
Abstract
We developed a method for controlling the spheroid formation of adult rat primary hepatocytes simply by optimizing the pillar diameters and patterns of nanopillar sheets. To investigate the effects of the pillar parameters on the spheroid formation, rat primary hepatocytes were cultured on nanopillar sheets with pillars that had one of five different diameters and that had been precoated with a solution containing one of two different concentrations of type I collagen. Spheroids with a compact morphology that were adhesive to the substratum and had an optimal size (50 to 100 μm) were obtained using a sheet with a pillar diameter of 2.0 μm that was precoated with 100 ng/mL of type I collagen solution. Immunohistochemistry revealed that the spheroids had a structure similar to that of native liver tissue. We then assessed the effect of overlaying reconstituted spheroids with Matrigel™ with the aim of achieving a simulated in vivo environment. The mRNA expression levels of MRP2, albumin, and P450-3A3 for spheroids determined by semiquantitative real-time PCR were significantly higher than those for spheroids cultured without the Matrigel overlay or for hepatocytes cultured using a conventional two-dimensional method. The spheroids obtained exhibited higher structural polarity and functional bile canaliculi compared with hepatocytes cultured using a conventional two-dimensional method.
Introduction
In contrast, primary hepatocytes maintain differentiated hepatocellular functions when they are induced to self-assemble into three-dimensional aggregates or spheroids by culturing on an ECM or substrate such as proteoglycan, 5 alginate scaffold, 2 poly(2-hydroxymethyl methacrylate), 11 titanium dioxide, 7 hollow fibers, 12 polyurethane foam, 13 and spinner flask. 14 Several investigators have reported that hepatocyte spheroids possess structural polarity and transporters similar to those of the bile canaliculi in the native tissue, which explains the enhanced hepatocellular activities of spheroids.2–4 Accordingly, the spheroid culture system is an alternative culturing technique for evaluating drug metabolism and toxicity in the course of drug screening.1,2 A great number of cell culture devices have been developed for forming three-dimensional structures (such as spheroids), including nanopillar (NP) sheets.7,15–18 In one case, NP sheets were fabricated with high aspect ratio structures with a pillar diameter of 160–1000 nm and a height of 280–1000 nm using nanoimprinting technology. 15
In the previous research on the effect of ECM patterns in the micrometer range during the initial phases of cell adhesion and spreading, Ingber's group found that the long-term effect of cell shape on growth and viability is not a consequence of the type of matrix protein but depends on the local geometric architecture of the substratum. 19 Bastmeryer et al. analyzed the effect of ECM patterns in the micrometer range by controlling the size of ECM squares (dots) and the distance between them. They found that a dot distance of around 2–5 μm affected the cellular shape and functions. 20
On the basis of these findings, we investigated how the NP surface structure affects cell adhesion, migration, and consequent spheroid formation. We used NP sheets with pillar diameters of 0.18, 0.5, 1.0, 2.0, and 5.0 μm and with pillar pitches (pillar center-to-center distance) that were twice the pillar diameter. The adhesion area of cells cultured on the NP sheets was restricted to the top of the pillars. This differs from culturing in a dish where the adhesion area is unrestricted. Accordingly, the mode of cell migration and the resultant morphology or aggregates for an NP sheet should differ from those for a conventional culture dish. From this point of view, our objective was to identify the optimal pillar diameters for spheroid formation in terms of cell adhesion and migration. We examined the structural polarity and biological functions of spheroids formed on NP sheets and investigated the effects of overlaying them with Matrigel on the hepatocellular functions.
Materials and Methods
NP sheet preparation
Polystyrene film with a thickness of 1.0 mm was spin-coated onto a glass substrate. A nanomold was made of silicon wafer and fabricated using photolithography. An NP structure was formed by pressing the mold onto the film at 423 K and then releasing it at room temperature. 15 The surface patterns of the fabricated NP sheets are illustrated in Figure 1.
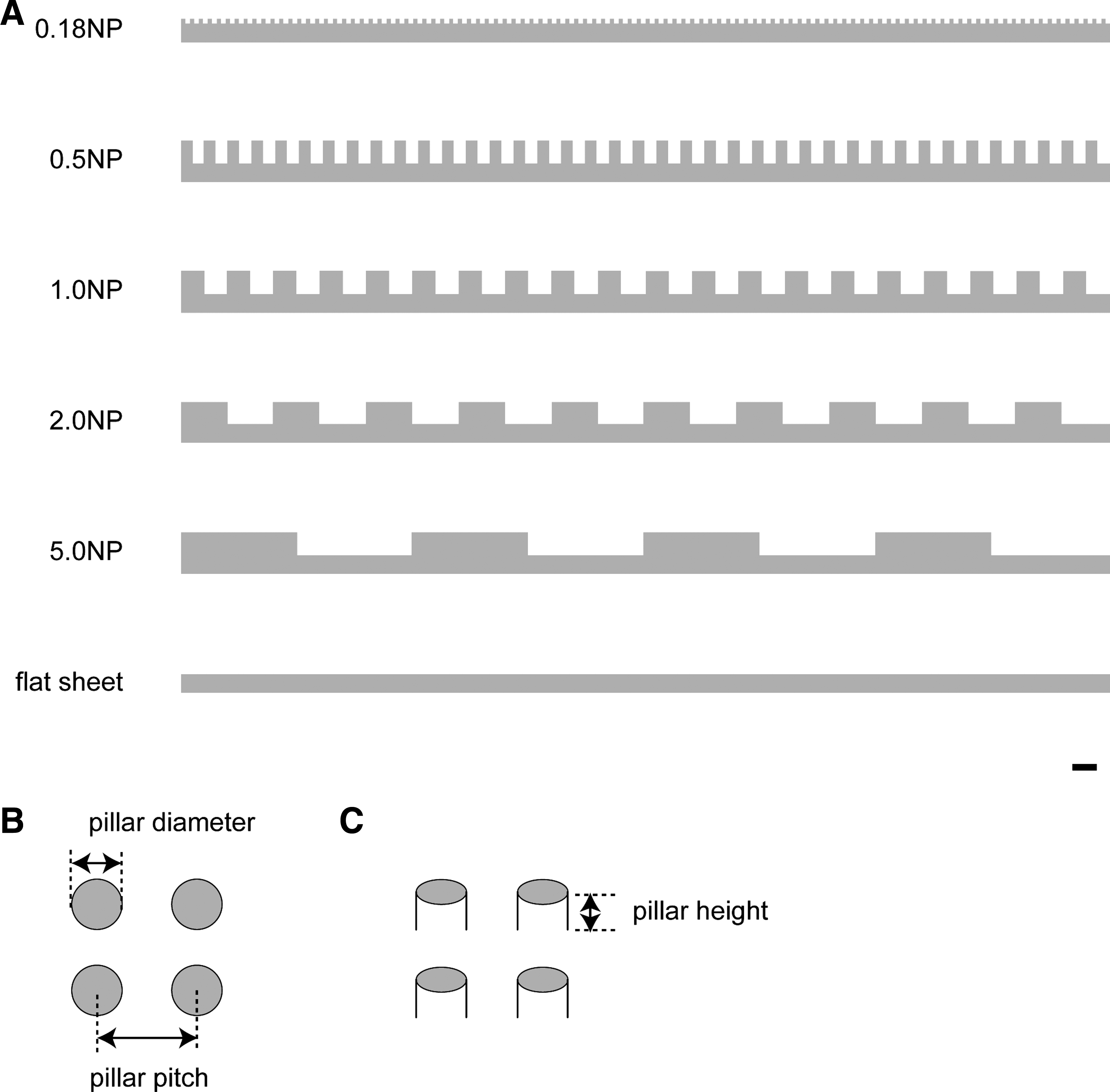
Surface patterns of nanopillar (NP) sheets used. (
Hepatocyte culture
Hepatocytes were isolated from 6- to 8-week-old specific viral pathogen–free male Wister rats (Charles River Japan Inc., Kanagawa, Japan) weighing about 150–250 g by using a modified two-step in situ collagenase perfusion method 21 and purified by isodensity Percoll centrifugation. 22 The viability of the hepatocytes was determined by trypan blue exclusion, and hepatocytes with over 85% viability were used for culturing.
The hepatocytes were resuspended in William's E medium containing 10% fetal bovine serum, supplemented with 8.6 nM insulin (Sigma-Aldrich Corp., St. Louis, MO), 255 nM dexamethasone (Nacalai Tesque, Inc., Kyoto, Japan), 50 ng/mL epidermal growth factor (Sigma-Aldrich Corp.), and 5 Kallikrein Inactivator Units (KIU)/mL aprotinin (Wako Pure Chemical Industries, Ltd., Osaka, Japan), and were seeded at a density of 1 × 105 cells/cm2 onto prepared NP sheets and into a 35-mm-diameter culture dish. The sheets were precoated with a solution containing 10 μg or 100 ng/mL of type I collagen. The type I collagen–precoated dish was purchased from AGC Techno Glass Co., Ltd., Chiba, Japan. The seeded hepatocytes were incubated in a humidified chamber with 5% CO2 at 37°C. Twenty-four hours after seeding, the medium for the NP sheet culturing was replaced with serum-free William's E medium containing the same supplements described above. The culture medium was changed daily. At the same time, the culture medium for the dish was replaced with serum-free William's E medium containing Matrigel (BD Bioscience, San Jose, CA) with the same supplements, which has been dubbed a sandwich culture.8,23 Forty-eight hours postseeding, the medium for the NP sheet culture was replaced with a serum-free medium containing Matrigel with the same supplements.
Immunohistochemistry
After a total of 96 h of culturing, the spheroids cultured on the NP sheets and the hepatocytes cultured in the dish were washed with phosphate-buffered saline (−) [PBS(−)] after removing the culture medium and then fixed in 4% paraformaldehyde in PBS(−) and permeabilized in 0.05% Triton-X in PBS(−). After blocking by fetal bovine serum in PBS(−), the samples were incubated in rabbit anti-human E-cadherin polyclonal antibody in a 100-fold dilution (SC-7870; Santa Cruz Biotechnology, Inc., Santa Cruz, CA), washed with 0.05% Tween 20 in PBS(−), and incubated in biotinylated anti-rabbit IgG antibody (BA-1000; Vector Laboratories, Burlingame, CA). The samples were then washed with 0.05% Tween 20 in PBS(−) and incubated in Streptavidin–Fluorescein (NEL720; PerkinElmer and Analytical Science, Inc., Waltham, MA). They were then washed again with 0.05% Tween 20 in PBS(−) and incubated in rhodamine phalloidin (R415; Invitrogen Corp., Carlsbad, CA). After incubation, the samples were washed with PBS(−) and incubated in Hoechst 33258. After a final washing in deionized water (DW), the samples were embedded in glass slides. Fluorescence images of them were taken with an optical microscope (AX70; Olympus Corp., Tokyo, Japan).
Resin embedding and microscopy
Rat hepatocytes cultured on the NP sheets were fixed with 2.5% glutaraldehyde in 0.1 M phosphate buffer( pH 7.4), incubated overnight at 4°C, and rinsed with 0.1 M phosphate buffer. Postfixation was performed with 1% osmium tetroxide in 0.1 M phosphate buffer (pH 7.4). After being washed with 0.1 M phosphate buffer and DW, the hepatocytes on the NP sheets were embedded in 1.5% SeaPlaque® agarose (Lonza Ltd., Basel, Switzerland). Agar-containing hepatocytes was dehydrated with a serial dilution of ethanol, treated with propylene oxide to dissolve the NP sheet, and embedded in epon-araldite mixed resin. Vertical sections of the samples were prepared using an ultramicrotome (Sorvall MT-6000; Du Pont Company, Wilmington, DE). Semithin sections (500 nm) were stained with toluidine blue and viewed with an optical microscope (AX70; Olympus Optical). Ultrathin sections (60 nm) were stained with uranyl acetate and lead citrate, and examined with a transmission electron microscope (H-7100; Hitachi Ltd., Tokyo, Japan).
Semiquantitative gene expression analysis
Total RNA was extracted using an RNeasy Tissue Kit (Qiagen GmbH, Hilden, Germany) every 24 h from rat hepatocytes cultured on NP sheets with and without Matrigel, from ones conventionally cultured as a monolayer, and from ones sandwich cultured. Two micrograms of the total RNA was reverse transcribed, and quantitative real-time PCR was conducted in a Thermal Cycler Dice Real Time System (Takara Bio Inc., Shiga, Japan) using a final reaction mixture of the RT product, TaqMan Gene Expression Assay (Applied Biosystems, Foster City, CA), Premix Ex Taq™ (Takara Bio Inc.), and DW. The 2−ΔΔCt method was used to calculate the relative change in the gene expression 24 ; TATA-binding protein was used as an endogenous control. The relative changes in expression were determined toward the control sample that freshly isolated (time 0).
Biliary excretion analysis
Every 24 h after cell culture started, biliary excretion assay was performed. Before each assay, the medium was washed by Hanks' balanced salt solution (Sigma-Aldrich Corp.) with calcium and magnesium. After that, it was replaced with a medium containing 5-carboxy-2′, 7′-dichlorofluorescein diacetate (CDFDA; Invitrogen Corp., Carlsbad, CA). After incubation at 37°C, the extracellular CDFDA was rinsed out by Hanks' balanced salt solution with calcium and magnesium. Fluorescence and phase-contrast images of the samples were acquired using an optical microscope (Axiovert 200; Carl Zeiss MicroImaging GmbH, Göttingen, Germany)
Results
Formation of rat hepatocyte spheroids on NP sheets
To examine the effect of the NP diameter on spheroid formation, hepatocytes were seeded on NP sheets with a pillar diameter of 0.18, 0.5, 1.0, 2.0, or 5.0 μm, which are referred to as 0.18NP, 0.5NP, 1.0NP, 2.0NP, and 5.0NP, respectively. A culture dish without NP (flat sheet) was used as a control. Side views of their surfaces are shown in Figure 1A. The pillar diameter, pitch, and height were defined as shown in Figure 1B and C, and the dimensions are summarized in Table 1. To examine the effect of the amount of type I collagen in the coating solution on spheroid formation, we observed spheroid morphology at 96 h postseeding. Since hepatocytes are conventionally cultured in a culture dish precoated with a solution containing 1–1.5 mg/m of type I collagen,9,10 we tested a sequential 100-fold dilution series: 1 ng/mL, 100 ng/mL, 10 μg/mL, and 1 mg/mL. In preliminary experiments, a spheroidal morphology was observed for both 1 and 100 ng/mL, and a flat, monolayer morphology was observed for 10 μg/mL and 1 mg/mL (data not shown). Accordingly, in this study, we used 100 ng/mL as representative of spheroidal morphology and 10 μg/mL as representative of flat, monolayer morphology. In experiments using these different NP sheets and solutions with different concentrations of type I collagen, we observed spheroids with a compact morphology in which individual cells could barely be distinguished on the 0.18NP, 2.0NP, 5.0NP, and flat sheets coated with a solution containing 100 ng/mL type I collagen (Fig. 2B, K, N, Q). Intermingled monolayer hepatocytes and spheroids were observed on the 0.5NP and 1.0NP sheets with the same concentration of collagen (Fig. 2E, H). Similar results were observed for the uncoated NP and flat sheets. However, unlike the spheroids on the NP sheets coated with 100 ng/mL collagen solution, the spheroids on the uncoated 0.18NP, 2.0NP, 5.0NP, and flat sheets (Fig. 2A, J, M, P) had weak adhesion to the NP sheets. The number of spheroids decreased over time due to the changing of the culture medium. Moreover, the spheroids were exactly like cell aggregates, which do not have a compact morphology as a whole. Rat hepatocytes were spread on NP sheets and flat sheet coated with 10 μg/mL of type I collagen solution in monolayer configuration for all pillar diameters (Fig. 2C, F, I, L, O, R). This revealed that the hepatocyte and spheroid morphologies depended on the pillar diameter and/or the concentration of type I collagen. Spheroids with a compact morphology adhered to the substratum were accomplished on 0.18NP, 2.0NP, 5.0NP, and flat sheets coated with a solution containing 100 ng/mL of type I collagen.
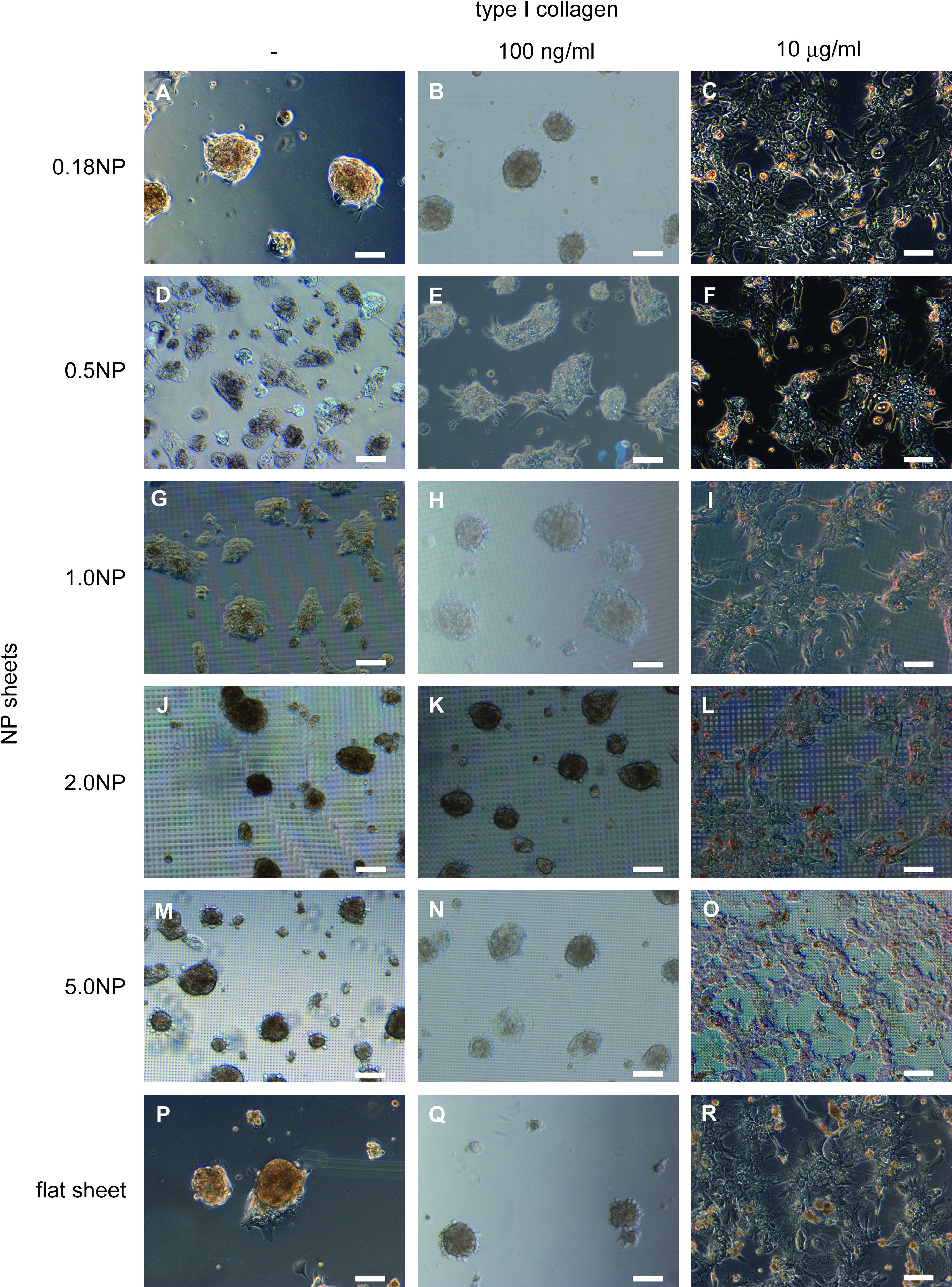
Phase-contrast micrographs of hepatocyte cultures on NP sheets 96 h after seeding. Hepatocytes were cultured on 0.18NP (
NP, nanopillar.
To determine the optimal size of pillar diameter for spheroid formation, we calculated the number of spheroids and the distribution of their diameters on the basis of micrograph data for spheroids that formed on NP sheets coated with a solution containing 100 ng/mL of type I collagen. Most of the spheroids on the 0.18NP, 2.0NP, and flat sheets were 50–100 μm in size, and most of those on the other NP sheets were 100–150 μm in size (Fig. 3). As shown in Table 2, the 2.0NP sheet had the most spheroids (153.6) and the smallest standard deviation (±5.3), indicating that spheroids of relatively uniform size formed on the 2.0NP sheet. Since spheroids up to 100 μm in diameter have higher viability or effective cellular function, 25 2.0NP is suitable for formation of spheroids because both the number (58.1 ± 13.3) and ratio (37.8%) of spheroids on the 2.0NP sheet ranging in size from 50 to 100 μm were the highest among the NP sheets (Fig. 3) and because the number of 1–100 μm spheroids on the 2.0NP sheet versus the other NP sheets was consistently larger with statistically significant differences (p < 0.05, Table 3). Consequently, 2.0NP sheets coated with a solution containing 100 ng/mL of type I collagen were used in the subsequent experiments.

Size distribution of hepatocyte spheroids. Hepatocytes were cultured on 0.18NP, 0.5NP, 1.0NP, 2.0NP, and 5.0NP sheets and on a flat sheet coated with a solution containing 100 ng/mL of type I collagen solution. Spheroids were counted 96 h postseeding.
The recruitment of E-cadherin and actin to regions of intercellular contact is essential for the formation and stabilization of adherens junctions, resulting in appropriate tissue structure. 26 Accordingly, the localizations of E-cadherin and actin were investigated using immunohistochemistry to compare the structure of hepatocyte spheroids with native liver tissue. Figure 4 shows double-staining images for a frozen section of rat native liver, an NP-cultured spheroid, and a monolayer-cultured hepatocytes. Rhodamine-phalloidin was used to stain the actin (red), and anti-E-cadherin antibody was used to stain the E-cadherin (green). The E-cadherin of the rat liver and the spheroids formed on the NP sheet were highly concentrated at the cell–cell junction (Fig. 4D, E) as was the actin (Fig. 4A, B). In contrast, the E-cadherin from the conventional monolayer culture was below the limit of detection (Fig. 4F), and actin stress fibers were observed (Fig. 4C).

Immunohistological staining of actin and E-cadherin for hepatocytes cultured on 2.0NP and in conventional collagen-coated dish for 96 h. Frozen section of rat native liver, hepatocyte spheroids on NP sheets, and hepatocyte monolayer cultured in dish were stained with rhodamine-phalloidin (red:
Effects of ECM overlay on hepatocyte spheroids
Structural analysis
It is well established that an ECM can modulate gene expression in many biological systems, and it has been suggested that a complex ECM, but not its purified components, is needed to maintain differentiated hepatocytes in vitro for a long period. 27 Accordingly, we expect the effect of ECM, which simulates the environment of a living organism by surrounding the cells, and overlaid Matrigel, which includes complex basement membrane extract but not purified components, on the spheroids formed by NP culturing.
To observe the morphology and examine cellular viability before and after overlaying the Matrigel, we cultured rat hepatocytes on a 2.0NP sheet coated with a solution containing 100 ng/mL of type I collagen. Semithin sections of reconstituted spheroids were stained with toluidine blue. Rat native liver tissue and sandwich-cultured hepatocytes were also examined for comparison. In the sandwich culture system, monolayer-cultured hepatocytes are held between two kinds of thin-layered ECM components, and in the NP culture system with Matrigel, Matrigel is overlaid on spheroids formed on the NP sheets. The hepatocytes cultured without the Matrigel overlay had a variety of shapes but were spheroidal overall (Fig. 5A, B) while those cultured with Matrigel were individually round polygons (Fig. 5C), much like those of native liver (Fig. 5E). The sandwich-cultured hepatocytes were uniformly flat (Fig. 5D). These results indicate that the use of a Matrigel overlaying onto spheroids is more effective than the use of monolayer culturing for recovering the native morphology. The viability of the hepatocytes was then examined. The ratio of dead hepatocytes represented by asterisks in Figure 5 was increased time dependently under the condition of without the Matrigel overlay (Fig. 5A, B; 48 and 96 h postseeding). Almost all the hepatocytes of spheroids cultured with the Matrigel overlaid at 48 h were still alive at 96 h (Fig. 5C). The viability of the hepatocytes was thus improved by the Matrigel overlay (Fig. 5A–C). The bile canaliculi that grew in the Matrigel spheroids were 5–10 μm in diameter (Fig. 5C), larger than those that grew in the non-Matrigel spheroids (Fig. 5A, B).

Toluidine blue staining of hepatocyte spheroid sections on 2.0NP (
To investigate the bile canaliculi structures and their peripheral parts in more detail, we used a transmission electron microscope (TEM; H-7100; Hitachi Ltd.). We found that bile canaliculi divided by a tight junction formed in all the reconfigured structures: spheroidal (Fig. 6A–C) and sandwich (Fig. 6D). Bile canaliculi with well-developed microvilli those were in native tissue (Fig. 6E) were observed in spheroids cultured with and without Matrigel (Fig. 6A, C). Bile canaliculi with and without well-developed microvilli were observed in spheroids cultured with Matrigel (Fig. 6B, C), but only bile canaliculi without well-developed microvilli could be observed in sandwich cultured (Fig. 6D).

Transmission electron microscopy (TEM) images of ultrathin section of hepatocyte spheroids cultured on 2.0NP for 96 h without Matrigel overlay (
Semiquantitative gene expression analysis
The real-time PCR was performed to measure the relative amounts of gene expression among four kinds of culture conditions. Hepatocytes of soon after isolating were used as the calibrator sample, which is indicated as time 0 in Figure 7. Expression of MRP2 tended to increase with time except for the monolayer culture, and that at 96 h for spheroids cultured on NP sheets with and without Matrigel was higher than that for the sandwich- and monolayer-cultured hepatocytes (p < 0.05) (Fig. 7A). Expression at 72 h for spheroids cultured on the NP sheet with Matrigel overlay at 48 h was substantially higher than that for those cultured without Matrigel. However, the final values for these expressions at 96 h were virtually the same and were close to the initial level at time 0 (Fig. 7A). Although a substantial increase was also observed during the next 24 h after Matrigel overlay for the sandwich culture, a further increase over time was not observed.
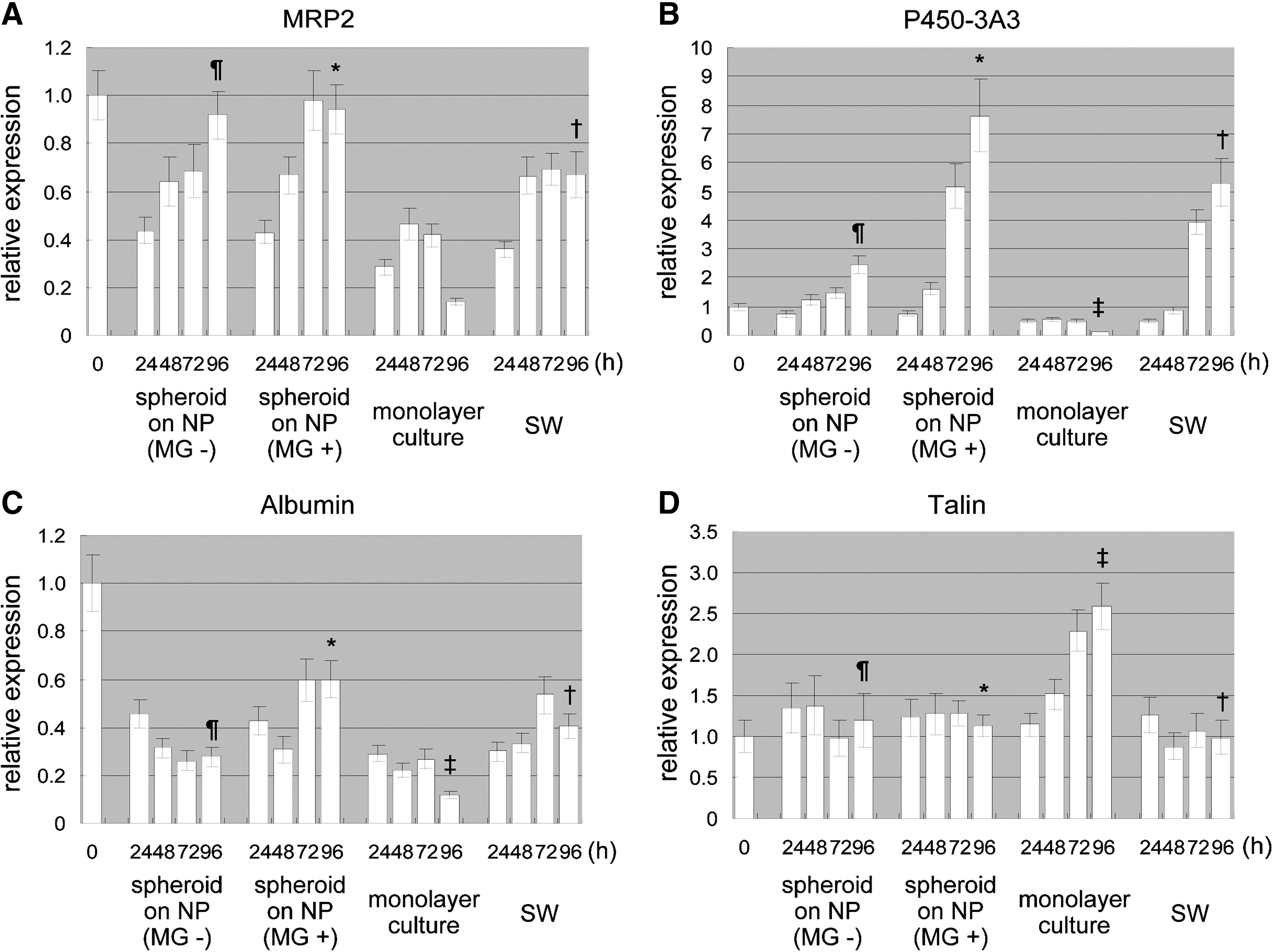
Semiquantitative real-time PCR. Relative amounts of mRNA expression of MRP2, P450-3A3, albumin, talin, and E-cadherin were measured every 24 h under four culture conditions: culture on 2.0NP without Matrigel (MG−), culture on 2.0NP with Matrigel overlaid at 48 h postseeding (MG+), monolayer culture in collagen-coat dish (monolayer culture), and sandwich culture (SW). Expression values of each gene were compared to control hepatocyte sample (time 0, before seeding). Statistical comparison was performed for 96-h samples for each culture condition. (
Under all conditions except for the monolayer culture, the expression level of cytochrome P450-3A3 (P450-3A3) increased over time (Fig. 7B). Although the expression level for spheroids cultured on the NP sheet with Matrigel overlay took 24 h after the overlay to substantially increase, that for the sandwich culture took 48 h; that is, there was a time lag between these two culture conditions. Comparison of the expressions at 96 h revealed that the one for spheroids cultured on the NP sheet with Matrigel was much higher than the others (p < 0.05).
Although albumin expression levels for spheroids cultured on the NP sheets and the monolayer-cultured hepatocytes tended to decrease during the first 48 h, those for spheroids cultured on the NP sheet with Matrigel increased after Matrigel overlay at 48 h (Fig. 7C). The expression level for the sandwich-cultured hepatocytes trended upward for the first 72 h and then turned downward. Comparison of the expression levels at 96 h showed that the one for spheroids cultured on the NP sheet with Matrigel was much higher than that for the others (p < 0.05). However, the expression levels at 96 h were at most around 60% of the initial level.
The expression patterns for talin differed from those for the three other genes. The expression level for the monolayer-cultured hepatocytes increased over time, resulting in a 2.5-fold higher level at 96 h than the initial level (Fig. 7D). In contrast, for the other three conditions, the expression levels remained fairly constant, and the levels at 96 h were substan-tially lower than that for the monolayer culture (p < 0.05).
The expression patterns for E-cadherin for spheroids cultured on the NP sheet with Matrigel showed a substantial increase in a 24 h period after Matrigel overlay at 48 h (Fig. 7E). For the sandwich-cultured hepatocytes, expressional induction caused by Matrigel overlay was not showed within a 24 h. Although the expression level for spheroids cultured on the NP sheet with Matrigel at 72 h was higher than that for spheroids cultured on the one without Matrigel and the sandwich-cultured hepatocytes, the expression levels at 96 h were not significantly different at p < 0.05 for all conditions except monolayer culture.
Biliary excretion
To examine whether MRP2 works as a hepatocyte efflux transporter, a capability revealed in a previous gene expression study (Fig. 7), a biliary excretion assay was conducted under two culture conditions: NP and sandwich. Carboxydichlorofluroscein diacetate (CDFDA), a substrate of MRP2, 28 was used to observe the canalicular uptake and efflux. Fluorescence and phase-contrast micrographs were taken with an optical microscope every 24 h (Fig. 8). Analysis of the micrographs revealed that CDFDA had accumulated inside both the NP-cultured spheroids and sandwich-cultured hepatocytes for 24 h (Fig. 8A, C). Subsequently, CDF gradually accumulated inside both types of cells, and bile canaliculi formed in a time-dependent fashion (Fig. 8E, G). Almost all the CDF were excreted into the canalicular networks in the final 48 h (Fig. 8I, K, M, O, Q). In comparison to those in the spheroids formed by culturing without Matrigel (Fig. 8Q), the bile canaliculi that reconstituted in the spheroids formed by culturing with Matrigel (Fig. 8M) were longer and more numerous.

Biliary excretion assay. Hepatocytes were cultured on 2.0NP with or without Matrigel and in sandwich configuration. 5-Carboxy-2′, 7′-dichlorofluorescein diacetate was used to enable canalicular uptake and efflux to be observed. Phase-contrast micrographs and fluorescent micrographs were taken every 24 h. Matrigel for NP sheets was overlaid at 48 h postseeding. (
Discussion
In this study, hepatocyte spheroids with a compact and homogenous morphology in which the hepatocytes were individually round polygons, a configuration close to that of those in native liver, were formed by culturing with the NP sheets although the extent of spheroid formation depended on the pillar diameter. Experiments showed that spheroid formation could be adjusted by altering the pillar pitch and diameter (Fig. 2). The downregulation of cell–substratum adhesion-related molecules of talin that is capable of linking integrins to the actin cytoskeleton, 29 were observed to form spheroids (Fig. 7). It is conceivable that the pillar diameter modifies the cell–cell or cell–substratum adhesion force and thus the extent of cell migration, resulting in enhanced spheroid formation. Further study is needed to clarify the relationship among the pillar diameter, expression and localization of cell–cell and cell–substratum adhesion molecules, and the signal transductions mediated by cellular adhesion and small GTPase that hydrolyze guanosin triphosphate (GTP) such as RhoA 30 and for detailed understanding about the mechanism of pillar-diameter-dependent spheroid formation.
Spheroid formation has been observed through many experiments by use of substrates with different surface physicochemical properties and precoated with various proteins or biological materials.1,2,5,7,11–14,16–18 Although these experiments showed that spheroid formation achieved positive results regarding hepatocellular functions, there were differences in spheroid characteristics. For example, although alginate biomaterial is convenient to prepare with high guluronic content by freeze-drying, the scaffold-seeded hepatocytes that did not aggregate and form spheroids underwent apoptosis because of weak adhesiveness. 2 In another case, the culture substratum of a titanium dioxide gel surface was formed simply by spin-coating and hydrolysis; however, the number of cells gradually decreased during the culturing owing to the detachment of hepatocyte spheroids from the surface, and the diameter of the spheroids increased with culture time due to fusion of floating spheroids. 7 Controlling the spheroid diameter is another critical issue to avoid the decrease of cellular viability or metabolic activity in terms of oxygen limitation at the center of spheroid. Culturing using microimprinting technology 18 or a two-dimensional microarray technique 16 is more effective because spheroids are immobilized at a defined location and their diameter can be controlled by defining cell adhesion and nonadhesion areas. On the other hand, although it was also observed in the NP culture system that the number of hepatocytes or spheroids gradually decreased as the medium was changed because of weak adhesiveness to the substratum, this was improved by optimizing the pillar diameter in our experiments (Figs. 2 and 3). In this experiment, as 2.0NP had relatively uniform-sized spheroids of 1–100 μm (Table 3) and its ratio is 50.1%, which is apparently higher than that of other four kinds of NP sheets (Fig. 3), there is no need to immerse them in a chemical solution such as polyethylene glycol.16,18 Moreover, since the pillar height of the NP sheet can be controlled, the required ingredients and oxygen for cell viability are supplied from the underside hepatocytes of spheroids attached to the top of the pillars. Accordingly, these results indicate that the NP sheets have an inherent potential to not only control the spheroid diameter simply by optimizing the pillar diameters but also make uniform the hepatocellular functions of each spheroid. It is reported that the no-oxygen limitation will take place in spheroids up to 100 μm in diameter. 25
The hepatocytes of spheroids cultured on the NP sheet with Matrigel were round polygons, a morphology close to that of those in native liver (Fig. 5). This is because the hepatocytes were surrounded by the ECM of the Matrigel, resulting in the recovery of primary polarity by simulation of the native environment. One of the representative molecules indicating recovery is MRP2, which is a canalicular multispecific organic anion transporter and located exclusively in the canalicular membrane. 31 The upregulation of the MRP2 of spheroids that cultured on the NP sheet with Matrigel and that confirmed by gene expression analysis supports the positive effect of Matrigel on the recovery of primary hepatocellular polarity. The cells that did not contact the Matrigel directly showed a polygonal morphology (Fig. 5). This indicates that intracellular signals were transmitted to all the hepatocytes constituting the spheroid. This phenomenon is considered to reflect the native structure, which supports a cuboidal cell shape, distinct polarity, and a three-dimensional cellular communication system. 18 A polygonal morphology was not observed for the hepatocytes sandwich cultured with a Matrigel overlay. This may be because the hepatocytes were cultured in higher concentrations of type I collagen than those cultured on the NP sheet. As a result, the cell–substrate adhesion force was stronger, causing the hepatocytes to be uniformly flat rather than polygonal. The use of a Matrigel overlay is thus effective for reconstituting morphologies similar to that of hepatocytes in native liver. It is effective when overlaid on spheroids and not on structures with a flat morphology like those formed by sandwich culturing.
Bile canaliculi with well-developed microvilli, like those in native tissue, were observed in the three-dimensional structures that were formed, that is, spheroids (Fig. 6). However, the bile canaliculi of the spheroids cultured on the NP sheet with a Matrigel overlay and of the hepatocytes sandwich cultured with a Matrigel overlay tended to be larger, and microvilli were not detected (Fig. 6). The presence of large-bore bile canaliculi in spheroids formed using Matrigel was confirmed by the results of biliary excretion assay (Fig. 8). This is because reconstituted in vitro bile canaliculi were the closed structure where the excreted metabolites were accumulated. The uptake and efflux of the metabolites became even more active due to the Matrigel overlay, and eventually the closed bile canaliculi became larger. Since accumulated metabolites harm the hepatocytes and decrease cell viability,2,32 a method is needed to prevent this accumulation to obtain a more effective system for evaluating drug metabolism and toxicity.
The use of real-time PCR enabled us to quantitate the amount of expressed gene. The MRP2 expression levels of the spheroids cultured on NP sheets with and without Matrigel were higher than for those of the sandwich- and monolayer-cultured structures (Fig. 7). This indicates that spheroid formation on an NP sheet is effective for MRP2 expression. P450 is a ubiquitous superfamily of heme-containing monooxygenase enzymes that play a fundamental role in the metabolism of chemically diverse compounds, including endogenous chemicals and pharmaceutical agents.33,34 Serum albumin is a major protein synthesized almost exclusively by liver cells in animals, and its concentration increases from low levels early in fetal development to a high plateau level in adults. 35 Both P450 and serum albumin are major indicators of hepatocellular function. The expression levels of these two genes were the highest for the spheroids cultured with Matrigel (Fig. 7), suggesting that overlaying Matrigel on three-dimensional structure (spheroids) is more effective on transcription level of the two genes than overlaying that merely on two-dimensional hepatocytes (sandwich culture). In contrast, the expression level of talin was the highest for the monolayer-cultured hepatocytes (Fig. 7). These findings are consistent with those of the immunohistochemical analysis (Fig. 4), which showed that, with monolayer culturing, the actin formed stress fibers, unlike spheroids cultured on NP sheets, which means that the talin was overexpressed. 36 The expression level of E-cadherin of the spheroids cultured with Matrigel at 72 h was higher than other culturing conditions at 72 h (Fig. 7). This indicates that Matrigel has a faster effect on E-cadherin expression due to overlaying on spheroids and not flat structures as was done in the sandwich culturing. There was no particular culturing condition showing a particularly high expression of E-cadherin at 96 h among the NP culturing with, without Matrigel, and sandwich culturing (Fig. 7). It has been reported that E-cadherin expression needs to form three-dimensional structures such as spheroids,37,38 which is consistent with our findings (Fig. 7).
In conclusion, the present results suggest that the spheroid formation of rat primary hepatocytes was controlled by optimizing the pillar diameters of NP sheets and that the combination of spheroid formation and Matrigel overlaying has positive effects on hepatocyte viability, polarity, and functions. The present study demonstrates that it is practically possible to control the formation of spheroid only by optimizing the physical property of substrate surface. Our next challenge is controlling the diameter of spheroids, which may be effective in regulating their biological function. It is an essential factor to be considered in the development of not only high-throughput but also higher-reproducible and reliable in vitro assay systems to evaluate the drug metabolism and toxicity.
Footnotes
Acknowledgments
The authors thank Shigeharu Nishiuchi, Masabumi Nemoto, and Taku Saito for supplying the NP sheets and their helpful comments and suggestions.
Disclosure Statement
No competing financial interests exist.
This work was performed at Advanced Research Laboratory, Hitachi Ltd., Hatoyama, Saitama, Japan.