
Abstract
Baculovirus efficiently transduces human mesenchymal stem cells (hMSCs) and transplantation of hMSCs transduced with a bone morphogenetic protein 2–expressing baculovirus (Bac-CB) into nude mice results in ectopic bone formation. To attest the clinical potential of baculovirus in bone regeneration, hereby we explored whether the hMSCs genetically modified by Bac-CB were tolerant in immunocompetent rats and further healed the critical-sized calvarial bone defect. The histological and computed tomographic studies demonstrated that Bac-CB–engineered hMSCs promoted the cell differentiation and new bone formation in the immunocompetent rats. Immunohistochemical staining revealed that the transplanted human cells remained detectable at 1 and 4 weeks posttransplantation, attesting the immunoprivileged properties of hMSCs. In the recipients, the donor cells aggregated and appeared osteoblast like at later stages, which paralleled the infiltration of macrophages, CD3+, and CD8+ T cells into the graft. Administration of immunosuppressive drugs prolonged the cell survival and improved the bone regeneration, yet it failed to entirely abolish the immune response and complete the bone healing. Our data altogether implicate the potential of Bac-CB for hMSCs engineering and calvarial bone repair, but the use of hMSCs cannot overcome the immunological barrier.
Introduction
Aside from these vectors, baculovirus (Autographa californica multiple nucleopolyhedrovirus) is an insect virus, but it also efficiently transduces a wide variety of mammalian cells without replication and appreciable cytotoxicity (for review see Refs.9–11 ). These features have inspired the development of baculovirus vectors carrying mammalian expression cassettes for in vitro and in vivo gene therapy studies, development of cell-based assays, surface display of eucaryotic proteins, study of gene functions, production of viral vectors, production of virus-like particles, delivery of vaccine immunogens (for review see Refs.10,12), and genetic modification of chondrocytes for cartilage regeneration.13,14 Further, baculovirus transduces human bone marrow–derived MSCs (i.e., hMSCs) at efficiencies up to 95% under optimized conditions, 15 and the transduced hMSCs remain capable of differentiation into adipogenic, osteogenic, and chondrogenic lineages. 16 Given these findings, we constructed a recombinant baculovirus (Bac-CB) expressing bone morphogenetic protein-2 (BMP-2) under the control of cytomegalovirus immediate-early promoter. Bac-CB transduction of hMSCs ex vivo triggered in vitro differentiation of hMSCs into osteoblasts, and xenotransplantation of the transduced cells into the back subcutis of immunodeficient nude mice resulted in ectopic bone formation. 17
Many studies exploiting hMSCs for tissue repair have transplanted the cells into immunodeficient or allogeneic animals to circumvent the immune attack. Although hMSCs are immunosuppressive and can be tolerated after allotransplantation, whether hMSCs are suitable for xenotransplantation remains to be established. For instance, hMSCs were depleted rapidly after direct injection into the myocardium of rats,18,19 while xenotransplantation of rat MSCs into the quadriceps of immunocompetent mice led to elevated immune responses that obstructed bone formation, although some donor cells remained detectable at week 11. 20 In contrast to these results, injection of hMSCs into the liver of Sprague–Dawley rats led to the engraftment and differentiation of cells into human hepatocytes with the aid of immunosuppression. 21 Xenotransplantation of hMSCs into the rat spinal cord led to long-term survival of cells under, but not without, appropriate immunosuppression. 22 Strikingly, intravenous injection of hMSCs into the rats led to the survival and migration of hMSCs to the spinal cord injury site and improved the functional recovery even without immunosuppression. 23 A recent study also reported the long-term survival and chondrogenic differentiation of hMSCs after xenotransplantation into the rat intervertebral discs without immunosuppression. 24
To attest the clinical potential of baculovirus in hMSCs engineering and bone regeneration, and to address the question regarding whether the immunoprivileged hMSCs can escape the immune surveillance in the xenogeneic setting, the primary objective of the present study was to evaluate the calvarial bone repair mediated by the Bac-CB–transduced hMSCs in the immunocompetent Fisher 344 rats, in the presence and absence of immunosuppressive drugs.
Materials and Methods
Preparation and culture of bone marrow–derived hMSCs
Bone marrow–derived human mononuclear cells were obtained from Cambrex (Walkersville, MD) and the subsequent hMSCs selection, enrichment, and immunotyping were performed as described. 25 The resultant hMSCs were cultured using α-modified minimal essential medium (Hyclone, Ogden, UT) containing 20% fetal bovine serum (Invitrogen, Carlsbad, CA), 4 ng/mL basic fibroblast growth factor (R&D System, Minneapolis, MN), 100 U/mL penicillin, and 100 mg/mL streptomycin in a 37°C, 5% CO2 incubator. The hMSCs were expanded to passage 10 for all subsequent experiments.
Baculovirus preparation and transduction
The recombinant baculovirus (Bac-CB) expressing BMP-2 was constructed earlier. 17 The virus titers (pfu/mL) were determined by end-point dilution method, 26 and the baculovirus transduction was performed as described 17 with minor modifications. In brief, hMSCs were cultured overnight in the T-150 flasks (6 × 106 cells per flask) and washed with Dulbecco's phosphate-buffered saline (PBS, pH 7.4) before transduction. For each flask, a certain volume of virus was diluted to 2 mL with modified Grace's Insect medium (TNM-FH) to adjust the multiplicity of infection (MOI) to 40, followed by mixing with 8 mL PBS. Transduction was initiated by directly adding the virus–PBS solution to the cells and continued by gentle shaking on a rocking plate at room temperature for 4 h. After the incubation period, the cells were washed, replenished with 20 mL α-modified minimal essential medium, and incubated at 37°C. For mock transduction, the cells were incubated with 2 mL TNM-FH medium plus 8 mL PBS for 4 h at 37°C. Supertransduction was performed at 6 days after initial transduction in a similar manner.
Poly (L-lactide-co-glycolide) scaffold preparation and cell seeding
To fabricate the porous poly (L-lactide-co-glycolide) (PLGA) scaffolds, the PLGA (Purac, Gorinchem, The Netherlands) dissolved in chloroform (0.1 g/mL) was mixed with sodium chloride (200–300 μm in diameter) and the solution was compressed into the Teflon mold (9 mm in diameter). After the chloroform vaporized, the scaffolds were immersed in double-distilled water with three changes to dissolve the sodium chloride, and gently removed from the Teflon mold with a fin-tip spatula. The resultant scaffolds (porosity ≈90%) were disinfected by immersion in 50%, 60%, and 70% ethanol for 30 min each, rinsed with double-distilled water and air dried in a laminar flow hood.
One day after supertransduction, 50 μL hMSCs were evenly pipetted onto the scaffold (5 × 105 cells per scaffold), allowed to adhere for 2 h and then supplemented with fresh medium for culture. One day later, the cell/scaffold constructs were transplanted into the calvarial defects.
Transplantation and immunosuppression
All animal experiments were performed in compliance with the Guide for the Care and Use of Laboratory Animals (Institute of Laboratory Animals Resources, National Science Council, Taiwan). Female Fisher 344 rats (10 weeks of age) were randomly assigned to designated groups. After anesthetization, 8-mm-diameter critical defects 1 were created in the calvaria, implanted with the blank PLGA scaffolds or the scaffolds impregnated with hMSCs, and the wounds were closed with suture. The immunosuppression scheme consisted of subcutaneous injection of 1 mg/kg of antithymocyte globulin (Genzyme Polyclonals S.A.S., Marcy l'Etoile, France) from 2 days before transplantation to 7 days after transplantation; daily oral administration of mycophenolate mofetil (Roche, Mannheim, Germany) dissolved in the drinking water (1 mg/mL); and daily subcutaneous injection of 10 mg/kg of cyclosporine (Roche). Tissue specimens were harvested at 1, 4, 8, and 12 weeks (n = 5 for each group at each time point) after transplantation.
Microcomputed tomography analysis
The animal computed tomography (CT) scan was performed using NanoSPECT/CT (Bioscan, Washington, DC) at the Molecular Imaging Center of Chang Gung Memorial Hospital. The CT system contained a power-adjustable X-ray source of 65 kVp and microfocus (<9 μm) tube, and the resolution was less than 100 μm. The CT image intensity was calibrated into Hounsfield scale using water (0) and air (−1000) as references. The CT data of the rat skull were acquired using a high-resolution frame as setup in the system, with tube voltage of 65 KeV, exposure time of 2000 ms, and 180 projections. The CT images were reconstructed using the software provided by the system using an ultrafine reconstruction mode with 512 × 512 × 480 voxels and 0.1 mm voxel size.
The new bone formation within the defect was quantified on a PMOD image workstation (PMOD Technologies, Zurich, Switzerland). The skull CT images were loaded, and the center of scaffold was identified. A volume of interest (VOI) in rectangular shape was defined to cover the defect and the scaffold. The VOI was then segmented to eliminate the unwanted part of the skull. The maximum intensity project of the segmented VOI was generated, and the percentage of new bone formation was calculated from the maximum intensity project images using the following formula:
% new bone formation = (area of new bone formation)/(area of original defect) × 100%
The contour of the new bone formation was defined by the absolute threshold above 200 Hounsfield units.
Hematoxylin and eosin and immunohistochemical staining
The tissue specimens were fixed, decalcified, and sectioned. The sections were deparaffinized and rehydrated using xylene and a gradient of ethanol (100%, 90%, 70%, 50%, and 0%), followed by hematoxylin and eosin staining to detect osteogenesis. For immunohistochemical staining, the sections were treated with the Target Retrieval solution (Dako, Hamburg, Germany) for 10 min at 90°C and then blocked with PBS containing 10% fetal bovine serum for 30 min at room temperature. The sections were incubated at 4°C overnight with the primary mouse monoclonal antibody specific for human mitochondria (1:100 dilution; Bio Genex, San Ramon, CA), rat macrophage (1:100 dilution; Abcam, Cambridge, United Kigdom), rat CD3 (1:10 dilution; AbD Serotec, Oxford, United Kingdom), or rat CD8 (1:100 dilution; Gene Tex, San Antonio, TX). After washing, the sections were incubated with goat anti-mouse IgG conjugated with Alexa Fluor 488 (Invitrogen) for 1 h in the dark. Finally, the sections were mounted with the mounting medium containing 4′,6-diamidino-2-phenylindole (Vector Labs, Burlingame, CA) and observed using a fluorescence microscope.
Results
Effects of Bac-CB transduction and immunosuppression on calvarial bone formation
The calvarial bone formation starts by the migration and aggregation of mesenchymal cells (for review see Ref. 27 ). The progenitor cells differentiate into osteoblasts and secrete the bone matrix (osteoid) that initially appears as small, irregularly shaped spicules. The spicules undergo calcification with time, enlarge, and become joined in trabecular network. During the process, the osteoblasts actively secreting matrix molecules surround the growing spicules whereas some osteoblasts are enclosed within the matrix and differentiate into mature osteocytes.
To explore whether the baculovirus-transduced hMSCs were tolerated in immunocompetent rats and repaired the calvarial bone defect, the hMSCs were transduced by Bac-CB at an MOI of 40 and were supertransduced (MOI 40) 6 days later. The transduced and mock-transduced hMSCs were seeded to PLGA scaffolds and transplanted into the critical defects (8 mm) at the rat calvaria (n = 5 for each group at each time point). As controls, blank PLGA scaffolds were transplanted in a similar fashion.
The hematoxylin and eosin staining (Fig. 1A) revealed that the control groups (scaffold only and mock-transduced hMSCs) barely led to cell aggregation at 4 weeks after transduction, and only gave rise to a few osteoids at week 12. In sharp contrast, the transduction group resulted in conspicuous formation of numerous calcified bone matrices with osteoblast-like cells lining the matrix (indicated by the arrows) at week 4. However, at week 12 a considerable fraction of the calcified bone matrix disappeared and no further bone formation was observed.
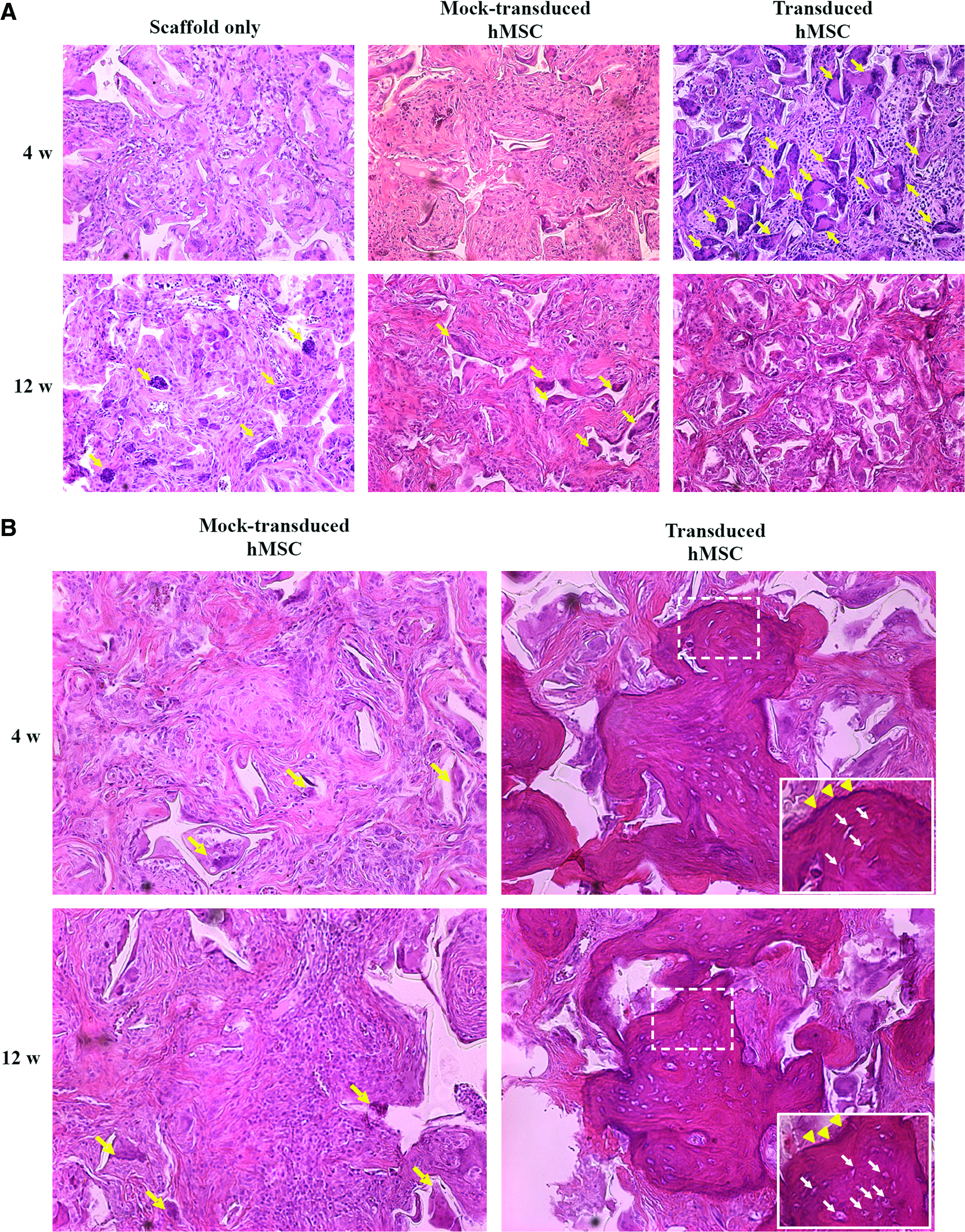
Effects of Bac-CB transduction on calvarial bone formation in the absence (
It is well established that xenotransplantation causes rejection and failed engraftment. Therefore, the experiments were repeated as in Figure 1A, but the rats were administered with immunosuppressive drugs (antithymocyte globulin, mycophenolate mofetil, and cyclosporine) to repress the immune responses. With immunosuppression (Fig. 1B), the mock transduction group still merely gave rise to a few osteoids at weeks 4 and 12, indicating a low degree of spontaneous osteogenic differentiation and bone formation without Bac-CB transduction. With the aid of immunosuppressive drugs, the transduction group led to evident bone spicule formation at weeks 4 and 12. Notably, the osteoblasts were localized to the surface of bone spicules (arrowheads in the insets) and the osteocytes resided within the lacuna (arrows in the insets), indicating the cell differentiation.
CT monitoring of calvarial bone repair
The mineralization and calvarial bone repair process were monitored by CT imaging (Fig. 2) and compared with the area of new bone within the defects (Table 1). Without immunosuppression (Fig. 2A), the control groups (scaffold only and mock transduction) only manifested bone formation near the periphery of the defect at week 4, and the bone formation barely improved at week 12. The transduction group resulted in the formation of bone islands in the central and peripheral areas of the defect at week 4, whose bone area (4.6 ± 1.2%) was approximately two times that of the control groups (2.4 ± 0.6% and 2.6 ± 0.7%). At week 12 more bone was formed near the edge of the defect, yet the bone area (7.3 ± 2.6%) was not significantly larger than that of the control groups (6.2 ± 2.3% and 7.3 ± 3.2%).
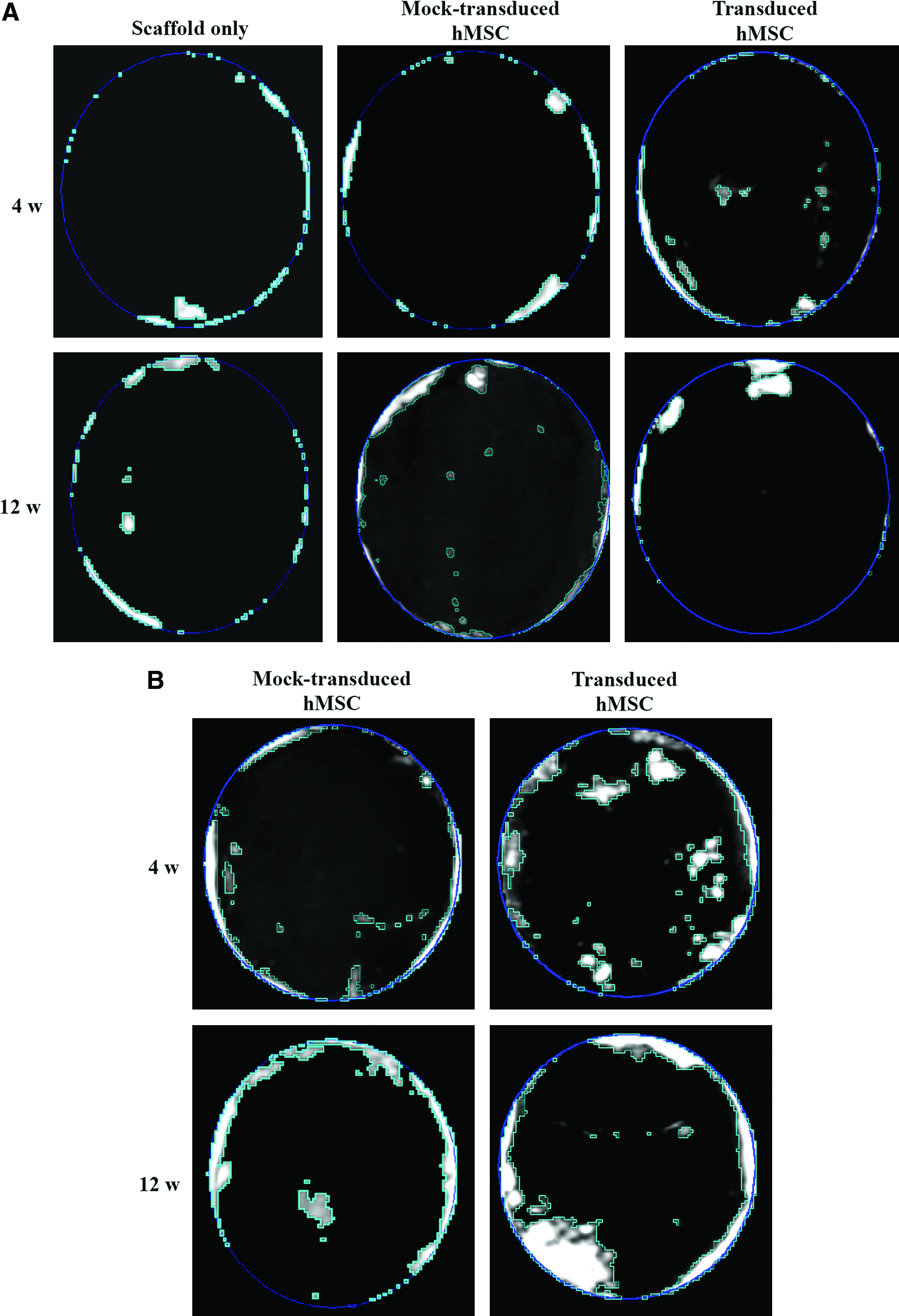
Computed tomography monitoring of bone repair in the absence (
n = 5 for each group at each time point.
hMSCs, human mesenchymal stem cells; N/D, not determined.
With immunosuppression (Fig. 2B), the mock-transduced hMSCs still failed to stimulate manifest new bone formation at weeks 4 and 12. Nevertheless, the transduced hMSCs gave rise to very evident formation of new bone that filled ≈ 14.7% and ≈ 28.1% of the defects at weeks 4 and 12, respectively (Fig. 2B and Table 1). The new bone was not completely bridged but the repair was significantly superior to those mediated by the mock transduction group with immunosuppression and the transduction group without immunosuppression.
Assessment of immune responses
Figures 1 and 2 collectively demonstrated that xenotransplantation of Bac-CB–engineered hMSCs into the defects was able to promote the calvarial bone repair in the immunocompetent rats, but the inferior bone repair in the absence of immunosuppression suggested that rejection responses retarded the bone-healing process. To confirm the roles of immune responses, the mock-transduced and transduced hMSCs were seeded to scaffolds and transplanted as in Figure 1, and the rats were bred without immunosuppressive drugs. The survival of the donor cells and the infiltration of host cells representative of innate immunity (macrophages) and adaptive immunity (CD3+ and CD8+ cells) were assessed by immunohistochemical staining at different time points.
Figure 3A (left column) illustrates the persistence and dispersion of numerous mock-transduced human cells within the graft at week 1, as stained by human mitochondrial antibody. In accord with this was that only a few macrophages and no T cells infiltrated into the graft, suggesting weak immune responses evoked by the mock-transduced, undifferentiated hMSCs. The transduced donor cells (right column, Fig. 3A) exhibited signs of osteogenic differentiation and formation of osteoids at week 1, as evidenced by the cell aggregation. Concomitant with the differentiation, markedly more macrophages and T cells (CD3+ and CD8+) invaded into the graft.
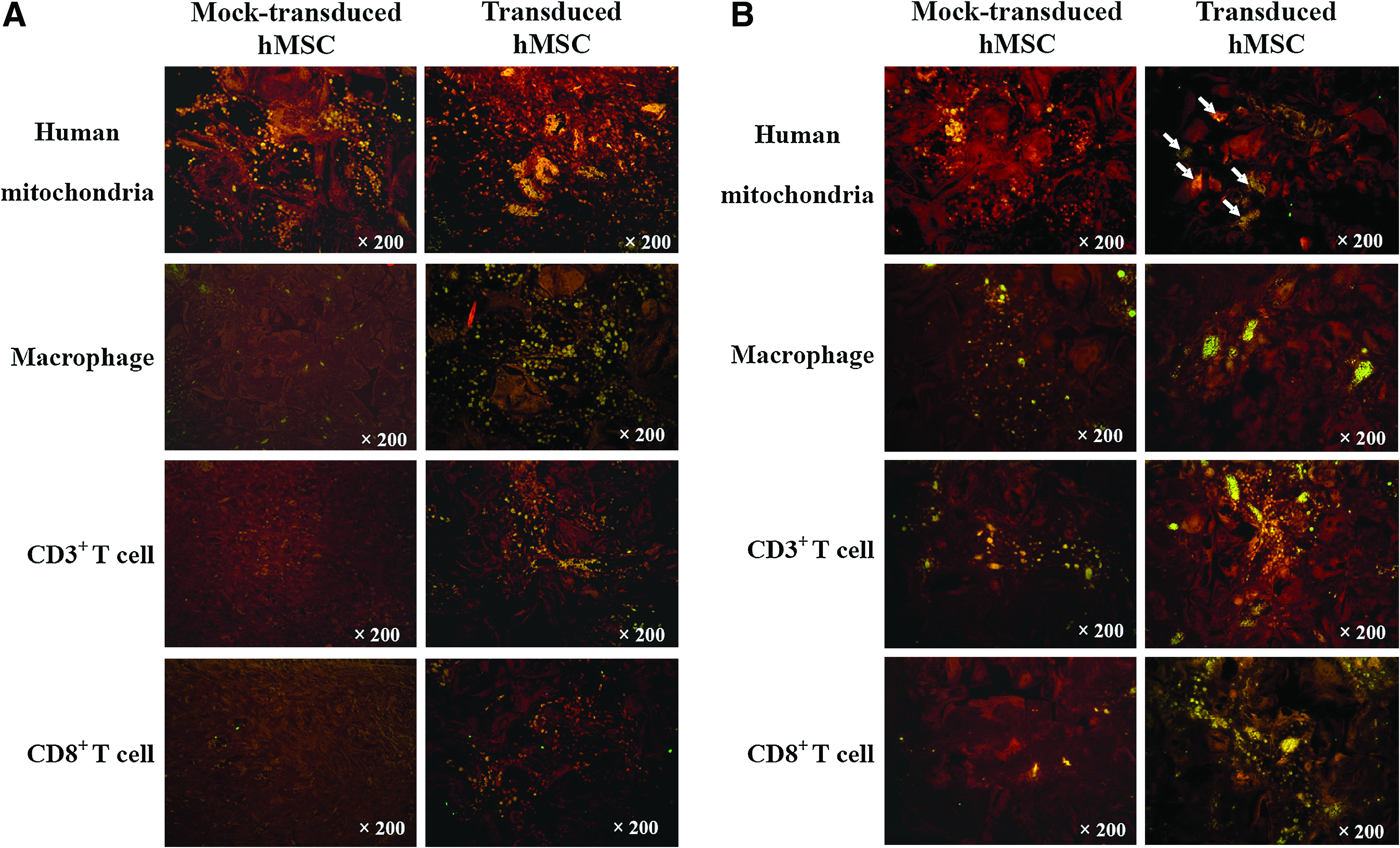
Survival of the transplanted cells and infiltration of macrophage and T cells at weeks 1 (
At week 4 (Fig. 3B), a number of mock-transduced human cells (left column) continued to persist, and some of them aggregated as clusters, indicating the onset of cell differentiation in the absence of Bac-CB transduction. Concurrently, macrophages, CD3+ T cells, and some CD8+ T cells infiltrated into the graft. Meanwhile, some of the transduced human cells remained detectable and colocalized with the bone matrix (indicated by arrows, Fig. 3B), suggesting the participation of the transplanted human cells in the new bone formation. When compared with the mock transduction group, the transduction was concomitant with more pronounced infiltration of macrophages and T cells (CD3+ and CD8+) into the bone matrix. These data suggested that the mock-transduced human cells were less susceptible to immunosurveillance than the transduced cells in the xenogeneic recipients.
hMSCs survival after xenotransplantation
To track the fate of the donor hMSCs in the long term, we performed the immunohistochemical staining specific for human cells at 8 and 12 weeks after transplantation. Without immunosuppression, significantly fewer mock-transduced cells were detectable at weeks 8 and 12 (Fig. 4A, left column) than at weeks 1 and 4 (Fig. 3). An even more dramatic decrease in cell number was observed in the transduction group at week 8, and the human cells completely vanished at week 12 (Fig. 4A, right column).

Survival of the transplanted cells in the absence (
In the animals receiving the immunosuppressive drugs (Fig. 4B), considerably more mock-transduced cells remained resident within the graft at weeks 8 and 12 than their counterparts without immunosuppression (Fig. 4A), proving the effect of the immunosuppressive drugs in prolonging the cell survival. Tremendously fewer cells in the transduction group remained viable at weeks 8 and 12, but the survived human cells surrounded the newly formed bone spicules (as marked by the dashed lines) in a way similar to the osteoblasts do upon normal bone formation. 27
Discussion
In this study we demonstrated that the mock-transduced hMSCs remained largely undifferentiated and stimulated poor bone matrix deposition and ossification, with or without immunosuppression (Figs. 1 and 2). In contrast, Bac-CB transduction considerably boosted the hMSCs aggregation (Figs. 1A and 3), ameliorated the accumulation of mineralized bone matrix (Fig. 1A), and initiated the bone island formation (Fig. 2A) at week 4 even without immunosuppression. With the aid of immunosuppression, the transduction group led to the regeneration of trabecular bone (Fig. 1B) that filled 28.1 ± 4.4% of the defect area at week 12 (Fig. 2B and Table 1), which significantly excelled as compared with the mock transduction group. Additionally, the embedding of differentiated donor cells within the bone matrix (Fig. 3) and cell lining the bone spicule (Fig. 4B) suggest that the Bac-CB–engineered hMSCs also responded to BMP-2 in an autocrine and/or paracrine fashion and participated in the new bone formation, a phenomenon that is also observed for bone formation from the muscle-derived stem cells. 28 These data confirmed that Bac-CB transduction directed the commitment of hMSCs along the osteogenic lineage and promoted the calvarial bone repair. This stimulatory effect is at least partly attributed to the efficient baculovirus transduction of hMSCs15,25 and ensuing BMP-2 expression to a level sufficient to induce bone regeneration. 17
Meanwhile, we detected the survival of numerous mock-transduced cells at 1 and 4 weeks after transplantation even without immunosuppression (Fig. 3). Although the number of engrafted cells remarkably decreased at weeks 8 and 12 (Fig. 4A), this suggested that the xenotransplanted hMSCs were fairly tolerated in the immunocompetent rats presumably, thanks to the immunosuppressive and immunoprivileged properties of the hMSCs. Alternatively, the survival might be ascribed to the protection of donor cells in the scaffold, which avoided prompt confrontation with the host immune cells. Moreover, many Bac-CB–transduced cells remained detectable in the graft at 1 week after transplantation, despite the onset of innate and adaptive immune responses (Fig. 3A). The mounting of cell-mediated immune responses concurred with the elimination of donor cells and inferior bone repair in case of no immunosuppression (Figs. 3B and 4A). The administration of immunosuppressive drugs considerably prolonged the cell survival to week 12 (Fig. 4B) and augmented the bone repair, which, however, was incomplete (Table 1).
The incomplete bone regeneration can be attributable to (1) the ineffectiveness of baculovirus-engineered hMSCs in orthotopic bone regeneration or (2) the immune responses. It has been shown that short-term BMP-2 expression is sufficient to irreversibly trigger bone formation in vivo.17,29 Further, Bac-CB transduction of MSCs derived from the bone marrow of New Zealand White (NZW) rabbits and subsequent allogeneic transplantation into a large critical-sized defect (8 mm in diameter and 10 mm in length) at the femora of NZW rabbits led to the segmental bone repair (manuscript in preparation). Since the femoral segmental model represents a more difficult bone-healing scenario, the success of Bac-CB in femoral bone healing confirmed that Bac-CB–engineered MSCs are capable of healing large bone defects and ruled out the first hypothesis. As such, the immune responses were primarily responsible for the incomplete bone healing.
Interestingly, the magnitude of immune responses appeared to correlate with Bac-CB transduction (Fig. 3), which can be accounted for by several factors. First, baculovirus transduction itself might mitigate the immunoprivileged properties of hMSCs, thereby exposing the transduced donor cells to immune surveillance. This assumption is unlikely because after xenotransplantation into Fisher 344 rats, the hMSCs that were transduced with a baculovirus expressing no transgene did not elicit considerably stronger macrophage and T-cell responses than the mock-transduced hMSCs (data not shown). Further, we recently demonstrated that hMSCs transduced with a baculovirus expressing no transgene retained the capability to suppress lymphocyte proliferation, and allotransplantation of baculovirus-transduced rat MSCs into Fisher 344 rats did not provoke apparent macrophage and T-cell responses. These data indicate that the baculovirus-transduced MSCs remained immunoprivileged in the allogeneic host 30 and excluded the first assumption. Second, the expression of human BMP-2 in the rats resulted in anti-BMP-2 antibodies and associated immune responses that eliminated the BMP-2–expressing cells. This is possible, but less likely to play a major role because secretion of BMPs alone does not adversely affect the cell survival in vivo. 31 Further, the successful segmental bone healing mediated by the Bac-CB–engineered rabbit MSCs in the immunocompetent NZW rabbits suggests that the anti-BMP-2 immunity (if any) does not explicitly impede the tissue regeneration.
The last and most likely possibility is that Bac-CB transduction accelerated the hMSCs differentiation, leading to the loss of immunoprivileged properties and hence depletion by the intrinsic immune systems of xenogeneic rats. This assumption is in line with the recent finding that the immunosuppressive potential of mouse MSCs is lost after transplantation, and hence the cells are rejected. 32 Moreover, Bac-CB transduction accelerated the bone formation, which is in nature accompanied by neovascularization. The formation of vasculature might increase the flux of blood, facilitate the recruitment of immune cells into the transplantation site, 20 and hence lead to a more serious graft rejection response than the mock transduction group. Further experiments to scrutinize these hypotheses are ongoing.
In conclusion, in this study we demonstrated that Bac-CB–engineered hMSCs remarkably promoted the calvarial bone regeneration in the immunocompetent rats, implicating the potential of baculovirus in MSCs engineering and tissue repair. Our data supported the notion that the xenotransplanted hMSCs are able to evade the immune surveillance initially (e.g., at 1 week after transplantation), but are eventually rejected even with immunosuppression. Although administration of immunosuppressive drugs prolonged the survival and augmented the bone healing, the immunosuppression was unable to entirely abolish the immune response, leading to the incomplete bone healing. These results partly concurred with recent reports that stem cells xenotransplanted into the femoral defect, 33 intervertebral discs, 34 or posterolateral lumbar spine 35 are able to survive for a long term (up to 6 months) and contribute to new tissue formation (e.g., bone). However, these studies did not report cell clearance as a result of immunological rejection, probably because the animal models were immunodeficient 33 or the transplantation sites (e.g., intervertebral discs) are relatively immunoprivileged. 24 Despite cell survival, no definitive, complete tissue healing was reported in these studies. Therefore, the use of hMSCs appears to be restricted to autologous or allogeneic transplantation.
Footnotes
Acknowledgments
The authors acknowledge the financial support from the National Tsing Hua University Booster Program (97N2511E1, 98N2901E1), National Tsing Hua University-Chang Gung Memorial Hospital Joint Research Program (CMRPG380101), National Science Council (NSC 97-2627-B-007-014), and National Health Research Institutes (NHRI-EX97-9412EI), Taiwan.
Disclosure Statement
No competing financial interests exist.