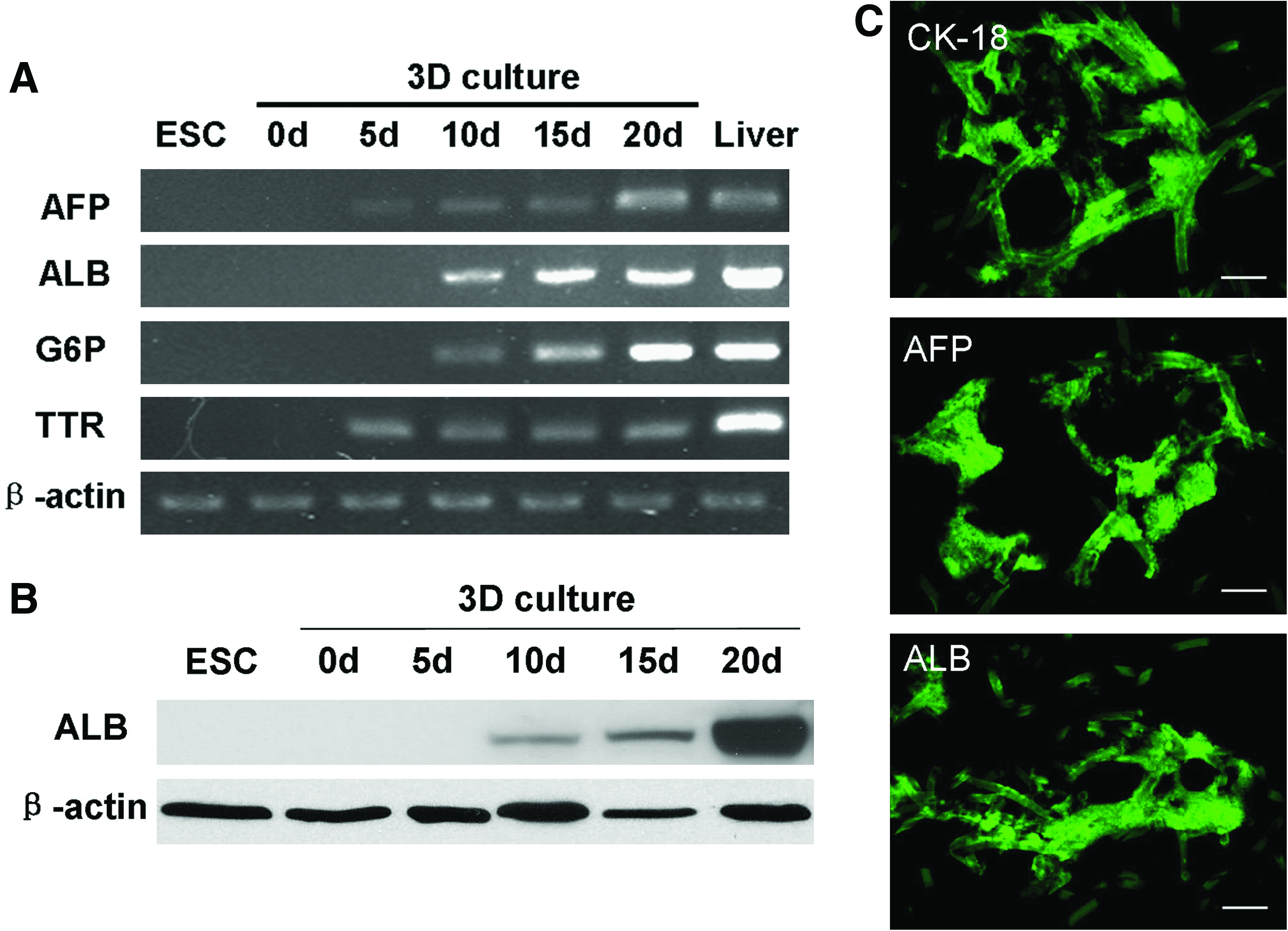
Analysis of gene expression using different methods. (
Immunostaining and western blot analysis
Figure 4C shows the results of immunofluorescence staining of CK18, AFP, and ALB in differentiating mouse ESCs. Intense staining for CK18, AFP, and ALB proteins was observed in cryostat-cut sections of the day-20 3D constructs. To further assess the differentiation of mouse ESCs into hepatocyte-like cells at the protein level, ALB expression in differentiated cells was analyzed by western blotting (Fig. 4B). Undifferentiated mouse ESCs did not express ALB protein. However, the protein band representing ALB was visible in EB-derived cells on day 10 of the 3D scaffold cultures and reached a maximum level on day 20. There results demonstrate the hepatic maturation of mouse ESCs in 3D scaffold cultures.
Evaluation of liver-specific functions
ALB production is a unique function of hepatocytes. To determine if the differentiated mouse ESCs in 3D scaffold cultures were functional hepatocytes, ALB production was measured using a competitive enzyme-linked immunosorbent assay method (Fig. 5A). No ALB secretion was detected in the differentiating mouse ESCs on days 0, 5, and 10, but it was present on days 15 and 20. The ALB quantity secreted on day 20 was four- to fivefold higher than that secreted on day 15. Glycogen production is a major metabolic function of hepatocytes. No stored glycogen was observed in undifferentiated mouse ESCs, but it was present in 3D scaffold cultures of mouse ESCs after 15 days of differentiation (Fig. 5B). PAS-positive cells in cultures at day 20 of differentiation and in control cultures were 56.5% ± 12.7% and 6.6% ± 1.8%, respectively (data are expressed as mean ± standard deviation, n = 5, p < 0.01). LDL is a major type of lipoprotein that carries cholesterol to peripheral tissues for use by various cells. Most LDL is metabolized by hepatocytes. The ability of differentiated cells to take up LDL was assessed by DiI-Ac-LDL staining. No DiI-Ac-LDL uptake was detected in 3D scaffolds cultured for 0, 5, or 10 days, or in undifferentiated mouse ESCs, but uptake was detected in differentiated cells on day 15 by CLSM (Fig. 5C).
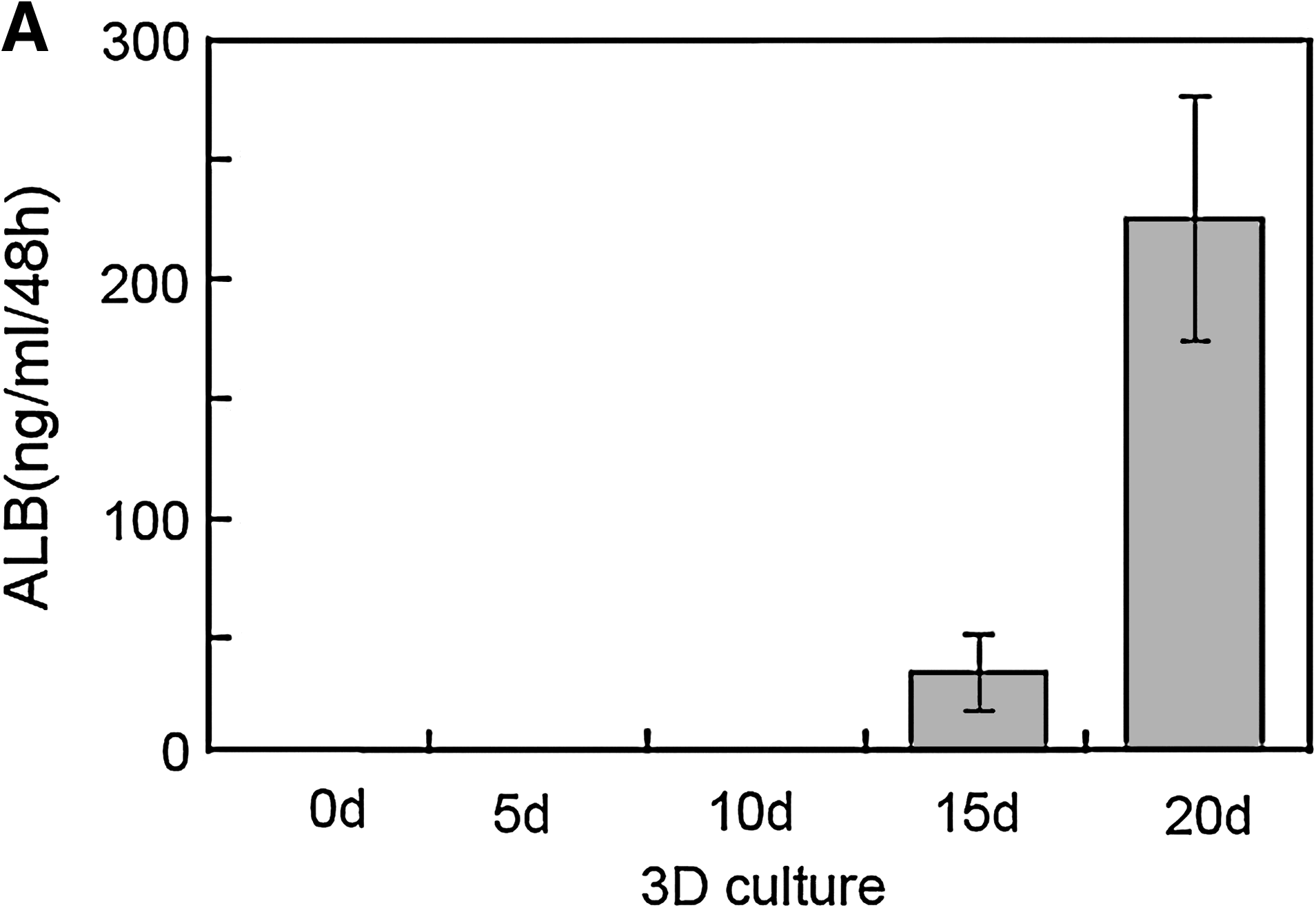
Evaluation of ALB secretion, glycogen storage, and LDL uptake. (
Discussion
Regulation of embryogenesis involves extracellular matrix (ECM) proteins, growth factors, signaling molecules, hormones, transcription factors, cell–cell interactions, and biomechanical forces acting in temporally and spatially organized processes.16,17 Most regulators operate within 3D microenvironments in the body. Recent studies have demonstrated the advantages of a 3D environment for producing specified cells from ESCs.18,19 In this study, we focused on creating a 3D supportive environment suitable for directing the differentiation and organization of mouse ESCs into hepatocytes or liver-like structures.
We used biodegradable PLLA/PGA polymer scaffolds. The scaffolds provided a 3D spatial environment allowing the cells to grow and attach, while nutrient flow was maintained throughout the matrix. The pores between the spun fibers were created using a nonwoven textile process and facilitated the seeding and ingrowing of cells. Scaffolds were seeded with 5-day-old EB-derived cells and Matrigel and cultured with sequential supplements of exogenous growth factors and hormones for 20 days.
EB-derived cells in the scaffold culture system expressed several liver-specific mRNAs. TTR and AFP, which are markers of early hepatocyte differentiation, were expressed from days 5 to 20. Other hepatocyte-specific genes, including ALB, an early fetal and mature hepatocyte differentiation marker, were expressed from days 10 to 20. G6P, which is predominantly expressed in hepatocytes during late gestation or at the perinatal stage, was also expressed from days 10 to 20. Although TTR, AFP, and ALB are sometimes expressed in other cells, G6P is selectively expressed in the liver. Expression of ALB, AFP, and CK18 on day 20 was confirmed by immunofluorescence staining. The results of western blot analysis further confirmed the expression of ALB at the protein level and were consistent with the results of RT-PCR analysis. Thus, the pattern of gene expression in cells derived from EBs was characteristic of differentiated hepatocyte-like cells.
However, the aforementioned genes can also be expressed during the development of nonhepatocytes and their expressions are therefore not conclusive evidence of hepatocyte differentiation. Hepatocyte-specific functions have been regarded as the gold standard for assessing the presence of mature hepatocytes. ALB and glycogen production and LDL uptake tests were therefore also performed. ALB secretion increased rapidly at day 20. Glycogen production was also detected in 3D scaffold cultured cells at day 15. LDL uptake, which is a liver-specific function, was assessed by DiI-Ac-LDL uptake and was detected in 3D scaffold cultured cells on day 15 by CLSM. These results confirmed the differentiation of ESCs into hepatocytes in these cultures.
Hepatic differentiation has recently been achieved in 3D cultures in several studies.13,20–22 These studies have provided valuable information regarding the effective hepatic differentiation and large-scale production of hepatocytes from ESCs. In these studies, however, undifferentiated ESCs or whole EBs were directly immobilized within different types of scaffolds with no predifferentiation, no separation of EBs, and no accompanying ECM. Scaffolds seeded with undifferentiated ESCs or whole EBs may exhibit more heterogeneous differentiation, and less uniform growth and survival compared with differentiating ESCs. 23 The interaction between cells and ECM plays an important role in inducing cell differentiation; the developmental fate of ESCs is determined not only by soluble factors, but also by physical interactions with the surrounding ECM. Many studies have highlighted the benefit of Matrigel, a basement membrane extract from the Engelbreth–Holm–Swarm mouse sarcoma, which serves as an ECM and effectively promotes the attachment and differentiation of hepatocytes.24–26 Further, Matrigel can capture EB-derived cells in a 3D microenvironment that can control cell density. This process is emerging as a new means of regulating differentiation. 27
Levenberg et al.23,28 examined the multipotential differentiation and neurotrophin-induced differentiation of ESCs in biodegradable copolymer PLLA/PGA scaffolds and ECM, but to the best of our knowledge, no one has reported the hepatic differentiation of ESCs in the same 3D environment. Our results show that copolymer PLLA/PGA scaffolds provide a good environment for the 3D growth, proliferation, and differentiation of mouse ESCs. Our results also show that both cell–cell and cell–matrix interactions in the 3D environment promote hepatic differentiation of mouse ESCs.
Both Matrigel and PLLA/PGA can capture mouse ESCs in 3D microenvironments. Mouse ESCs can spontaneously form spheroids or a mesh in the pores of the polymer and along or between the fabric fibers. Growth factors and hormones were used to further stimulate specific hepatic differentiation in the polymer scaffolds. However, teratoma-like or dead tissues were found in the scaffolds after a prolonged period in culture. Few previous studies have reported on the long-term fate of ESC cultures after completion of the hepatic differentiation procedure. Rapid loss of differentiation may be a fundamental process, because even primary hepatocytes lose normal hepatocyte structure and function in long-term 2D cultures. Alternatively, the 3D cultures may experience problems due to lack of optimal nutrition and oxygenation in the center of the growing spheroids. Teratoma-like tissue may develop because of heterogeneous differentiation. In addition, our induction cocktails of growth factors and hormones were based on modifications of methods 29 previously used in 2D cultures; recent novel hepatocyte differentiation strategies10–12 may prove to be more suitable than that used in our study. However, these novel studies used 2D human ESC cultures, and their relevance to hepatic differentiation in 3D cultures remains unknown. Further studies are therefore needed to determine the optimal induction cocktails and nutritional support and also the most suitable stereochemical scaffold structure needed to promote functional hepatocyte development in 3D human ESC cultures.
In conclusion, both PLLA/PGA and Matrigel have been shown to provide adequate and stable microenvironments for the growth and differentiation of mouse ESCs in a 3D model. With the stepwise addition of stimulating factors, EB-derived cells were able to differentiate into hepatocyte-like cells. The strategy has several potential advantages: liver-like tissues with large numbers of hepatocyte-like cells can be obtained rapidly in 3D cultures; hepatocyte-specific functions can be maintained for longer, because the technique recreates conditions resembling natural liver development and architecture found in the living organism. Further studies of 3D cultures are needed to determine the optimal mixtures of inducing factors. The results of this study suggest that this method combining PLLA/PGA polymer scaffolds, Matrigel, growth factors, and hormones may provide a novel alternate method for promoting large-scale differentiation of mouse ESCs into functional hepatocytes or liver tissue.
Footnotes
Acknowledgment
This study was supported by the National Science Foundation of China (grant no. 30670526).
Disclosure Statement
No competing financial interests exist.