
Abstract
Mechanical properties of engineered tissues should ideally match those of the tissues that are replaced. Engineered skin (ES) is often orders of magnitude weaker than normal skin, which can lead to damage during application and improper function after engraftment. Hypothetically, application of strain during culture of ES may lead to improved mechanical properties. ES comprised of electrospun collagen scaffolds, human dermal fibroblasts, and epidermal keratinocytes were fabricated and cultured at the air–liquid interface. ES was loaded in vitro into a strain apparatus, strained to 0% (restrained), 5%, 10%, 20%, or 40%, with unstrained ES as a control, and cultured for 10 days. ES cultured under 10% and 20% strain were significantly stronger than unstrained controls. ES cultured under 20% strain showed upregulation of many genes encoding structural extracellular matrix proteins, including collagen type I alpha 1 and fibronectin 1. Mechanical stimulation significantly increased epidermal cell proliferation and enhanced epidermal differentiation with 5%, 10%, and 20% strain. Improved strength in the 10% and 20% strain groups is likely a result of increased extracellular matrix production coupled with enhanced epidermal differentiation. These improvements to ES may facilitate surgical application, prevent damage during transplantation, and may result in improved functional outcomes after engraftment.
Introduction
A majority of studies have focused on altering the mechanical properties of the scaffold to improve the strength of ES. Chemical crosslinking methods have been utilized to slow degradation rates and improve mechanical properties of collagen scaffolding used for ES.13–16 Although chemical crosslinking has been shown to improve biochemical stability and mechanical strength compared with noncrosslinked collagen,12,17,18 this increase in scaffold strength does not always correlate with significant increases in tissue strength. 12 For example, the tensile strength of ES generated from 1-ethyl-3-3-dimethylaminopropylcarbodiimide hydrochloride (EDC)-crosslinked collagen sponges was more than an order of magnitude weaker than published values for native human skin strength.9,10,12 Thus, ES strength is not directly proportional to the strength of the polymer scaffolds used.12,19 A combination of superior biological properties, including cell viability and tissue morphogenesis, and scaffold properties are necessary to generate a high-strength engineered tissue.
Mechanical stimulation has been widely used to promote the in vitro formation of engineered tissues such as blood vessels, heart patches, bone, and cartilage.20–24 For instance, mechanically stimulated mesenchymal stem cell-seeded collagen constructs were significantly stiffer than their nonstimulated counterparts. 25 Intermittent hydrostatic pressure increased cellular proliferation and glycosaminoglycan content in engineered articular cartilage. 26 In dermal substitutes comprised of fibroblasts on collagen scaffolds, cyclic strain resulted in increased proliferation and thicker tissue constructs. 27 Straining monolayer cultures of keratinocytes to 20% elongation increased proliferation, increased keratin 6 production, and decreased keratin 10 production. 28 Although mechanical stimulation of the proper magnitude and duration has been shown to have positive effects on many tissues, little is known regarding the effects of mechanical stimulation on three-dimensional ES containing both fibroblasts and keratinocytes.
In this study, the effects of mechanical strain on ES development, mechanical properties, and gene expression were evaluated. Multicellular, three-dimensional ES was fabricated and strained during culture (Fig. 1). Tissue organization, morphogenesis, cellular proliferation, strength, and gene expression in the resultant ES were assessed.
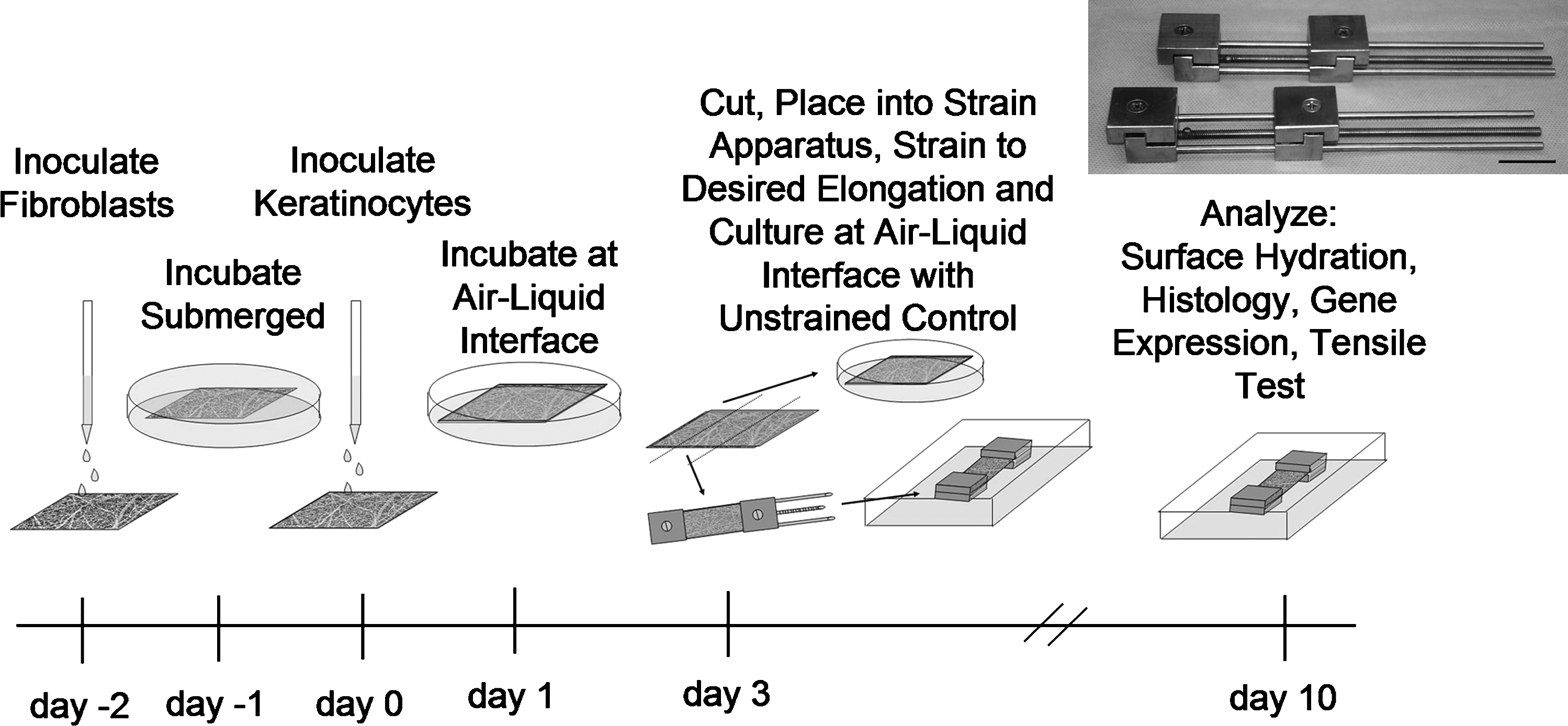
Experimental timeline for the inoculation, application of uniaxial strain, and subsequent culture of ES. Inset: Photograph of custom-designed static strain devices. Scale bar = 2 cm. ES, engineered skin.
Materials and Methods
Collagen scaffolds
Collagen scaffolds were prepared from acid-soluble collagen from bovine hide (Kensey Nash, Exton, PA). Collagen, solubilized in hexafluoropropanol (Sigma-Aldrich, St. Louis, MO) at a concentration of 10% (wt/vol), was electrospun at a potential of 30 kV onto a grounding plate that was positioned 20 cm from the tip of the needle. Electrospun scaffolds were physically crosslinked by vacuum dehydration at 140°C for 24 h and then chemically crosslinked in a solution of 5 mM EDC (Sigma-Aldrich) in 100% ethanol for 24 h. The scaffolds were then disinfected in 70% ethanol for 24 h and rinsed thoroughly following a procedure previously described 29 for the preparation for cell inoculation.
Fiber orientation
The morphology of the electrospun collagen scaffold was examined by scanning electron microscopy (FEI Sirion, Hillsboro, OR). The as-spun scaffold was mounted onto aluminum stubs, sputter coated with gold–palladium, and imaged in secondary electron mode with a 5 kV accelerating voltage. To evaluate the changes in scaffold morphology as a result of mechanical strain, scaffolds were left floating in N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid buffered saline (HBS) or loaded into custom-designed strain devices (Fig. 1). Scaffolds were cut into strips (2 cm × 6 cm), loaded into the strain devices with the scaffolds held in place by compression between the grips, and strained to 0%, 5%, 10%, 20%, or 40%. All scaffolds were incubated for 1 week in HBS. The scaffolds were then rinsed for 2 × 5 min with phosphate-buffered saline, dehydrated in a graded alcohol series, and dried in a graded ethanol:hexamethyldisilazane (Ted Pella, Redding, CA) series. 29 The dried samples were mounted and examined via scanning electron microscopy as earlier, with the principal axis of strain oriented vertically within the field of view. Images were collected and fiber orientation with respect to the principal direction of stress was calculated from at least 300 fibers from each group quantitatively via Image J software (http://rsbweb.nih.gov/ij/). Fiber orientation was binned (5°/bin) and plotted as frequency per bin.
Preparation of ES
ES was prepared from adult human keratinocytes (HK) and fibroblasts isolated from skin from elective reduction mammoplasty, which was obtained with approval of the University of Cincinnati Institutional Review Board. The cells were grown in selective growth media 30 and cryopreserved at passage 0 (primary culture) or passage 1, providing a stock of cells for the ES. For inoculation, electrospun collagen scaffolds were measured and placed on top of a polyprolylene mesh (N-terface; Winfield Labs, Richardson, TX) and a polyvinyl alcohol sponge (Merocel; Medtronic, Minneapolis, MN). Human fibroblasts were inoculated onto the electrospun scaffolds at a density of 5.0 × 105 cells/cm2 and incubated at 37°C and 5% CO2 in UCMC 160 medium 30 for 1 day. Scaffolds were again measured and then inoculated with HK at a density of 1.0 × 106 cells/cm2. One day following HK inoculation (ES incubation day 1), the cell–scaffold constructs were placed onto a perforated stainless steel platform covered by a cotton pad to establish an air–liquid interface (Fig. 1).
Mechanical stimulation
ES, at culture day 3, was cut into 2 cm × 6 cm strips and loaded into the strain devices (Fig. 1). ES on N-terface was loaded into each strain device with the epidermal side facing up (i.e., in contact with air) such that the distance between the grips was between 53 and 55 mm (Loriginal) after the ES on N-terface was pulled tight. The N-terface was then cut from the device to ensure that it would not interfere with the application of strain. ES was strained to 0% (restrained with no additional strain), 5%, 10%, 20%, or 40% strain ([Lengthfinal − Lengthoriginal]/Lengthoriginal × 100 = % strain) and placed into separate bioassay dishes (Corning, Corning, NY). Medium was added to the assay dish until it reached the bottom of the ES in order to establish an air–liquid interface. Strips of ES were kept on the lifting platforms as a positive control (Fig. 1). All ES were kept in culture for a total of 10 days with medium exchanged every other day.
Histology
Biopsies for histology from test samples were collected at day 10, embedded in OCT™ compound (Sakura Finetek, Torrance, CA) and cryosectioned to ∼7 μm thickness using standard histological procedures. Sections were stained with hematoxylin and eosin and imaged with light microscopy. Bright-field images were collected with SPOT Advanced Imaging software (Diagnostic Instruments, Sterling Heights, MI), with a total of nine samples per condition.
To observe basement membrane formation, epidermal differentiation markers, integrin expression, and extracellular matrix (ECM) synthesis, biopsies were collected at day 10 for cryosectioning. The cryosections were fixed in methanol for 8 min followed by acetone for 2 min. After fixation, the samples were immunostained for human involucrin (Sigma-Aldrich), human laminin 5 (Biodesign International, Saco, ME), human loricrin (Covance, Emeryville, CA), human CD44 (Santa Cruz Biotechnology, Santa Cruz, CA), or human fibronectin (Chemicon International, Billerica, MA) and were detected with fluorescent secondary antibodies (Invitrogen, Carlsbad, CA). The labeled sections were examined via epifluorescence microscopy (Nikon Microphot FXA, Melville, NY) and images were collected with SPOT Advanced imaging software, with a total of four specimens per group per antibody.
Bromo-deoxyuridine labeling
To determine the proportion of cells in the in vitro skin substitutes that were actively proliferating, ES pieces were exposed to medium containing 65 μg bromo-deoxyuridine (BrdU) for 24 h prior to sample collection. The samples were taken at day 10 in vitro, embedded in OCT compound, and processed for cryosectioning. The slides were then fixed in methanol for 8 min, followed by acetone for 2 min, and rehydrated in phosphate-buffered saline. The sections were colabeled with anti-BrdU-fluorescein isocyanate antibody (BD Biosciences, Franklin Lakes, NJ) and anti-pan-cytokeratin antibody (Invitrogen) by overnight incubation at 4°C. The double-labeled sections were then examined microscopically, with a total of 40 fields of view per group. The number of BrdU-positive cells per microscopic field was recorded and data presented as the number of BrdU-positive cells per field (mean + standard error of the mean [SEM]).
ECM gene expression
After 10 days in culture, biopsies from strained and unstrained control ES were collected and RNA was isolated using RNeasy Mini Kits (Qiagen, Valencia, CA). RNA was converted to biotin-labeled cRNA using the TrueLabeling-AMP 2.0™ Kit (SuperArray Bioscience, Frederick, MD) with biotin-UTP (PerkinElmer, Boston, MA). The labeled cRNA was hybridized following the manufacturer's instructions to the ECM and Adhesion Molecules Oligo Array (SuperArray Bioscience), which can simultaneously analyze the expression of 113 genes. The expression data were analyzed using GEArray Expression Analysis Suite software (SuperArray Bioscience); expression levels were normalized based on signal intensity of control oligonucleotides spotted on the microarray membranes.
Tensile testing
The mechanical properties of mechanically stimulated and control ES were assessed via uniaxial tensile testing to failure (n = 18 for each group). In addition, human skin from elective reduction mammoplasty was obtained with approval of the University of Cincinnati Institutional Review Board to quantify the mechanical properties of NHS (n = 4). ES and NHS were cut into dog-bone–shaped specimens with a gauge length of 20 mm and width of 4 mm. Specimens were mounted into the grips of a material testing system (TestResources) with a 5-N load cell for ES and a 1000-N load cell for NHS and tested to failure at a strain rate of 2 mm/min to avoid specimen dehydration during testing. Samples that did not break within the gauge length were discarded and samples that delaminated during testing were recorded. Percent elongation at failure, ultimate tensile strength (UTS), and stiffness values were measured and reported as mean + SEM.
Statistical analysis
For quantitative assays, repeated measures analysis of variance and Tukey post hoc tests were used to determine statistical significance as a function of mechanical stimulation. The data were presented as mean + SEM, with p < 0.05 considered statistically significant.
Results
Mechanical strain had a profound effect on the orientation of fibers within the acellular scaffold. The control, nonstrained scaffolds had a random fiber orientation with approximately the same number of fibers oriented in all directions (Fig. 2). As the percent strain increased, there was a dramatic increase in fiber alignment in the direction of principal stress. At 40% strain, 87.9% of the fibers were +20° of the principal direction of stress (oriented vertically in the images).

Histograms of collagen fiber alignment as a function of increasing strain in scaffolds incubated in HBS for 10 days: (
Histological evaluation of the ES at culture day 10 revealed that all groups had excellent cellular organization. Fibroblasts densely populated the collagen scaffold of all the ES (Fig. 3). The epidermis was well stratified, with a continuous layer of basal keratinocytes and stratum corneum formation present in all conditions. No gross difference in tissue organization was found between nonstimulated and stimulated skin. However, the epidermis of the static group was observed, in general, to be slightly thinner than that of the mechanically stimulated groups (Fig. 3). Analysis of cell proliferation via BrdU labeling revealed statistically significant increase in cell proliferation in mechanically stimulated groups (p < 0.05; Fig. 4) compared with the control. Mechanically stimulated groups were not statistically different from one another.
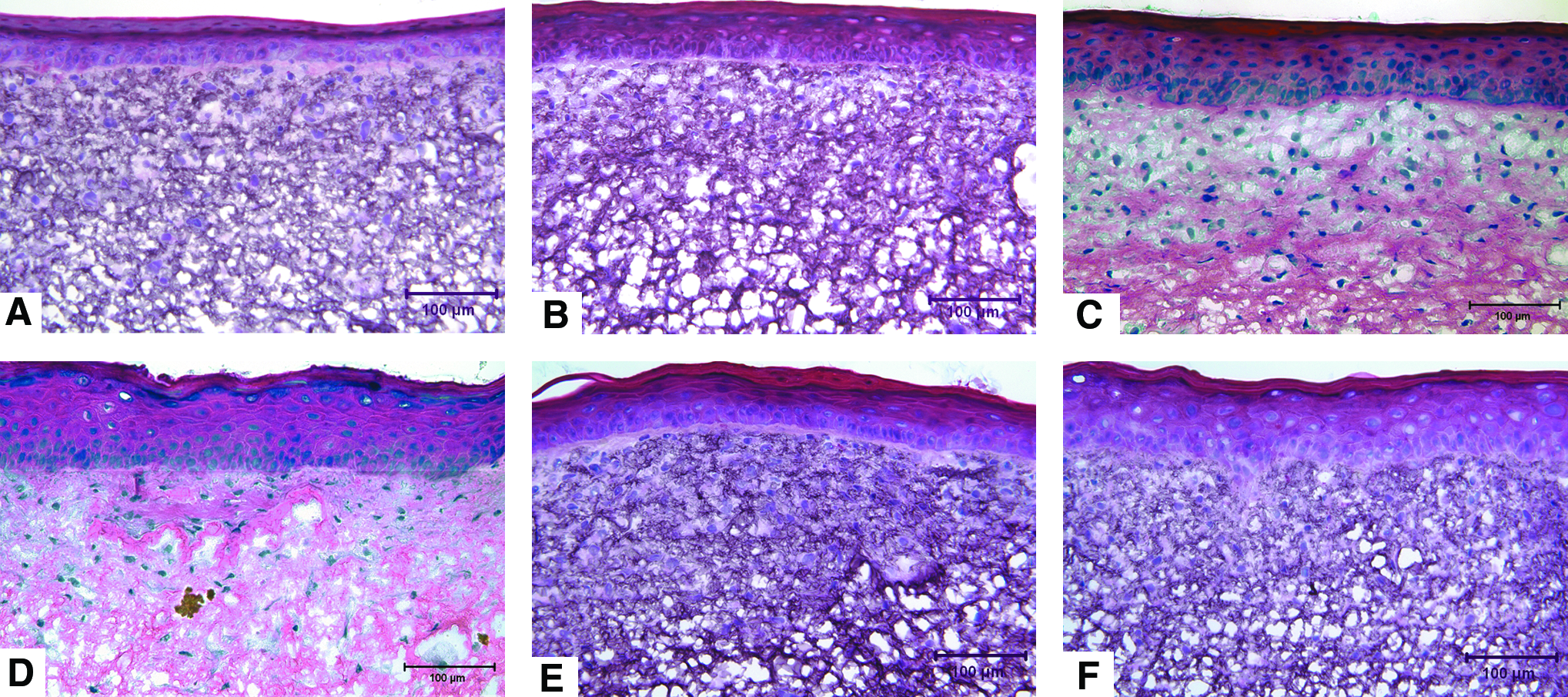
Histological sections of ES at day 10 with (
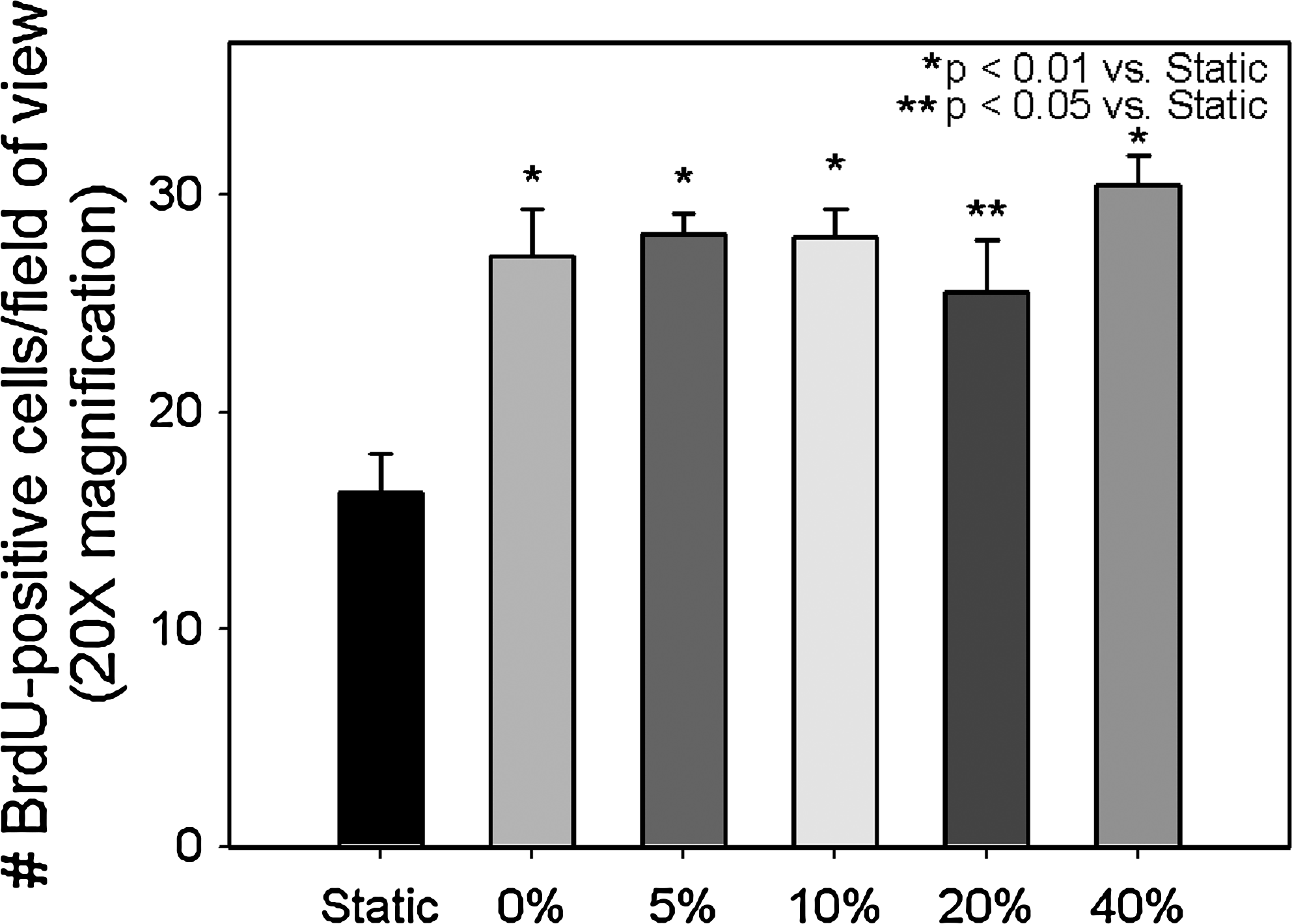
Quantification of actively proliferating cells in ES at day 10. n = 6 per group. BrdU, bromo-deoxyuridine.
Uniaxial tensile testing of the nonstimulated and mechanically stimulated ES after 10 days in culture indicated a significant improvement in strength of ES cultured under 10% and 20% static strain, but all groups were significantly weaker than NHS (Table 1). Linear stiffness scaled positively with mechanical stimulation (Table 1). The 10%, 20%, and 40% were significantly stiffer than the static, 0%, and 5% groups but not different from one another (Table 1). Average percent elongation at failure was lowest in the 40% group and scaled inversely with magnitude of mechanical strain (Table 1); however, there was no statistical difference among the groups.
p > 0.05 versus static.
p < 0.05 versus static and 0%.
p < 0.05 versus static, 0%, and 40%.
p < 0.001 versus all.
p < 0.05 versus static.
Gene expression of ECM proteins and adhesion molecules in ES was assessed at the end of the 10-day culture period (7 days after the mechanical stimulus was applied) in the static, 0%, 20%, and 40% strain groups, as the 5% and 20% groups' mechanical properties were not significantly different than those of the 0% and 10% groups, respectively. The application of 20% strain at day 3 resulted in an upregulation of many genes for ECM and adhesion molecules, including collagen type I alpha 1, laminin alpha 2–5, and integrins alpha 3, 4, and 5, compared with unstrained controls (Fig. 5 and Table 2). However, the relative change in gene expression between the control and stimulated groups was modest and typically ranged between a twofold upregulation and a twofold downregulation. For example, relative gene expression levels for collagen type I alpha 1 and collagen type XII alpha 1 were 2.85-fold and 1.43-fold greater in the 20% group than the control. Gene expression for fibronectin 1 was 1.78-fold greater in the 20% group (Table 2). A 3.34-fold and 2.04-fold upregulation of the gene encoding CD44, the main cell surface receptor for hyaluronate, was found in the 0% and 40% groups, respectively; however, the 20% group had similar expression levels to the unstrained control. Laminin-5 gene expression increased with the application of 0% strain, reached peak expression levels at 20% strain, and fell to control levels with the application of 40% strain (Table 2).

Heat map of gene expression in ES at day 10 cultured with or without mechanical stimulation. Expression levels for many genes were not detectable in the static, 0%, or 40% groups, but were upregulated in the 20% ES group. Color images available online at www.liebertonline.com/ten.
The values shown are normalized expression values for genes that were expressed in at least one sample analyzed, and the ratios of gene expression in 0%, 20%, or 40% strain compared with static culture.
Spots were considered “absent.”
Ratios highlighted in light gray indicate more than twofold increase from static condition. Ratios highlighted in dark gray indicate more than twofold decrease from static condition. Ratios based on one or more “absent” calls should be interpreted with caution.
Immunohistochemistry was used to examine expression differences at the protein level. There were no qualitative differences in CD44 protein expression between stimulated and nonstimulated groups, despite a 3.34-fold upregulation in gene expression in the 0% strain group compared with the control. CD44 was present only within the epidermis in all groups of ES (Fig. 6). A continuous layer of laminin-5 was present in all groups at the dermal–epidermal junction (Fig. 6), with a slightly thicker layer of laminin-5 present in the 5%, 10%, and 20% groups. Immunostaining for fibronectin 1, a high-molecular-weight ECM glycoprotein that binds to integrins, indicated that fibronectin was present within the dermis of all ES (Fig. 6). The static, 0%, and 40% strain groups did not show any positive staining for fibronectin directly below the dermal–epidermal junction, whereas dense staining for fibronectin was seen throughout the dermis of the 10% and 20% strain groups (Fig. 6). Positive staining for involucrin, a cytoplasmic protein precursor of the epidermal cornified envelope, was present in all groups and within all the epidermal layers except for the basal cell layer (Fig. 7). In contrast, loricrin, a major component of the cornified envelope of terminally differentiated keratinocytes, was not present or was localized in small areas in the static, 0%, and 40% strain groups (Fig. 7). Staining for loricrin was positive in a continuous layer of keratinocytes within the upper epidermis of the 5%, 10%, and 20% groups (Fig. 7). Loricrin was more prevalent in the 40% group than the control or 0% groups but was not a continuous layer (Fig. 7).
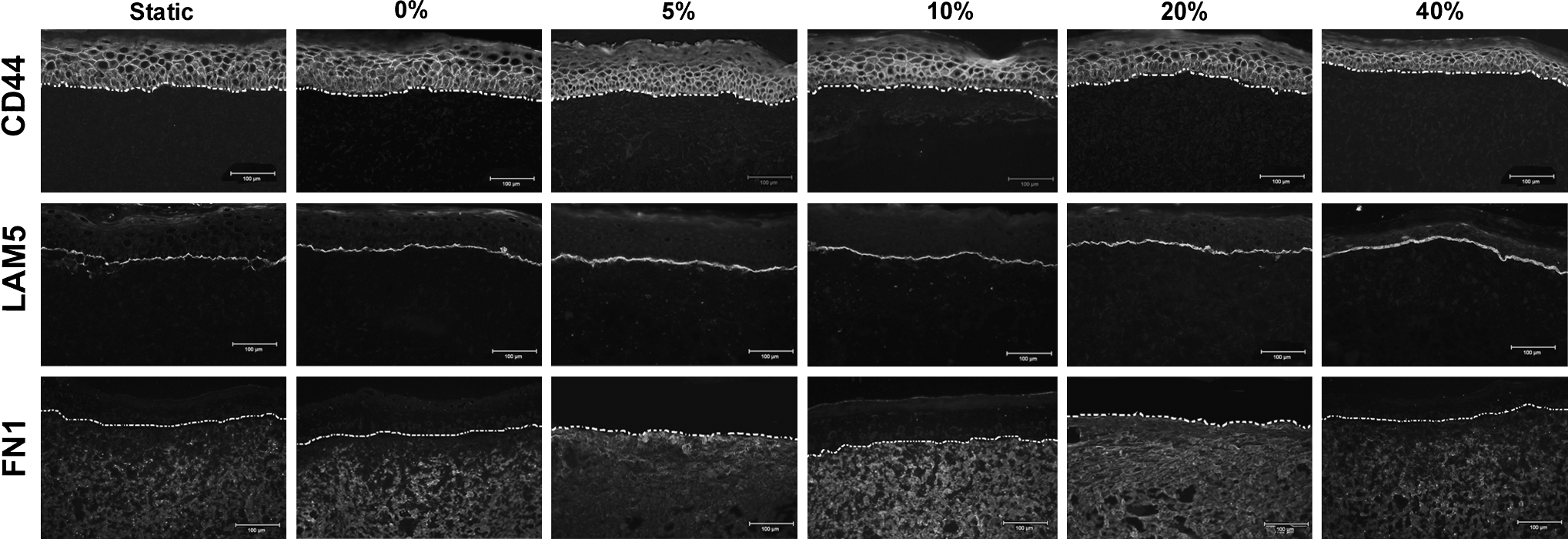
Immunostaining for CD44, laminin-5, and fibronectin 1 in ES at day 10. Dashed white line in each panel indicates the dermal–epidermal junction. Scale bar = 100 μm.

Immunostaining for the nucleus (DAPI) and epidermal markers of differentiation, involucrin and loricrin, in ES at culture day 10 and in normal adult human skin. Dashed white line in each panel indicates the dermal–epidermal junction. Scale bar = 100 μm. DAPI, 4′,6-diamidino-2-phenylindole; NHS, normal human skin. Color images available online at www.liebertonline.com/ten.
Discussion
Mechanical forces are key regulators of tissue homeostasis and are essential for tissue formation, remodeling, and normal function. Mechanical forces also play a central role in the physiology of normal skin and in wound healing. This study demonstrated that mechanical stress impacts the biopolymer scaffold structure, gene expression, and morphogenesis of ES.
Collagen fiber alignment within the acellular scaffold was significantly enhanced in response to mechanical strain. A change in fiber orientation to become more aligned with the principal direction of stress was expected. This response is also seen in collagen fibers within tissues. Uniaxial strains of 5% or greater in the preferred fiber direction in small intestinal submucosa resulted in increased collagen fiber orientation in the direction of stress. 31 Collagen fibers within bovine pericardium altered their peak orientation angle in a strain-dependent manner, with a 30% strain resulting in collagen fiber orientation shifting from 135° to 90° (direction of stress). 32 In this study, a significant change in average fiber orientation was seen at 10% strain and small amounts of alignment were seen in the 0% strain group, which is in contrast to published data that suggest a minimum of 5% strain is required to induce permanent change in fiber orientation. The increase in fiber alignment in the 0% group may be a result of the drying process used to prepare the samples for evaluation on the scanning electron microscope. During the dehydration step, the scaffolds contracted slightly (∼5%) adding to the effective strain the scaffold experienced.
Histological evaluation of the ES at day 10 revealed that there was no significant difference in cellular organization, fibroblast density, or cell penetration among the groups (Fig. 3). However, the epidermis appeared thicker in the mechanically stimulated groups, which correlated with BrdU-cell proliferation data that indicated the keratinocytes within the mechanically stimulated ES were significantly more proliferative (Fig. 4) than the control (static) group. Increased keratinocyte proliferation seen in the ES agrees with previous monolayer culture studies where cyclic strain led to increased cell proliferation compared with no load.28,33–37
Significant improvements to UTS of ES were seen in the 10% and 20% strain groups, with average strength scaling positively with the magnitude of strain applied up to 20% (Table 1). Fiber alignment has previously been shown to have a profound effect on the mechanical properties of acellular scaffolds. When fibers were aligned parallel to the direction of applied force, their UTS showed significantly improved yield stress and modulus in proportion to the alignment. 38 Although ES cultured under 10% and 20% static strain was significantly stronger than nonstimulated controls, it is unlikely that simple scaffold fiber reorientation was solely responsible for the observed increase in mechanical strength, as the strength of the ES is, on average, an order of magnitude stronger than the acellular scaffolds. In addition, the scaffold with the highest amount of fiber alignment, 40% strain, did not result in improved strength compared with the control (Table 1); only linear stiffness was improved which is highly dependant on fiber orientation.
Increases in gene expression of many structural components of the ECM, including collagen type I alpha 1, collagen type XII alpha 1, fibronectin 1, and laminin-5, were seen in the 20% strain group compared with the static, 0%, and 40% groups. Increases in ECM synthesis as a result of mechanical stimulation have been seen in a wide array of cell types. For example, an increase in ECM production, including collagen type I, was found in fibroblasts cultured on flexed membranes.39,40 Although the magnitude of gene upregulation in collagen type I alpha 1, collagen type XII alpha 1, fibronectin 1, and laminin-5 was small (between 1.3 and 2.8 times the control), the additive effect of these changes may have resulted in a stronger, more dense ECM. It has been proposed that increased ECM production is one of the contributing factors to the increase in mechanical strength seen in the 10% and 20% groups. However, it is unlikely that it is the sole contributing factor.
Much of the tensile strength of ES is attributed to a well-formed epidermis. 8 Within the epidermis the protein envelope of the cornified epidermis, consisting of structural proteins including involucrin, loricrin, and trichohyalin, imparts biomechanical strength to the epidermis.41,42 When loricrin is absent from the cornified envelope, the skin is more susceptible to damage by mechanical stress as seen in loricrin-deficient mice.43,44 Immunostaining for involucrin, a marker of epidermal differentiation, revealed no major differences in the amount or localization among the groups. Loricrin was either not seen or present in low quantities in the static, 0%, and 40% strain groups. In contrast, the 5%, 10%, and 20% groups had a thick continuous layer of loricrin in the upper epidermis in a pattern similar to NHS, suggesting appropriate terminal differentiation and epidermal barrier formation in vitro. A possible mechanism for changes in loricrin production in response to midlevel strain (5–20%) may be an increase in ion channel activity. Mechanical strain has been shown to increase ion channel activity, allowing Ca2+, K+, and Na+ cations to pass through these channels during intervals of stretch.45,46 Production of loricrin mRNA has been shown to be dependent on extracellular Ca2+ levels. 47 Increases in external Ca2+ concentration induce increases in intracellular Ca2+ concentrations which are required for keratinocyte differentiation. 48 Thus, mechanical stretch may activate Ca2+ ion channels, leading to an increase in intracellular Ca2+ and more rapid keratinocyte differentiation.
In contrast, the application of 40% strain to the ES did not result in a significant increase in loricrin production or an increase in mechanical properties. Immunostaining of the 40% ES for loricrin revealed that loricrin was present at far lower levels than the midlevel strains and exhibited noncontinuous, punctuate staining. A shift in balance between differentiation and proliferation in the early stages of ES culture may be responsible for this phenomenon. The application of 40% strain likely generated significantly greater tension than that of normal resting skin. When increased tension was applied to normal skin, such as through the use of a tissue expander, a complex series of events ensued, resulting in cell growth and tissue regeneration. 49 It is possible that the application of 40% strain to the newly forming ES caused the epidermal keratinocytes to proliferate greatly in order to restore physiologic levels of tension, as shown in Figure 4. The amount of proliferation required to return the 40% strained ES to physiological levels of strain (between 5% and 12% strain) would have been far greater than in the groups where midlevel strain was applied. This could result in a delay of keratinocyte differentiation and subsequently minimal amounts of loricrin production by culture day 10. This reduced level of differentiation within the epidermis coupled with lower ECM protein production could result in weaker ES.
The current data suggest that static strain during culture can improve the strength of ES. It is postulated that a combination of enhanced scaffold organization, increased production of ECM proteins as a result of upregulation of genes encoding structural ECM proteins, and enhanced epidermal differentiation is responsible for these observed improvements. Although the increased strength of ES may facilitate more facile surgical application and prevent damage during transplantation and dressing changes, additional studies are required to investigate the molecular mechanisms behind these changes to fully exploit the benefit of mechanical stimuli.
Footnotes
Acknowledgments
H.M.P. thanks the Shriners Hospitals for Children for a Postdoctoral Research Fellowship (grant no. 8450) and The Ohio State University for supporting these studies. The authors thank Deanna Leslie within the Histology Core Facility at the Shriners Hospitals for Children—Cincinnati for histological preparation and the Campus Electron Optics Facility at The Ohio State University for the use of their scanning electron microscope. This study was supported by grants from the Shriners Hospitals for Children (grant nos. 8507 and 8450).
Disclosure Statement
No competing financial interest exists.