
Abstract
Articular cartilage injuries and degeneration affect a large proportion of the population in developed countries world wide. Stem cells can be differentiated into chondrocytes by adding transforming growth factor-β1 and dexamethasone to a pellet culture, which are unfeasible for tissue engineering purposes. We attempted to achieve stable chondrogenesis without any requirement for exogenous growth factors. Human mesenchymal stem cells were transduced with an adenoviral vector containing the SRY-related HMG-box gene 9 (SOX9), and were cultured in a three-dimensional (3D) hydrogel scaffold composite. As an additional treatment, mechanical stimulation was applied in a custom-made bioreactor. SOX9 increased the expression level of its known target genes, as well as its cofactors: the long form of SOX5 and SOX6. However, it was unable to increase the synthesis of sulfated glycosaminoglycans (GAGs). Mechanical stimulation slightly enhanced collagen type X and increased lubricin expression. The combination of SOX9 and mechanical load boosted GAG synthesis as shown by 35S incorporation. GAG production rate corresponded well with the amount of (endogenous) transforming growth factor-β1. Finally, cartilage oligomeric matrix protein expression was increased by both treatments. These findings provide insight into the mechanotransduction of mesenchymal stem cells and demonstrate the potential of a transcription factor in stem cell therapy.
Introduction
Many surgical therapies have been developed to restore the integrity of articular cartilage. They use either pieces of cartilage tissue (mosaicplasty) or autologous chondrocytes harvested from a non-load-bearing area.2,3 Their main drawback is that they introduce new damage sites into the joint. Surgical methods, such as spongialization and microfracturing, facilitate the migration of cells with regenerative capacity from the marrow space.4,5 These produce fibrocartilage, the presence of which temporarily alleviates pain but is functionally inadequate in the long term.
Clearly, it is necessary to develop a repair strategy using autologous, multipotent cells, and introduce them in a way that they are able to integrate well into the existing cartilage structure. Bone-marrow-derived multipotent cells are able to differentiate into chondrocytes and are potentially ideal candidates to replace autologous chondrocytes.6,7
A possible solution is to differentiate these cells after isolation by overexpressing the chondrogenic master-gene SRY-related HMG-box gene 9 (SOX9) and use these cells to fabricate a cartilage-like implant by seeding them on a scaffold.
SOX9 binds to enhancer sequences in the promoter region of several cartilage genes, thereby enhancing their expression in chondrocytes. In collagen type II α1 chain (COL2A1) it binds within a 48 bp enhancer region located in the first intron. 8 It acts in concert with two cofactors: long form of SRY-related HMG-box gene 5 (L-SOX5) and SRY-related HMG-box gene 6 (SOX6), and these three factors regulate chondrogenic genes such as aggrecan. 9 The two cofactors are normally expressed in mesenchymal cells; therefore, a virus containing SOX9 is sufficient to enhance the level of chondrocytic genes. 10
Mechanical stimulation is a natural constituent of the chondrocyte environment and is known to affect gene expression and re-differentiation of chondrocytes. 11 We have shown in our custom-made bioreactor that chondrocytes respond to dynamic compression by changing their gene expression profile. 12 Other cell types are also affected by mechanical stimulation. The chondrogenic differentiation of chick limb bud stem cells was enhanced by cyclic compression. 13 The differentiation of precursor cell types, such as mesenchymal stem cells (MSCs), is also affected by load.14–17 We have previously demonstrated that the chondrogenic differentiation induced by load involves transforming growth factor-β1 (TGF-β1) signaling. 16
Further, in a possible tissue engineering scenario, SOX9-transduced MSCs could be delivered into the cartilage defect, and the constructs mechanically loaded in vivo. We used a previously established fibrin–polyurethane (PU) scaffold, which allows for chondrogenesis while providing a construct that can be subjected to load.16,18 Our bioreactor system was designed to mimic the mechanical environment of human joints to model such a situation as closely as possible in vitro. 19
We aimed to investigate the interplay between SOX9 overexpression and mechanical stimulation within the context of a tissue engineering approach. We hypothesized that both treatments would enhance the chondrocytic phenotype of human MSCs and that a combination of the two would result in a superior matrix quality. The experiments were carried out in the absence of exogenous TGF-β and dexamethasone. This was to more accurately mimic the in vivo situation to determine what would occur if the transduced cells were directly implanted without receiving a chondrogenic signal in vitro.
Materials and Methods
MSC isolation
Human bone marrow was obtained with ethics approval and the written consent of the patients. The bone marrow was diluted 1:5 with Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Paisley, United Kingdom) containing 5% fetal calf serum (FCS). The mixture was layered on a Ficoll (Sigma-Aldrich Chemie, Steinheim, Germany) cushion and centrifuged at 800 g for 20 min. Detailed isolation procedure was described previously. 20
MSC expansion and freezing
Isolated cells were cultured at 37°C, 5% CO2, and 95% humidity in α-modified essential medium (Invitrogen), 10% FCS, and 5 ng/mL recombinant human basic fibroblast growth factor (Fitzgerald Industries International, Concord, MA). The medium was changed after 2–3 days and then twice a week. Cells were passaged once, grown to confluence, and then stored at −80°C in fetal bovine serum (Invitrogen), containing 10% dimethyl sulfoxide at 2 million cells per mL. Cells were thawed and expanded to the desired number (one to two passages) in α-modified essential medium 10% FCS and 5 ng/mL basic fibroblast growth factor.
MSC transduction
Third-passage cells were transduced with adenovirus containing either green fluorescent protein (GFP) (AdGFP) or both GFP and SOX9 (AdSOX9).21,22 The lanthofection procedure was described previously by Palmer et al. 23
Briefly, virus was diluted in 8 mL DMEM per 300 cm2 flask to about 10,000 viral particles/cell (which is approximately 100 MOI), and LaCl3 (Sigma-Aldrich GmbH, Buchs, Switzerland) was added dropwise to a final concentration of 100 μM. After 30 min of incubation the solution was added to the flasks containing 20 mL DMEM with 10% fetal bovine serum and incubated overnight. On the average 95% transduction efficiency was reached with this method.
MSC seeding
The cells were pretreated with 70 U/mL collagenase (Worthington, Lakewood, NJ), and detached with 0.5 g/L trypsin + 0.2 g/L Na4EDTA (Invitrogen). The collected cells were seeded on cylindrical fibrin-PU scaffolds (d = 8 mm, h = 4 mm, 2 × 106 cells per scaffold) as previously described.24,25
Culture conditions
The cells were cultured at 37°C, 5% CO2, and 95% humidity in DMEM with 4.5 g/L glucose and 2.2 g/L NaHCO3, nonessential amino acids, containing 11.5 mg/L L-proline (Invitrogen), 50 μg/mL ascorbic acid 2-phosphate sesquimagnesium salt hydrate (Sigma), ITS + 1 (10 μg/mL insulin from bovine pancreas, 5.5 μg/mL human transferrin [substantially iron free], 5 ng/mL sodium selenite, 0.5 mg/mL bovine serum albumin, and 4.7 μg/mL linoleic acid; Sigma), and 100 U/mL penicillin + 100 μg/mL streptomicin (Invitrogen). Also, 5 μM ɛ-aminocaproic acid (Sigma) was added to inhibit fibrinolysis by plasmin. 26 The medium was changed three times a week.
Bioreactor and loading
Mechanical conditioning of cell–scaffold constructs was performed using an incubator-installed bioreactor at 37°C, 5% CO2, and 85% humidity. 19 Briefly, a commercially available ceramic hip ball (32 mm in diameter) was pressed onto the cell-seeded scaffold. Interface motion was generated by oscillation of the ball about an axis perpendicular to the scaffold axis. Superimposed compressive strain was applied along the cylindrical axis of the scaffold. After a 6-day preculture the samples were exposed to ball oscillation of ± 25° at 1 Hz with 10% static offset strain for 1 h per day for 8 days. In the following 6 days, additional dynamic compression was applied at 1 Hz with 10% sinusoidal strain, superimposed on the 10% static offset strain, resulting in strain offsets ranging between 10% and 20%. Between loading cycles, constructs were kept free swelling, without surface contact with the ball. Groups of nonloaded constructs served as controls. After 6 days in preculture and 14 days of loading in total (week 3), constructs were vertically cut into two halves, one half for biochemical analysis and the other half for gene expression analysis.
Groups
The study contained four treatment groups: AdGFP-transduced, nonloaded (GFP–control); AdGFP-transduced, loaded (GFP–load); AdSOX9-transduced, nonloaded (SOX9–control); AdSOX9-transduced, loaded (SOX9–load).
In the first two experiments there were also nontransduced controls that showed no significant difference compared to their AdGFP counterparts and were omitted in later experiments.
DNA content
Scaffolds were digested with 0.5 mg/mL recombinant Proteinase-K (Roche Diagnostics, Mannheim, Germany) at 56°C overnight. DNA concentrations were determined with the Hoechst method. 27 Fluorescence intensity was measured with an HTS 7000 Perkin Elmer Bio Assay Reader (Norwalk, CT).
Glycosaminoglycan content
Glycosaminoglycan content (GAG) was determined with dimethylmethylene blue using bovine chondroitin sulfate as the standard. 28 The total GAG content of the culture medium, collected at each medium change, was also measured.
Real-time polymerase chain reaction
Samples were lysed with TRI-reagent (Molecular Research Center, Cincinnati, OH), and RNA was isolated according to the manufacturer's instructions. RNA was reverse transcribed with TaqMan reverse transcription kit (Applied Biosystems, Foster City, CA) using random hexamers. For real-time polymerase chain reaction (PCR) we used TaqMan PCR, on a GeneAmp 7500 Real-Time PCR System (Applied Biosystems) with 18S as the control. 29 The primers and probes are listed in Table 1.
B3GAT3, β-1,3-glucuronyltransferase 3; CCL3, chemokine (C-C motif) ligand 3; COL2A1, collagen type II α1 chain; COL10A1, type X collagen α1; COMP, cartilage oligomeric matrix protein; IL1B, interleukin-1β; PRG4, proteoglycan 4; SOX9, SRY-related HMG-box gene 9; TGFB1, transforming growth factor-β1.
TGF-β1 enzyme-linked immunosorbent assay
Medium samples taken at day 20 (conditioned for 2 days) were analyzed for TGF-β1 concentrations by enzyme-linked immunosorbent assay (ELISA) (DY240; R&D Systems, Minneapolis, MN) according to the manufacturer's instruction.
35 S incorporation
On day 20 the medium was changed and supplemented with 2.5 μCi/mL 35 S (Na2SO4). The samples were incubated for 24 h, after which the conditioned medium was collected and the scaffolds were digested with proteinase-K. Samples were eluted on PD-10 desalting columns (GE Healthcare, Freiburg, Germany), and each 0.5 mL fraction was measured separately in a scintillation counter. The first four fractions (high-molecular-weight compounds) were considered as incorporated 35 S.
Aggrecan immunohistochemistry
Scaffold samples were fixed in 100% methanol before cryosectioning at 20 μm. The sections were reduced with 10 mM dithiothreitol, and alkylated with 40 mM iodoacetamide. Then, the sections were pretreated with Chondroitinase ACII at 0.025 U/mL and probed with anti-aggrecan antibodies (1-C-6; Developmental Studies Hybridoma Bank, University of Iowa, Iowa City, IA) at 4°C overnight. The primary antibody was detected using horse anti-mouse followed by incubation with avidin–biotin–peroxidase complex (Vectastain ABC Kit; Vector Laboratories, Burlingame, CA). Peroxidase activity was observed using 3-30-diaminobenzidine as the substrate, and sections were counterstained with Meyer's hematoxylin.
Promoter analysis
We used the Database of Orthologous Promoters to screen the binding motifs conserved across taxonomic groups. 30 About 3000 bp upstream of the transcription initiation sites of SOX5 and SOX6 were analyzed.
Statistics
Statistical analysis was performed with the R package for statistical computing. 31 None of the data sets were normally distributed; therefore, the Kruskal–Wallis rank sum test was used to determine between-group differences. If the difference was significant at a p ≤ 0.05 level, pair-wise Wilcoxon rank sum test comparisons were calculated, and the p-values were adjusted by Bonferroni's method.
The linear regression lines were calculated with the least square method. For correlation tests, Pearson's product–moment correlation coefficients were calculated and are shown on the graphs (r).
Results
Gene expression
Mechanical loading resulted in a trend toward increased SOX9 expression, which was not significant (data not shown). However, load consistently induced expression of lubricating protein proteoglycan 4 (PRG4) compared to nonloaded controls by week 3 (p ≤ 0.01). Overexpression of SOX9 alone had no effect on PRG4 expression (Fig. 1A). Type X collagen α1 (COL10A1) was also upregulated by mechanical load about fourfold with no effect induced by SOX9 (p ≤ 0.01 GFP–load and SOX9–load compared to GFP–control) (Fig. 1B).

Expression of cartilage-specific genes. After 1 week of preculture and 2 weeks of mechanical load, mRNA expression was calculated using the ΔΔCt method using 18S as the housekeeping gene and untransduced unloaded control as the calibrator. As can be seen, PRG4 (
Neither COL2A1 nor ACAN were upregulated by mechanical stimulation alone. COL2A1 was upregulated 100–1000 times by SOX9 transduction after 3 weeks (p ≤ 0.05) (Fig. 1C). Aggrecan expression was also increased about 100 times by AdSOX9 treatment compared to control (p ≤ 0.05) and load had no effect on this response (Fig. 1D).
Mechanical load alone upregulated cartilage oligomeric matrix protein (COMP) levels 10-fold (p ≤ 0.05) and SOX9 overexpression induced 4-fold higher expression than control (p ≤ 0.05) (Fig. 1E). The combination of the two treatments (SOX9–load) resulted in a 30-fold increase compared to control (p ≤ 0.05).
Gene expression of SOX9's two cofactors, L-SOX5 and SOX6, was strongly associated with SOX9 expression (Fig. 2). High SOX9 expression values were observed in the AdSOX9-treated groups (SOX9–control and SOX9–load). These groups also exerted higher expression of L-SOX5 and SOX6 (ΔCt normalized to 18S). SOX9 expression in the transduced cultures decreased with time to around 0.1% that seen on D0, and so did the message of the other two SOX proteins, although to a lesser extent. Nevertheless, they were still significantly higher than the controls at the last time-point (L-SOX5, threefold; SOX6, fourfold; SOX9, sevenfold; 3 weeks, data not shown).
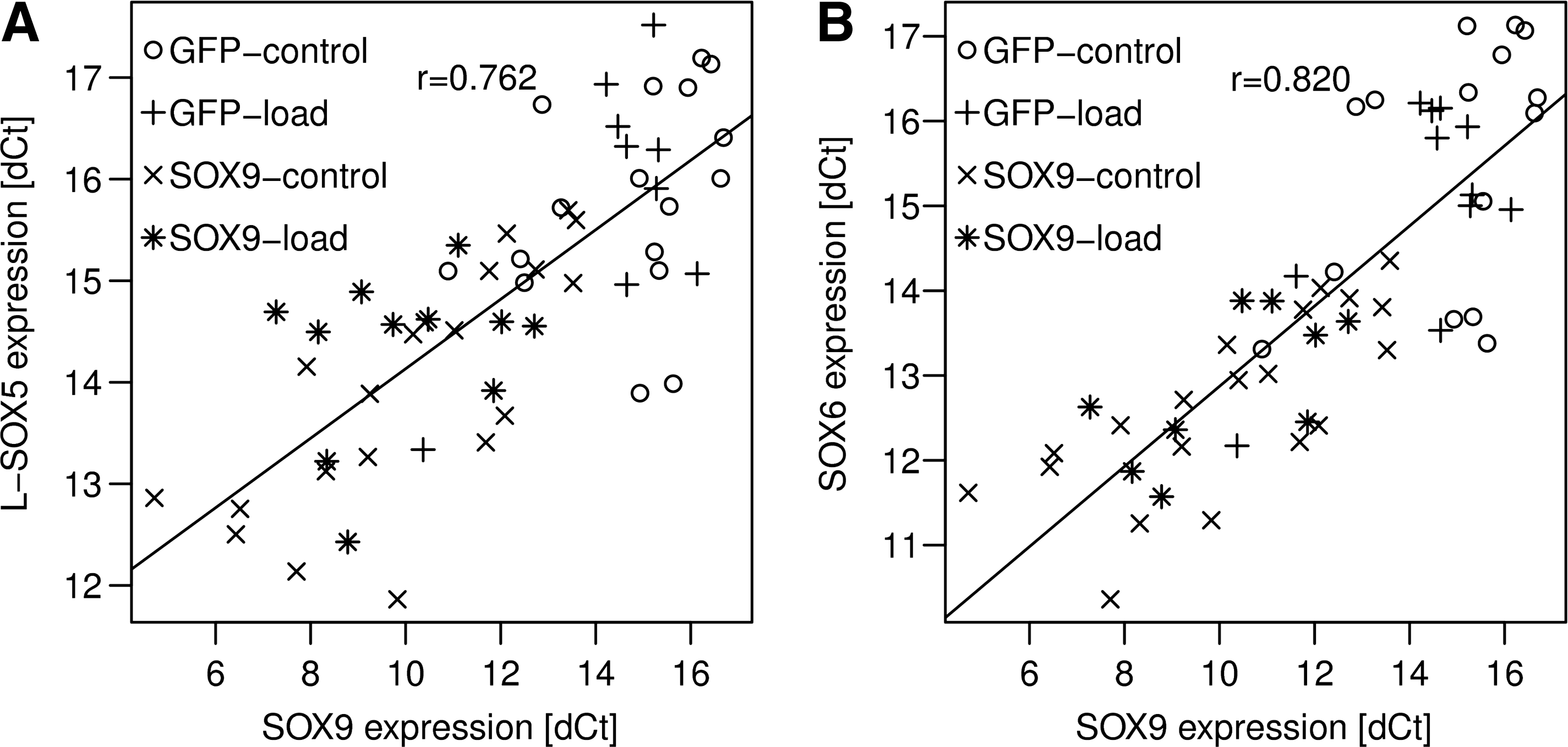
Correlation of SOX9 expression with L-SOX5 and SOX6 expression. Expression of L-SOX5, SOX6, and SOX9 was investigated in each of the four treatment groups. Expression of the L-SOX5 and SOX6 cofactors correlated highly with expression of SOX9. This correlation was independent of time, mechanical load, or SOX9 overexpression. Under all conditions, an increases in SOX9 lead to an increase in both L-SOX5 and SOX6 (p < 0.001). ΔCt values of SOX9 and L-SOX5 (
GAG production
No differences in DNA content were seen between any of the groups (data not shown). About 20%–30% of the GAGs produced during the 3 weeks were retained within the sponges, and the rest was released into the medium. This resulted in no significant differences between any of the treatments regarding the GAG content of the scaffolds (data not shown).
The released amount, however, showed consistent and significant differences (Fig. 3A). After 1 week of culture, the total GAG released from AdSOX9-treated scaffolds was only 75% of control. By the second and third week time-points the loaded scaffolds released 1.4-fold and 1.5-fold more total GAG than controls, respectively. The largest increase was seen in the SOX9–load group between weeks 2 and 3.

Glycosaminoglycan (GAG) production. (
In the final week, the SOX9–load group produced significantly more GAG (Fig. 3A, p < 0.05). The amount released per day was found to be on average 1.6 × higher in SOX9–load compared to GFP–load, and 2.5 × higher than nonloaded controls at the 3 week time-point (Fig. 3B).
35 S incorporation
To see whether this was a result of higher GAG synthesis, sulfate incorporation was measured with a radioactive assay after 3 weeks (Fig. 3C). It was found that SOX9–load samples incorporated at least twice as much 35 S isotope compared with other treatment groups (p ≤ 0.05). This correlates well with the GAG released into the medium on day 20 (Fig. 3B).
Immunohistochemistry
Increased GAG production resulted in an apparently stronger ACAN staining in the SOX9–load samples (Fig. 4). Interestingly, only a weak staining can be observed in the SOX9–control samples, despite high ACAN mRNA expression. AdGFP samples also produced some ACAN, which is localized along the edges of the sponges, whereas SOX9–load exerts strong pericellular staining in cells inside the PU sponge (Fig. 4, see arrowheads), as well as along the edges.

Localization of aggrecan. Representative images stained using aggrecan-specific antibodies. The left column shows an overview image. The negative controls from neighboring sections of the same specimen can be seen in the nested box (scale bars: 500 μm). The right column shows higher magnification images (scale bars: 100 μm). The edges stain more intensely due to the higher cell content. In the GFP–load and SOX9–load groups individual cells that have a positive staining for their extracellular matrix can be seen (arrowheads). All sections were counterstained with Meyer's hematoxylin.
TGFB1 gene expression and ELISA
Gene expression of TGFB1 was threefold higher in SOX9–load samples and twofold higher in GFP–load samples compared to GFP–control after 3 weeks (p ≤ 0.05) (Fig. 5A). ELISA of the cell culture supernatants at the same time-point showed that this translated into increased protein concentration only in the SOX9–load group (Fig. 5B). The TGF-β1 levels of SOX9–load samples were 2.3- to 3.5-fold higher than any other group reaching concentrations of 465 ± 277 pg/mL (p ≤ 0.05). There is strong positive correlation between the concentration of TGF-β1 and the GAG production rate of the respective sample (Fig. 5C).
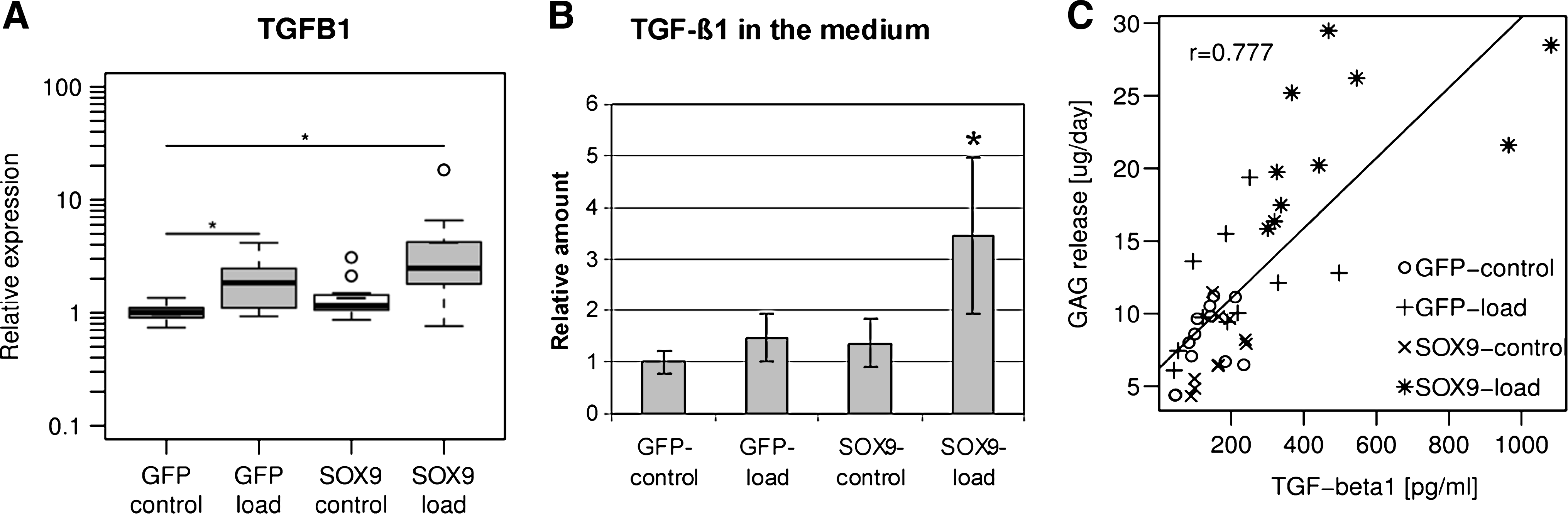
TGF-β1. (
Discussion
We investigated a cartilage tissue engineering strategy where the SOX9 transcription factor is overexpressed to induce the differentiation of MSCs into chondrocytes. The constructs were mechanically stimulated in a custom-made bioreactor, but no further chondrogenic stimulus was provided.
The observed genetic and biochemical markers can be divided into three groups. Some of them were influenced only by mechanical stimulation, some only by SOX9 overexpression, whereas the third group shows an additive or synergistic effect of the two treatments.
Mechanical load in the presence of further chondrogenic signals, such as dexamethasone with or without TGF-β1, has been shown to enhance chondrogenesis in a dose-dependant manner. GAG synthesis and gene expression levels are positively related to the frequency and amplitude of mechanical load. This has been seen in human MSC pellets and hydrostatic pressure, 32 static loaded mouse embryonic limb bud mesenchymal cells, 11 cyclic loaded chick limb bud mesenchymal cells, 13 and human stem cells seeded with the PU scaffold used in this study.16,33 Mukherjee et al. observed contrary results in dynamic fluid pressure-regulated rabbit periosteal explants, low levels of dynamic fluid pressure (13 kPa at 0.3 Hz) significantly enhanced chondrogenesis, whereas higher pressures (103 kPa at 0.3 Hz) completely inhibited chondrogenesis. 34 The reason for the difference is currently unclear.
In this study, instead of using load in combination with soluble chondrogenic stimuli, we investigated whether load could be used in combination with overexpression of SOX9 in a strategy that could theoretically be used within the clinic. As we have used an adenoviral vector, gene expression of SOX9 was transient. It peaked before day 6 and showed a decreasing trend through the rest of the culture period (data not shown). In this culture system, SOX9 overexpression led to the upregulation of its known cartilage-specific target genes (COL2A1 and ACAN) in the absence of exogenous TGF-β or dexamethasone. Their expression was stable throughout the 3-week culture period, which suggests that the amount of active SOX9 protein was adequate to enhance expression of these genes. Interestingly, COL2A1 and ACAN were not upregulated by load under these conditions. We have previously shown that mechanical stimulation in the presence of dexamethasone alone is sufficient for chondrogenesis, 16 suggesting that under in vitro conditions dexamethasone may be required. Using the classical chondrogenic culture medium (10 ng/mL TGF-β1 and 10−7 dexamethasone), the same study demonstrated an approximately 100-fold greater increase in Col II and ACAN expression under load when compared to that seen in this study in the SOX9-loaded group. This again indicates that under in vitro conditions further factors are required.
COMP is expressed in tissues subjected to mechanical strain, such as cartilage and tendon. 35 In cartilage expression of other, cartilage-specific genes (ACAN and COL2A1) are also seen, which are not seen in other tissues. COMP has been shown to be highly upregulated in response to load in chondrocytes. 36 It has also been shown to promote chondrogenesis, although in a cell line. 37 Taken together, these data would suggest an important role of COMP for cartilage development and its increase is a further indication of chondrogenesis. The two treatments had an additive effect on expression of COMP. The 30-fold upregulation by the combined treatment is approximately the product of 4-fold increase by SOX9 and 10-fold increase by load.
COL10A1 expression was found to be slightly induced by mechanical load. This protein is known as a marker of chondrocyte hypertrophy, but its applicability for in vitro studies has been questioned. 38 Also, the small change observed here (3–4 ×) may not be biologically significant.
We also showed that PRG4 expression was induced by mechanical load in human MSCs. Previously, its expression has been reported to be increased by mechanical load in chondrocytes. 12 It was postulated by Neu et al. that this happens through TGF-β1 signaling pathway. 39 This is in agreement with our findings in the SOX9–load treatment group, where both TGF-β1 protein and PRG4 mRNA levels were elevated. However, the GFP–load group showed a comparable increase in PRG4 expression despite the more subtle increase in TGF-β1 concentration (3.5 × vs. 1.5 × ). This—statistically insignificant—increase in TGF-β was similar to that of the SOX9–control group, where no PRG4 upregulation was observed (1.5 × vs. 1.3 × ). This suggests to us that there are other mechanisms involved in the mechanoregulation of PRG4 besides the TGF-β1 pathway.
Although ACAN mRNA expression was increased in all SOX9-treated samples, an increase in aggrecan synthesis was only observed in the group where SOX9 overexpression was combined with mechanical loading. The discord between ACAN increase and actual GAG synthesis has been reported in other studies and the reasons are as yet unknown.18,40,41 To try to explain this phenomenon, several aspects of the cell culture system were investigated. High expression of inflammatory cytokines can be responsible for lower expression and lower activity of β-1,3-glucuronyltransferase 3 (B3GAT3), which in turn causes deficient glycosilation. 42 No significant difference was found in expression of interleukin-1β (IL1B) or chemokine (C-C motif) ligand 3 (CCL3) (data not shown), both of which were implicated in inflammatory response in arthritis. 43 B3GAT3 expression of SOX9–control was 75% of that of GFP–control (data not shown). This difference may not be significant biologically and does not explain the higher GAG production rate in SOX9–load samples. SOX9 overexpression alone has been shown to increase GAG production in both chondrocytes and cartilage explants.44,45 This would suggest that some required cofactor, present in mature chondrocytes, is lacking in the chondrogenic environment the MSCs are exposed to within this study.
Under the culture conditions used in this study SOX9 alone did not stimulate GAG production. Mechanical load stimulated GAG production, but the combination of the two led to a more sustained increase, which was more noticeable in the third week of culture. About 20%–30% of the GAG produced during the 3 weeks was retained within the sponges and the rest was released into the medium. This release is due to a poorly structured pericellular matrix, which is still under development and therefore is inefficient at retaining the newly synthesized GAGs. Combined with the extremely porous nature of the scaffold–fibrin composite, this leads to a large proportion of the newly synthesized matrix being released. During the first 2 weeks the GFP–load and SOX9–load groups behaved comparably. However, in the final week of culture there was a significant increase in GAG synthesis in the SOX9–load group compared to the other treatments. We noted that the concentration of TGF-β1 in the medium was closely related to the GAG synthesis in all treatment groups. TGF-β1 has long been known to promote chondrogenesis of MSCs.6,7 According to the correlation coefficient calculated in our study, variation in TGF-β1 concentration is responsible for 60% of the variation in GAG production (r 2 = 0.604). Potentially, GAG production can be enhanced by increasing the TGF-β1 concentration or by the addition of dexamethasone. In an in vivo situation this would be possible by utilizing a drug delivery system. Although the immunohistochemistry demonstrated the presence of aggrecan, no staining for collagen II was detected. We believe the reason for this to be threefold. First, as the two additional components of chondrogenic induction medium were missing (TGF-β and dexamethasone) the chondrogenic response will be reduced. As previously mentioned we have carried out other studies where medium containing dexa but lacking in TGF-β was used and a more robust response was seen. 16 Second, the cell density is very low, so it is likely that under the short term of the experiment it would be difficult to produce (and retain within the scaffold) enough collagen to be detectable. Most of the early matrix synthesized will be released into the medium due to the porous nature of the scaffold. Third, collagen II synthesis is often poor even in optimal chondrogenic medium.
We have shown that SOX9 was able to induce expression of cartilage-specific genes, including its cofactors L-SOX5 and SOX6. It has been previously demonstrated that SOX9 leads to the upregulation of L-SOX5 and SOX6, 10 and we have demonstrated that there is an exponential correlation between expression of SOX9 and its two cofactors. Expression of this so-called SOX trio was restricted to cartilaginous tissues in a mouse embryo. 46 The binding of SOX9 and at least one of SOX6/L-SOX5 is necessary for expression of COL2A1, collagen type XI α1, and ACAN.9,46–49 We identified several instances of the 7 bp SOX-binding consensus sequence (CT/ATTGT/AT/A) in the 5′ upstream region of L-SOX5/SOX6. These putative binding sites are located at the following positions in human: 2124 and 764 bp before transcription start site of SOX6; and 2149, 1736, and 224 bp before the transcription start site of SOX5. Further promoter analysis is necessary to prove that SOX9 affects its cofactors directly through these binding sites, which is out of the scope of this report.
Growth factors such as TGF-β1 have diverse effects through several pathways on multiple transcription factors, and in the case of TGF-β1 this results in a more complete chondrogenesis. Our results show that SOX9 is able to initialize chondrogenic differentiation of MSCs in the absence of exogenous chondrogenic factors and dexamethasone in vitro but only in combination with mechanical stimulation. This suggests that this strategy would function in vivo where load would be applied during rehabilitation. It also indicates that expression of SOX9 alone only affects a subset of the genes involved in chondrogenesis and therefore other factors, such as load, are required. These findings confirm the feasibility of a cartilage repair strategy, where stem cells overexpressing SOX9 could be delivered into a cartilage defect without prior cultivation in the chondrogenic medium. During the rehabilitation procedure an appropriate mechanical loading regime would help their chondrogenic differentiation and hopefully the production of integrating stable articular cartilage tissue.
Footnotes
Acknowledgments
We wish to thank Prof. Chris Evans (Harvard Medical School, Boston, MA) for kindly providing the SOX9-containing adenoviral vector. The fibrin used in the experiments was generously supplied by Baxter Biosurgery (Vienna, Austria). This work was supported by the Swiss National Fund (SNF 320000-116846/1).
Disclosure Statement
No competing financial interests exist.