
Abstract
Epigallocatechin-3-O-gallate (EGCG) is known to have beneficial effects on the nonfrozen preservation of mammalian cells and tissues. In this study, we aimed at testifying the hypothesis that the deleterious effects of cold preservation of articular cartilages can be ameliorated by the addition of EGCG to the storage media. Articular cartilages were preserved in a storage solution composed of serum-free RPMI 1640 media with 1 mM EGCG at 4°C for 1–4 weeks. The regulatory effects of EGCG on cell cycle progression as well as expression levels of CyCliNS (CCNs) and nuclear factor-κB (NF-κB) were investigated in articular chondrocytes. Chondrocyte viability of cartilages preserved with EGCG was significantly well maintained for 2 weeks with high contents of glycosaminoglycan and total collagen. These beneficial effects of EGCG were confirmed by histological and immunohistochemical observations showing well-preserved cartilaginous structures and delayed denaturation of extracellular matrices. The compressive elastic modulus of cartilages preserved with EGCG was close to that of fresh specimens. Increased cell population at the G0/G1 phase by EGCG returned to the normal level after EGCG removal, whereas decrease at the G2/M phase did not. Negatively regulated expression of CCND1, CCNE2, or NF-κB in EGCG-treated cells was restored by removing EGCG, but not CCNA2 and CCNB1. After 8 weeks of in vivo implantation into full-thickness cartilage defects in rabbits, the cartilages preserved with EGCG were found to be integrated with the host environment and support tissue regeneration. It is suggested that EGCG plays effective roles in preserving and repairing articular cartilages by reversibly regulating cell cycle at G0/G1 phase and NF-κB expression.
Introduction
A good storage solution should prevent this peroxy lipid generation. Since this is related to cell proliferation and division, longer term tissue preservation for transplantation would become possible if cellular metabolism can be controlled. 9 From this point of view, our study focused on (–)-epigallocatechin-3-O-gallate (EGCG), the predominant catechin from tea, because it has a wide range of pharmacological activities, including antioxidant, anticancer, antiproliferative, antiinflammatory, and antimicrobial effects.10–12 Different from these biological activities of EGCG, its beneficial preservative effects on mammalian cells and tissues were also examined to design a cell- or tissue-preserving medium/solution at physiological temperature in our previous studies.13–15 A fresh osteochondral allograft is one of the most effective treatments for cartilage defects of the knee. Despite the clinical success, fresh osteochondral allografts have great limitations in relation to the short storage time that cartilage tissues can be well preserved. In the clinical use of fresh osteochondral allografts, the paradigm is to rapidly transplant harvested tissue to ensure delivery of a high level of viable chondrocytes. 16 The published clinical studies reflect the situation where grafts were harvested, stored in lactated Ringer's solution at 4°C, and transplanted between 2 and 5 days from the time of harvest.17,18 However, although the viability of cold-preserved articular cartilage is significantly diminished within 1 week,19–21 appropriate serology testing to minimize the chances for the disease transmission requires a minimum of 2 weeks. 16
In this study, a storage solution containing EGCG was used to test the hypothesis that cold preservation injury of articular cartilages can be prevented by EGCG-mediated reversible regulation of cell cycle. Moreover, the regulatory effects of EGCG on cell cycle progression as well as expression levels of CyCliNS (CCNs) and nuclear factor-κB (NF-κB) were investigated in human articular chondrocytes (hACs). Cartilage defects with full thickness were created surgically on the femoropatellar groove of the knee joints of rabbits. Cartilage specimens preserved without or with EGCG were transplanted in rabbits using a press-fit method. Rabbits were sacrificed at 4 and 8 weeks after surgery, and the repaired tissues were assessed both macroscopically and histologically. Our data provided support to a scenario in which EGCG might play a key role in preserving articular cartilages by reversibly regulating CCNs and NF-κB expression. Therefore, EGCG-based storage solution could be exploited to craft strategies for the long-term preservation of osteochondral allografts under cold storage conditions.
Materials and Methods
Cartilage specimen collection and cold preservation
Discarded articular cartilages were obtained from the knee joints of 10 patients aged 58–86 years (mean age, 72 ± 8.3 years; seven Japanese women and three Japanese men) who underwent total knee arthroplasty at Marunouchi Hospital, Matsumoto, Japan. These normal cartilage specimens (15–20 mm in diameter and 2–2.5 mm in thickness) were procured by osteotomy from the donors under sterile conditions and placed in saline for 0.5–1 h until the end of surgery. Immediately after surgery, the specimens were transferred in a storage solution (serum-free RPMI 1640 media [Sigma–Aldrich, St. Louis, MO] with 1% antibiotic–antimycotic solution [including 10,000 U penicillin, 10 mg streptomycin, and 25 μg amphotericin B/mL; Sigma−Aldrich]) without or with 1 mM EGCG (Teavigo™; DSM Nutritional Products, Basel, Switzerland) and kept at 4°C. The specimens were then delivered to the senior author (Prof. Hyon) within 1 day from procurement. Additionally, normal fresh cartilages were delivered in a complete media with 10% fetal bovine serum (Sigma − Aldrich) at room temperature after procurement from the donor. Because of this necessary processing delay between tissue procurements from the donor and its delivery, 1 day was set as the data point for the fresh specimen.
On receipt of the cartilage tissues, the specimens were replaced with either 20 mL of a storage solution without or with EGCG and then stored at 4°C for 1, 2, and 4 weeks without changing the storage solution. At the end of each storage period, biochemical, immunohistochemical, and biomechanical analyses were performed as described below. All procedures involving human subjects received prior approval from Marunouchi Hospital, Osaka City University Graduate School of Medicine and the Institutional Review Board of Institute for Frontier Medical Sciences, Kyoto University, and all subjects provided written informed consent.
Chondrocyte viability
The number of viable cells was quantified indirectly using a highly water-soluble tetrazolium salt [WST-8, 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfo-phenyl)-2H-tetrazolium, monosodium salt] (Dojindo Laboratory, Kumamoto, Japan), reduced to formazan dye by mitochondrial dehydrogenase. The viability of chondrocytes in the cartilages was found to be directly proportional to the metabolic reaction products obtained in WST-8. According to the manufacturer's instructions, the specimens following cold preservation were incubated with WST-8 in the last 4 h of the incubation period (24 h) at 37°C in the dark. Normal fresh cartilages were regarded as the controls. The absorbance was determined at 450 nm in a microplate reader (VersaMax™; Molecular Device, Sunnyvale, CA).
Glycosaminoglycan and collagen contents
Glycosaminoglycan (GAG) contents in the cartilages were determined using a dimethylmethylene blue dye–binding assay kit (Blyscan kit; Biocolor Ltd., Newtownabbey, Northern Ireland) according to the manufacturer's instructions. Chondroitin sulfate, provided with the kit, was used as the GAG standard. Briefly, 50 mg of cartilage was homogenized and solubilized. A 100 μL aliquot from each homogenate was added to 1 mL of dimethylmethylene blue and mixed periodically at 25°C for 30 min. Solutions were then centrifuged at 10,000 g for 10 min, and the supernatants were discarded. The remaining pellet was suspended in 1 mL of the provided dissociation reagent, and absorbance measurements were taken at 656 nm.
Total cartilage collagen levels in the cartilages were determined using a Sircol collagen dye–binding assay kit (Biocolor Ltd.) according to the manufacturer's instructions. Briefly, 50 mg of cartilage was homogenized, and total acid pepsin-soluble collagens were extracted overnight using 5 mg/mL pepsin in 500 μL of 0.5 M acetic acid. One microliter of Sircol dye reagent was added to 100 μL of each homogenate and incubated at 25°C for 30 min. After centrifugation, the pellet was suspended in 1 mL of alkaline reagent, and absorbance was read at 540 nm.
Histological and immunohistochemical analyses
At the completion of the predetermined preservation and incubation, each cartilage specimen was rinsed with phosphate-buffered saline (PBS; pH 7.4) and immediately fixed with 2% glutaraldehyde, 2% paraformaldehyde, and 0.2% CaCl2, followed by embedding in paraffin. The tissue blocks were sectioned by 5 μm thickness, stained with either hematoxylin and eosin (for general evaluation) or Safranin-O/fast green (for GAG content and distribution), and immunostained with rabbit monoclonal antibody (Ab) against type II collagen. The prepared sections were examined using an optical/fluorescence microscope (Biozero–8000; Keyence, Osaka, Japan).
Biomechanical analysis
At each data point, cartilage specimens were removed from storage solutions and placed immediately in a −80°C freezer until biomechanical testing was completed. At the time of testing, each specimen was thawed at room temperature (25°C) in PBS containing proteinase inhibitors. A cartilage compression disk with an intact articular surface measuring 10 mm in diameter and 1 mm in thickness was made from each specimen. To determine the compressive elastic modulus, this cartilage disk was then subjected to dynamic viscoelastic compression (Rheogel E-4000; UBM, Kyoto, Japan) with 0.5% strain. All data were collected at a frequency of 10 Hz.
Cell culture and EGCG treatment
The macroscopic cartilage degeneration was determined by staining femoral head specimens with India ink, 22 and fresh articular cartilage with smooth articular surface (unaffected cartilage) was resected and used to prepare hACs by enzymatic digestion as previously described. 23 Briefly, the harvested cartilages were minced separately using a scalpel and digested overnight in 2 mg/mL type II collagenase (Sigma−Aldrich). The resulting cell suspensions were precipitated, and then the supernatants were removed. The pellets were soaked in Dulbecco's modified Eagle's medium/Ham's F-12 (Sigma–Aldrich), supplemented with 10% fetal bovine serum (Sigma–Aldrich) and 1% antibiotic–antimycotic solution at 37°C in a humidified atmosphere supplemented with 5% CO2. Isolated hACs were plated at a density of 1 × 106/mL in 35-mm tissue culture dishes (Corning, NY). In some cases, the cells were passaged once to obtain sufficient cell numbers before usage.
To examine the effects of EGCG on cell cycle progression as well as expression of cyclins (CCNs) and NF-κB of hACs, the cells were seeded onto 48-well plates and then incubated with different treatment methods of EGCG as follows: in treatment 1 (E-Tr1), the cells were incubated with 25 μM EGCG for 1 day; in treatment 2 (E-Tr2), the cells were incubated with 25 μM for 1 day, washed five times with PBS, followed by replacement with fresh media, and then incubated for additional 2 days; in treatment 3 (E-Tr3), the cells were treated like E-Tr2 and then incubated for additional 8 days. As the nontreated control (E-Tr0), the cells were incubated without any treatment.
Cell cycle assay
For cell cycle analysis, hACs were differently treated with EGCG (E-Tr1–E-Tr3), collected by trypsinization, and washed with cold PBS (pH 7.2). The cells were resuspended in 95% cold methanol for 1 h at 4°C and then centrifuged at 250 g for 5 min. The pellet was washed twice with cold PBS and incubated with RNase A (20 U/mL, final concentration; Sigma–Aldrich) at 37°C for 30 min. Afterward, the cells were chilled over ice for 10 min and stained with propidium iodide (100 μg/mL; Sigma–Aldrich) for 1 h. No less than 20,000 cells were counted using a fluorescence-activated cell sorter (FACSCan™ System; BD Biosciences, San Jose, CA). The data obtained were analyzed using the histogram of CellQuest© software, written by Mac-App (BD Biosciences).
Western blot analysis
Following different EGCG treatments (E-Tr1–E-Tr3) to hACs, the cells were washed twice with cold PBS (10 mM, pH 7.4). Ice-cold RIPA lysis buffer (Santa Cruz Biotechnology, Santa Cruz, CA) was added to the cells for 5 min. The cells were scraped, and the lysate was cleared by centrifugation at 14,000 g for 20 min at 4°C. The resultant supernatant (total cell lysate) was used. The protein concentration was determined by a BCA™ protein assay (Pierce, Rockford, IL). For immunoblot analysis, 35–40 μg of protein was run on 4/20 polyacrylamide–sodium dodecyl sulfate gel (Daiichi Pure Chemicals, Tokyo, Japan) for 1 h at 30 mA and blotted to a polyvinylidene difluoride (PVDF) membrane for 50 min at 35 mA. The membrane was blocked in a blocking buffer (Nacalai Tesque, Kyoto, Japan) for 1 h at room temperature and then probed overnight with primary Ab. After incubation with horseradish peroxidase–conjugated secondary Ab (1:5000 dilution, anti-rabbit IgG from Santa Cruz Biotechnology; or 1:2000 dilution, anti-mouse IgG from Amersham Biosciences [Buckinghamshire, United Kingdom]), protein expression was radiated by a Chemilumi-one chemiluminescent kit (Nacalai Tesque) and detected with an X-ray film (Fujifilm, Tokyo, Japan). Densitometric analyses were performed using a Video-Imager (Bio-Rad Laboratories, Hercules, CA). Immunoblotting was performed using Abs directed against the following antigens: NF-κB (rabbit polyclonal Ab) used in a 1:200 dilution from Santa Cruz Biotechnology; CCNA2 (mouse IgG1), CCNB1 (mouse IgG2a), and CCND1 (mouse IgG1) at 1:250–500 dilution from BD Biosciences, CCNE2 (rabbit polyclonal Ab) at 1:1000 dilution from Cell Signaling Technology (Danvers, MA); β-actin (rabbit polyclonal Ab) at 1:5000 dilution from Novus Biologicals (Littleton, CO); and glyceraldehyde-3-phosphate dehydrogenase (mouse monoclonal Ab) at 1:5000 dilution from Chemicon International (Temecula, CA).
Statistical analysis
All variables were tested in three independent storages or cultures for each experiment, and each experiment was repeated twice (n = 6). Quantitative data were expressed as mean ± standard deviation. Statistical comparisons were carried out with a one-way analysis of variance (SAS Institute, Cary, NC), which was followed by the Bonferroni test for the multiple comparisons. A value of p < 0.05 was considered statistically significant.
Results
Effect of EGCG on chondrocyte viability of cartilages
Preserving the cartilage specimens without EGCG resulted in a significant (p < 0.05) time-dependent decrease in the cell viability (Fig. 1). Their viability was already at most 59% after 1 week, which was reduced to approximately 50% after 2 weeks and 28% after 4 weeks (p < 0.05). When the specimens were preserved with 1 mM EGCG, their viability loss was only 23% even after 2 weeks, indicating that EGCG suppresses the time-dependent cellular viability reduction. Although this decrease in the viability was accelerated with the progress in the incubation time, there was a significant (p < 0.05) difference between cartilages preserved without and with EGCG after 2 weeks. However, the protective activity by EGCG was relatively less exerted after 4 weeks, showing that the viability was reduced to about 50%.
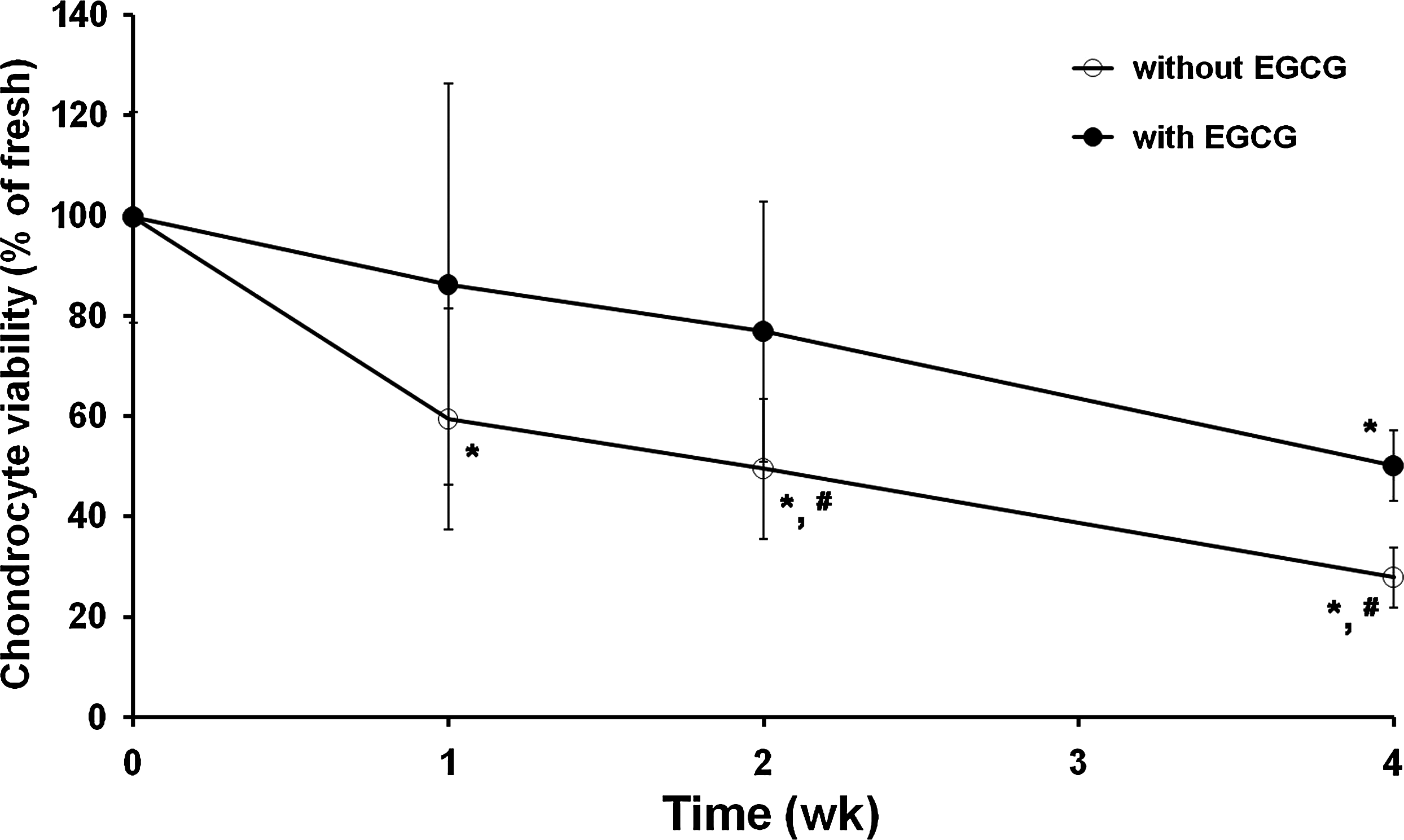
Chondrocyte viability in cartilages preserved without or with epigallocatechin-3-O-gallate (EGCG). Cell viability was measured with a WST-8 assay as described in the “Materials and Methods” section. The results are reported as mean ± standard deviation (SD) (n = 6) and analyzed by the Bonferroni test (*p < 0.05 vs. fresh control and #p < 0.05 vs. with EGCG at each time point).
Effect of EGCG on contents of GAG and collagen in cartilages
After 1 week of preservation without EGCG, GAG content was not more than 67% (Fig. 2A). With increase in the preservation time, it declined to about 57% at 2 weeks and then decreased dramatically to only 39% at 4 weeks. Storage with EGCG provided the improved preservation of chondrocyte metabolism, as compared with storage without EGCG. After 2 weeks of preservation with EGCG, GAG content was measured to be 73%, which was significantly (p < 0.05) higher than that obtained from preservation without EGCG. After 4 weeks of storage with EGCG, the content significantly (p < 0.05) decreased to 64% compared with the fresh. However, this was still significantly (p < 0.05) higher than storage without EGCG for the same duration. Collagen contents showed a similar pattern to those of GAG, but there was no significant difference between preservation without and with EGCG (Fig. 2B). These results imply that GAG and collagen contents of the cartilages preserved without or with EGCG correlate well with the cell viability of the corresponding cartilages (Figs. 2C, 2D).

Extracellular matrix (ECM) composition of cartilages preserved without or with EGCG. Glycosaminoglycan (GAG) contents (
Effect of EGCG on matrix structure of cartilages
Routine histological and immunohistochemical examinations were performed on cartilage specimens preserved without or with EGCG (Fig. 3). Through the hematoxylin and eosin staining, it was difficult to differentiate between cartilages preserved without and with EGCG (Fig. 3A). However, there was a numerical trend favoring specimens stored with EGCG for 2 weeks. The specimens stored with EGCG stained positively with Safranin-O, showing abundant presence of GAG, which was comparable to that of the fresh ones (Fig. 3B). In contrast, preservation without EGCG resulted in an appreciable decrease in the positivity for Safranin-O. Overall, the outside region of each cartilage had a propensity to have higher cell density and to be stained more intensely for GAG than the central region. The cells in the lacunas of fresh and EGCG-preserved cartilages were well maintained with an original chondrocytic phenotype compared with those preserved without EGCG. Immunohistochemical examination with monoclonal Ab against type II collagen showed the presence of this macromolecule in the cartilages preserved with EGCG, and again, the cell density and staining intensity were greatest in the periphery of the specimens (Fig. 3C). In each of the specimens examined, type II collagen primarily was found intercellularly. Comparison of cartilage tissues that stained positive for both GAG and type II collagen within the same specimen revealed that Safranin-O–positive tissues had the usual association with type II collagen typically seen in normal articular cartilage.
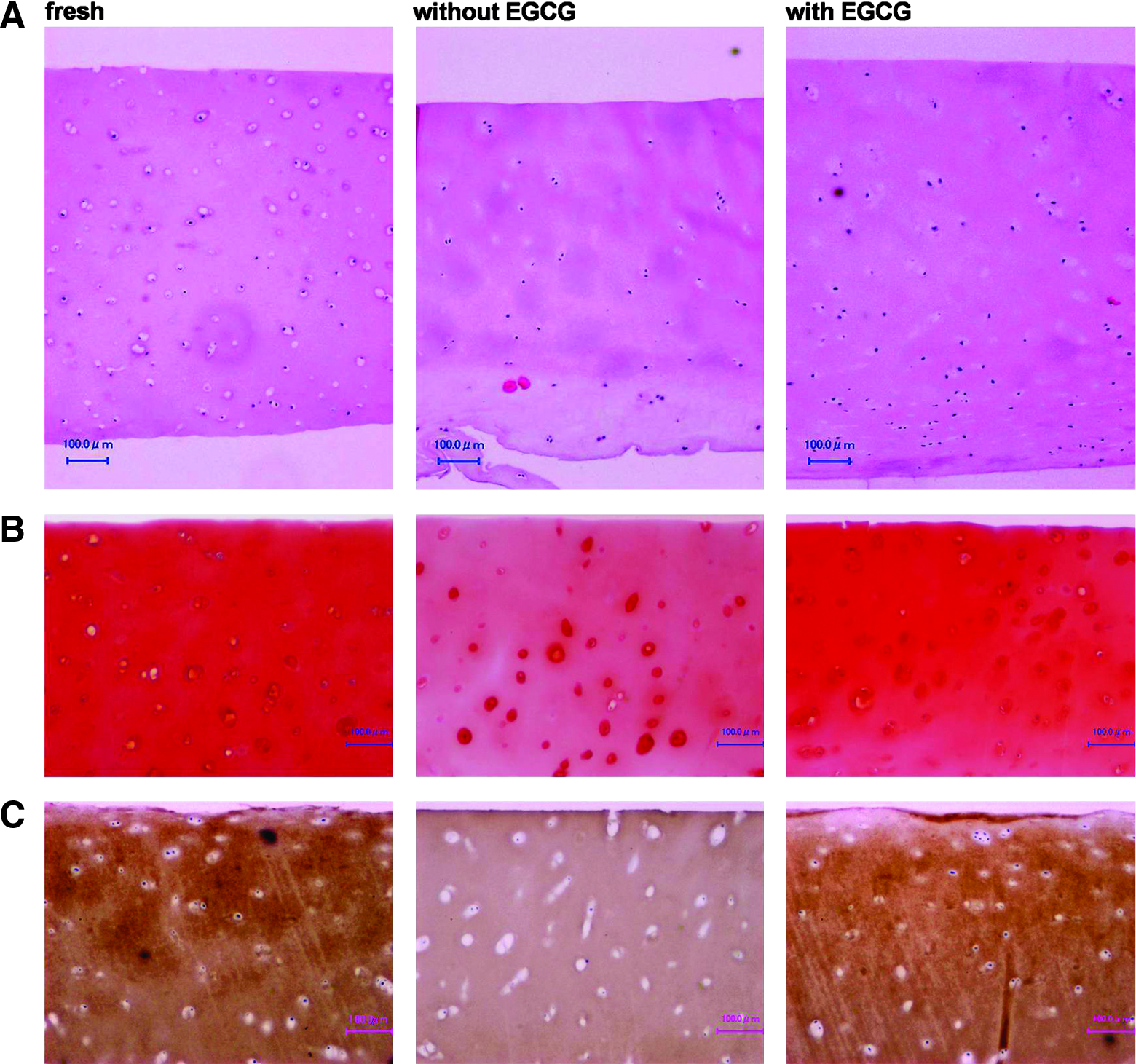
ECM structure of cartilages preserved without or with EGCG. Cartilage specimens were stained with either hematoxylin and eosin (
Effect of EGCG on biomechanical property of cartilages
The result of mechanical evaluation showing the functional property of the cartilage matrix was opposed to that of the biochemical assay for GAG and collagen contents. The compression elastic moduli of cartilages preserved without and with EGCG were increased with time (Fig. 4). Fresh cartilage specimens showed an initial modulus of 206.7 ± 39.1 kPa. Cartilages preserved without and with EGCG for 1 week had moduli of 287.2 ± 46.0 and 218.2 ± 45.0 kPa, respectively, close to that of the fresh cartilage. After 1 week, after storage without and with EGCG for 2 weeks, the moduli went up to 345.9 ± 93.9 and 302.2 ± 123.8 kPa, respectively. There were no significant changes in the mechanical strength, if any, slightly increased. However, the increasing rate steeply rose after 4 weeks irrespective of EGCG addition. Cartilages preserved without and with EGCG had moduli of 702.7 ± 144.6 and 458.0 ± 69.0 kPa, respectively, 340% and 222% of the normal value.
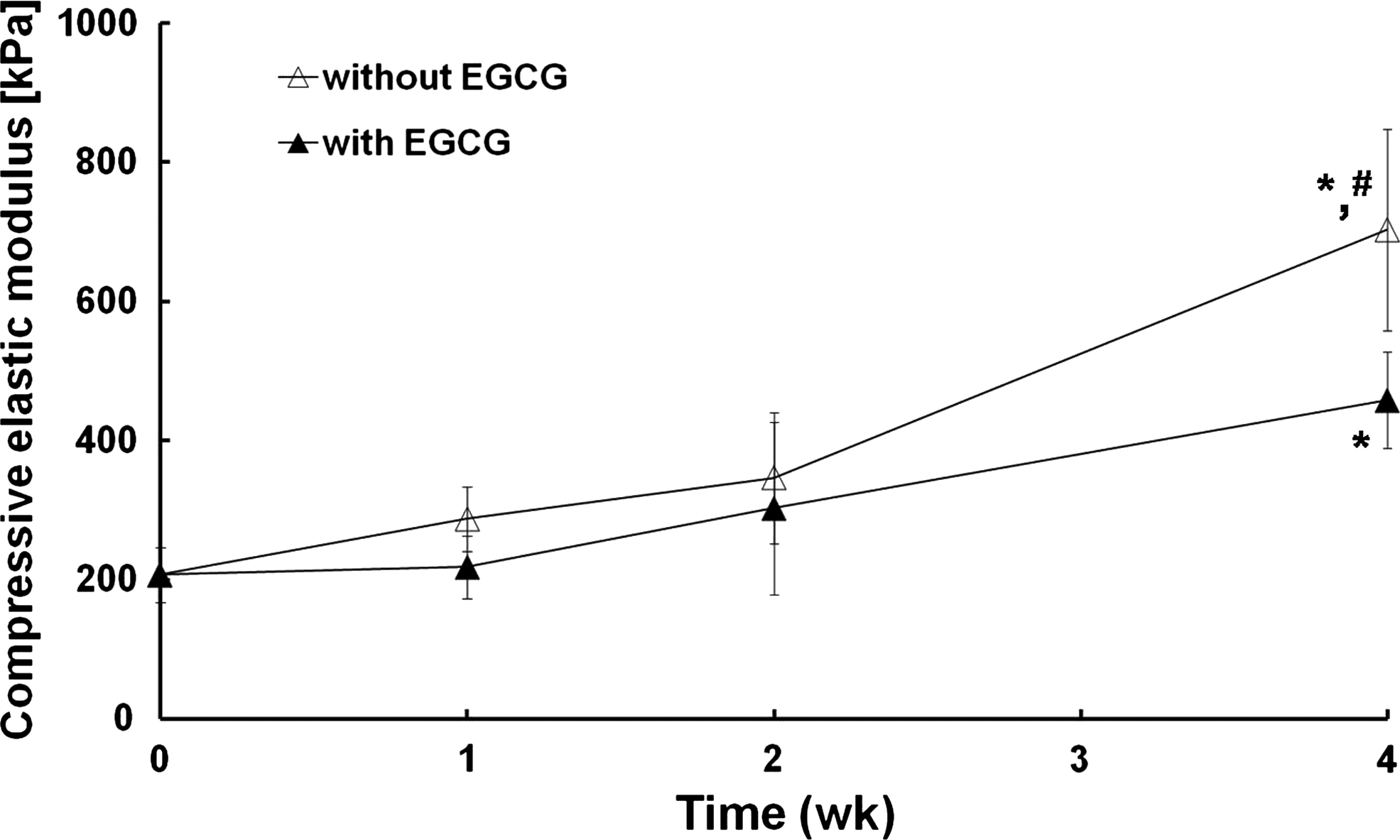
Biomechanical property of cartilages preserved without or with EGCG. Compressive elastic modulus was determined by dynamic viscoelastic compression as described in the “Materials and Methods” section. The results are reported as mean ± SD (n = 6) and analyzed by the Bonferroni test (*p < 0.05 vs. fresh control and #p < 0.05 vs. with EGCG at 4 weeks).
Effect of EGCG on cell cycle progression of chondrocytes
Cytometric analysis showed that EGCG treatment to hACs changed the profile of their DNA content (Fig. 5). The cell cycle of the cells treated with EGCG for 1 day (E-Tr1) was significantly (p < 0.05) delayed at G0/G1 phase compared with that of the nontreated control. In E-Tr2, the cell population at S phase was completely recovered after 2 days of EGCG removal from the culture media, but that at G0/G1 and G2/M phases was not. After 8 days of EGCG removal (E-Tr3), the population of cells at G0/G1 phase was restored near the level of control cells, but that at G2/M phase was kept at slightly lower level. These results mean that EGCG treatment resulted in significant (p < 0.05) increase of the cell population at G0/G1 phase with concomitant decrease at S and G2/M phases, but EGCG removal resulted in a return to normal levels except for those in the G2/M phase.

Effect of EGCG on cell cycle progression of human articular chondrocytes (hACs). EGCG was differently treated with cells (E-Tr0, nontreated control; E-Tr1, incubation with 25 μg/mL EGCG for 1 day; E-Tr2, incubation with 25 μM EGCG for 1 day, washing with phosphate-buffered saline and further incubation for 2 days; E-Tr3, E-Tr2 + additional 6 days incubation). Cell cycle was determined with a cytometric analysis, as described in the “Materials and Methods” section. The results are reported as mean ± SD (n = 6) and analyzed by the Bonferroni test (*p < 0.05 vs. E-Tr0 and #p < 0.05 vs. E-Tr1).
Effect of EGCG on CCNS and NF-κB expression in chondrocytes
Figure 6 demonstrates the regulating effects of EGCG on the expression level of CCNs and NF-κB proteins in hACs. The protein expression levels of CCND1 and CCNE2, which regulate the G1 phase and G1/S phase transition, respectively, were significantly (p < 0.05) decreased by EGCG treatment (E-Tr1), but they were restored after 2 days of EGCG removal (E-Tr2) (Fig. 6A). The expression level of CCNA2 and CCNB1, which mainly regulate the G2/M phase transition and M phase, respectively, was similarly decreased after EGCG treatment and remained at low levels even 2 days after the removal of EGCG (Fig. 6B). These results were well agreed with those of the cell cycle profile, showing that the cell cycle of EGCG-treated cells for 1 day was delayed at G0/G1 phase, and it was recovered after EGCG removal.

Effect of EGCG on CCNs (
To evaluate the involvement of NF-κB as the mechanism of EGCG-mediated reversible regulation of the cell cycle in hACs, antibodies directed against the p65 and p50 subunits of NF-κB were used (Fig. 6C). The expression of NF-κB/p65 protein was significantly (p < 0.05) suppressed by the treatment of EGCG for 1 day (E-Tr1), but its expression level completely was restored after 2 days of EGCG removal. The expression of NF-κB/p50 showed an almost similar pattern to that of NF-κB/p65. These results suggest that either NF-κB activation or translocation in the cells may be temporarily blocked by EGCG, but can be reversed immediately after EGCG removal.
Discussion
In this study, we found that the cold preservation injury of articular cartilages could be prevented by EGCG-mediated reversible regulation of cell cycle. By EGCG addition to a storage solution, cartilage tissues were well preserved for at least 2 weeks with high chondrocyte viability, high contents of GAG and total collagen, histological and immunohistochemical appearance of well-preserved cartilaginous structures, delayed denaturation of extracellular matrix (ECM), and slightly increased compressive elastic modulus (Figs. 1–4). After 8 weeks of in vivo implantation, the cartilages preserved with EGCG were found to be integrated with the host environment and support tissue regeneration showing histologically good subchondral bone connection and better surrounding cartilage infusion (Supplemental Fig. S1, available online at www.liebertonline.com/ten). Owing to added EGCG in the storage solution, better structural integrity and more maintenance of ECM were achieved in the specimens preserved with EGCG. These results can be attributed to the fact that EGCG might actively penetrate into the cartilaginous layer and protect the chondrocyte viability and metabolism in cartilages as well as the matrix structure.
Histologically, metachromatic staining of cartilage matrix preserved in a storage media without EGCG for 2 weeks was lost mainly from perichondrocyte area, suggesting that chondrocytes that lost their viability digested matrix by themselves. Some of the chondrocytes positively stained with Safranin-O in their pericellular region indicated the proteolysis of cartilage matrix. It was found that EGCG conjugated with fluorescein isothiocyanate penetrated into the matrices of cartilages and was incorporated into the cytosol of cells (Supplemental Fig. S2, available online at www.liebertonline.com/ten), enabling cartilaginous structures of preserved cartilages to be well preserved and allowing denaturation of ECM to be delayed. Because the viability of chondrocytes in cartilage was kept high at least 2 weeks, this procedure may be effective not only in preserving the allogeneic osteochondral graft but also chondrocyte transplantation, such as autologous chondrocyte implantation and allogeneic chondrocyte transplantation. In case of chondrocyte transplantation, we have to harvest cartilage and carry it to cell processing center, which takes a few hours to a couple of days. Because maintaining cell viability is highly important in cell transplantation, 24 this procedure may be useful in carrying cartilage.
A point to be solved is that there was a time-dependent decrease in the ECM contents and the compressive elastic modulus with the increase in the storage time. These findings correlated highly with chondrocyte viability. Mechanically, compressive elastic moduli were analyzed to compare between the cartilages preserved without and with EGCG and to show the gradual approaching of the repaired defect to normal tissue. After 4 weeks of preservation, the cartilage preserved without EGCG had a significantly higher modulus than that preserved with EGCG. In contrast, a recent report has demonstrated that the storage duration of human osteochondral grafts in serum-free Dulbecco's modified Eagle's medium/Ham's F-12 at 4°C may be extended to approximately 2 weeks with no significant detrimental changes to the biochemical or biomechanical properties of the tissue. 25 There are a number of explanations for these inconsistencies, including difference in the determination of cell viability and ECM contents as well as the measurement of compressive elastic modulus.
In this study, a storage solution containing EGCG was used to testify the hypothesis that the deleterious effects of cold preservation of osteochondral allografts can be ameliorated by the addition of EGCG to the storage media. EGCG has been shown to have suppressive effects on various cancer cells by inducing cell cycle arrest,26,27 whereas less attention has been paid to its beneficial effects on nonfrozen preservation of mammalian cells and tissues through regulating cell cycle. To our knowledge, however, little has been published on the beneficial effects of EGCG on the prolonged cold storage of human osteochondral grafts and the mechanism at cellular level. The cell cycle–related proteins, CCNs, were suppressed during the period when hACs were kept in a medium containing EGCG, but once EGCG was removed from the medium, the protein expression was restored to the original levels. This reversible cell cycle regulation showed a temporal correlation with the results in the cell cycle assay. The cell cycle profiles of EGCG-treated cells showed the increase of G0/G1 phase and the suppression of S and G2/M phases, indicating that EGCG delays the on-going cell cycle at G1 phase and prevents nuclear replication (Fig. 5). Removal of EGCG from the culture medium restored the profile of DNA contents, except for G2/M phase, similar to the one before EGCG treatment or the control, which suggests that the effect of EGCG on cell cycle progression is reversible. Our hypothesis was further validated by finding positive correlations with the results from immunoblotting. The results of immunoblotting showed similarity to those of the cell cycle profiles (Fig. 6). It is presumed that factors related to the translation of CCNA or CCNB in RNA were inhibited even after the removal of EGCG, but that of CCND or CCNE were not. It was also shown that the expression of NF-κB in hACs was negatively affected by EGCG treatment, but recovered after EGCG removal. The results of this study demonstrated that EGCG induces cellular hibernation through a temporary delay of the cell cycle, as shown previously, 28 which can be reversibly resumed after the removal of EGCG from the culture medium. The reversible effect of EGCG on the expression levels of cell cycle–related proteins suggests that EGCG may be a hibernation-inducing agent that allows a potential recovery of physiological functions and structural elements essential for cell and tissue preservation.
The cellular uptake of EGCG treated with hACs was clearly observed (Supplemental Fig. S3, available online at www.liebertonline.com/ten). It is not dangerous to conclude that these phenomena could be related to the intrinsic characteristics of polyphenolic compound, EGCG. It has already been known that this compound, because of its amphipathic properties, easily binds to ECM, phospholipid bilayered membranes, and any types of intracellular proteins.29–31 Therefore, it seems that this compound enables to be internalized into cell cytosol and further to be translocated into nucleus, which leads to modulating the exogenous signals directed to genes required for the survival and apoptosis of cells. Although exact mechanism of the internalization of EGCG into cells had not been elucidated yet, some evidence was reported showing that EGCG was bound to the membrane as well as incorporated into the cytosol and the nucleus of cancer cell lines, such as PC-9 and HT-29 cells.32,33 Consistently, recent study has demonstrated that expression of the metastasis-associated 67 kDa laminin receptor might confer EGCG responsiveness to cancer cells at physiologically relevant concentrations, suggesting that the gallate moiety of EGCG may be critical for receptor binding and subsequent activity.34,35 Moreover, the observation that nucleic acids extracted from catechin-treated cells were colored implied that because both galloyl and catechol groups of EGCG were essential for DNA binding, both groups seemed to hold strands of DNA via their branching structure. 36
Taking these results into consideration, it is concluded that EGCG plays effective roles in preserving articular cartilages by reversibly regulating cell cycle at G0/G1 phase and NF-κB expression, which is regarded as a feasible action mechanism of EGCG (Supplemental Fig. S4, available online at www.liebertonline.com). Thus, it is suggested that these results can be exploited to craft strategies for the long-term preservation of osteochondral allografts under cold storage conditions.
Footnotes
Acknowledgment
This work was supported by the Bio-Scientific Research Grant funded by the Pusan National University (PNU, Bio-Scientific Research Grant) (PNU-2008–101-20080608000).
Disclosure Statement
No competing financial interests exist.