
Abstract
We have investigated differences in bovine limbal epithelial cell differentiation when expanded upon intact (amniotic epithelial cells and basement membrane remaining) and denuded human amniotic membrane (AM), a commonly used substrate in ophthalmic surgery for corneal stem cell transplantation. Ex vivo expansion of the epithelial cells, in supplemented media, continued for 2 weeks followed by 1 week under air-lifting conditions. Before and after air-lifting the differentiated (K3/K12 positive) and undifferentiated (K14 positive) cells were quantified by immunohistochemistry, Western blotting, and quantitative polymerase chain reaction. Limbal epithelial cells expanded upon AM formed 4–6 stratified layers, both on intact AM (iAM) and denuded AM (dAM). On dAM the proportion of differentiated cells remained unaltered after air-lifting. Within cells grown on iAM, however, the number of differentiated cells increased significantly after air-lifting. These results have important implications for both basic and clinical research. First, they show that bovine limbal epithelia can be used as an alternative source of cells for basic research investigating ex vivo limbal stem cells expansion. Second, these findings serve as a warning to clinicians that the effect of AM on transplantable cells is not fully understood; the use of iAM or dAM can produce different results in terms of the amount of differentiation, once cells are exposed to the air.
Introduction
The cornea is often described as our window to the world and its transparency is paramount. Continual replenishment of corneal epithelial cells, which cover the outer surface of the eye, is critical if the cornea to maintain its transparency. Corneal stem cells are involved in the epithelial cell renewal and repair function. These stem cells are believed to be concentrated in the limbus, the outer vascular rim at the junction between the cornea and the conjunctiva,17,18 and are termed limbal epithelial stem cells (LESC). LESC share a number of features with other adult somatic stem cells, including small cell size, 19 a lack of expression of differentiation markers such as cytokeratin 3/12,17,20 and a high nuclear-to-cytoplasmic ratio. 21 However, although much effort has been made to identify potential molecular markers to distinguish LESC, such as P63, 22 ABCG2, 23 and cytokeratin 14, 24 there is still no definitive marker. 25 Integrin α6 is a cytoplasmic protein that plays an important role in cell–extracellular matrix adhesion,26,27 and since its distribution is located to the basal layers it is believed to be a potential stem cell marker. 25
For the ex vivo cultivation and expansion of limbal stem cells, AM, as a substrate for therapeutic stem cell delivery, has been shown to offer excellent support.2,4,7,14,15 As well as a substrate in the treatment of limbal stem cell deficiency, AM has also proven to be clinically useful in the management or program of various ocular surface disorders, including chemical/thermal burn injuries to the cornea.28–30 However, there are concerns about how to achieve the best preparation of AM before its clinical use,9,11,31 including our own recent study that has shown how structural differences might impact upon its therapeutic effect. 32
When considering methods to prepare AM for ex vivo limbal stem cell expansion and subsequent stem cell transplantation, the main debate is whether or not to denude AM of its native epithelial cells (and by default its basement membrane) before the expansion of donor limbal epithelial cells. While denuding the AM results in better cell stratification and attachment, 11 an intact AM (iAM) is thought to maintain an active stem cell niche for cultured corneal epithelial cells, thereby retaining the expanded limbal cells in an undifferentiated state.9,31 In this article we seek to investigate this point further. Specifically, by comparing limbal epithelial cell differentiation on denuded AM (dAM) and iAM before and after air-lifting, we aim to show the significance of AM epithelium/basement membrane removal on cell growth.
Air-lifting, as a cell culture technique, was originally developed to create sheets of skin cells for transplantation and to promote epidermal cell differentiation,33–36 and this technique has since successfully been applied to corneal cell culture.12,37 The established method for preparing AM for corneal cell culture is to manually remove the AM epithelium; however, this has recently been shown to remove not just the cells but also the basement membrane. 38 Therefore, the present study is not just a comparison between the presence (iAM) and absence (dAM) of amniotic epithelia, but also a comparison between the presence and absence of the basement membrane on bovine limbal stem cell differentiation before and after air-lifting.
Materials and Methods
Preparation of human AM
Cryopreserved human AM was collected from the Division of Ophthalmology and Visual Science, Queens Medical Centre, Nottingham, United Kingdom, with approval from the Local Research Ethics Committee (Nottingham). The membranes were washed with sterile phosphate-buffered saline (PBS) containing penicillin (100 IU/mL) and streptomycin (100 μg/mL) (Gibco) within a class II microbiology safety cabinet, before they were cut into pieces (∼4 cm2), and immediately stored at −80°C in PBS. Before use, the AM was thawed and washed in sterile PBS. dAM was obtained by incubating the iAM in 0.1% ethylenediaminetetraacetic acid (Invitrogen) at 37°C for 1 h to lessen cellular adhesion, before gentle scraping of the upper cell layer. iAM and dAM were positioned with either native epithelium or basement membrane side up within transwell inserts held inside six-well culture plates (Corning).
Scanning electron microscopy
Both iAM and dAM were fixed using 2.5% glutaraldehyde in PBS, washed three times for 10 min in PBS, and postfixed for 2 h in 1% aqueous osmium tetroxide. Fixed AM were then washed three more times in PBS before they were dehydrated within a graded ethanol series (50%, 70%, 90%, and 100%). AM samples were immersed in hexamethyldisilazane (Sigma) for 20 min, allowed to air-dry, and mounted on aluminum stubs. They were then sputter coated with gold before examination using an environmental scanning electron microscope (FEI Quanta FEG 600).
Culture of bovine limbal epithelial cells
Bovine cornea has a structure and biochemical makeup similar to human cornea. Like in human cornea, bovine corneal stem cells have been localized to the basal layers of the limbus, 24 and have successfully been used as an alternative source of human limbal stem cells for basic research. 39 Bovine eyes were collected (Chity whole-sale abattoir; Guildford) within 2 h of death and transported on ice before corneas were dissected for epithelial cell isolation. The whole limbal ring tissue from each eye, including the subconjunctival Tenon's capsule, was cut into 10–12 pieces (∼5 mm long) and incubated for 12 h at 37°C in a basal culture medium with 0.02% type IA collagenase (Sigma-Aldrich).
The epithelium was peeled from the enzyme-treated limbal pieces, and incubated with 0.05% trypsin–ethylenediaminetetraacetic acid for 10 min at 37°C, and the cell sheets were dissociated into single cells by agitation through a 22-gauge needle. The suspension of corneal limbal epithelial cells was placed in to supplemented media (Dulbecco's modified Eagle's medium and Ham's F12 (1:1) (Gibco) including 5% fetal bovine serum, 2 ng/mL human epidermal growth factor, 5 μg/mL insulin, B27 supplement40,41 medium, and penicillin streptomycin (Invitrogen), and seeded onto either iAM or dAM (spread on the bottom of culture inserts). AM and cells were submerged in supplemented media for 2 weeks and then exposed to air by lowering the medium level (air-lifting) for 1 week to promote corneal epithelial cell differentiation. Cultures were incubated at 37°C under 5% CO2 for ∼3–4 weeks; the medium was replaced every 2 days.
Immunohistochemistry
Limbal epithelial cells expanded upon AM were collected using a 6 mm biopsy (Brymill) before (2 weeks) and after (3 weeks) air-lifting, embedded in tissue Tek O.C.T. compound (Agar Aids), snap-frozen in liquid nitrogen, and stored at −80°C before being sectioned. Cryostat sections (7 μm thick) were placed onto poly-lysine coated slides and air-dried for 2 h. Slides were then fixed in 100% methanol at −20°C for 15 min and in acetone (−20°C) for 5 min before they were washed in PBS. Tissue sections were incubated with 1% goat serum or bovine serum albumin at room temperature for 30 min to block nonspecific binding.
Blocked sections were incubated in anti-K14 (1:100, guinea Pig, polyclonal; Progen), anti-K3 (1:50, mouse, monoclonal; Chemicon), anti-Integrin α6 (1:100, mouse, monoclonal; Progen), and anti-laminin (1:100, mouse monoclonal; Progen) primary antibodies overnight at 4°C and washed for 5 min in PBS. Horse anti-mouse IgG (1:50; Vector Labs)/Goat anti-guinea pig (1:100; Progen) secondary antibodies conjugated to fluorophores were applied for 60 min at room temperature. Unbound secondary antibodies were removed by washing in PBS and sections were mounted under glass cover slips using 4′,6-diamidino-2-phenylindole (DAPI) mounting media (Vector Labs). Negative controls were performed by replacing the primary antibody with PBS. Mounted sections were examined by fluorescence microscopy (Imager A1; Zeiss).
Isolation of RNA and cDNA synthesis
Total RNA was isolated from cells cultured on both iAM and dAM (n = 6), from fresh bovine cornea (central and limbal region, n = 4) as well as from iAM and dAM (n = 2) using the TRI reagent (Sigma), according to the manufacturer's protocol. Total RNA was quantified spectrophotometrically (GE Healthcare), and 1 ng RNA was reverse transcribed using RevertAid H Minus First Strand cDNA synthesis Kit (Fermentas), following to manufacturer's protocol.
Real-time quantitative polymerase chain reaction
A custom-made PerfectProbe assay (Primerdesign) was used to quantify Keratin 12 gene (accession number: XM_001255461), GAPDH (accession number: NM_001034034), and SDHA (accession number: NM_174178) expression. Each reaction was performed in triplicate on three different replicate samples with a final reaction volume of 20 μL containing 10 μL 2 × quantitative polymerase chain reaction (qPCR) Mastermix (Primerdesign), 1 μL reconstituted perfectprobe primer–probe mix (Primerdesign), 4 μL PCR-Grade water (Primerdesign), and 5 μL cDNA (1:10 of original concentration). Nontemplate controls were also run. Real-time reactions were performed within a 96-well plate (Fisher) in the ABI PRISM 7700 Sequence Detector (Applied Biosystem). The real-time PCR data were analyzed by ΔΔCT method; therefore, the K12 mRNA expression level was relative to GAPDH and SDHA expression levels.
Western blotting
Proteins from bovine limbal epithelial cells, AM alone and epithelial cells grown on iAM and dAM with and without air-lifting (4 μg total protein for each condition; estimated using the modified Lowry assay), were separated by one-dimensional sodium dodecyl sulfate–polyacrylamide gel electrophoresis using 10% gels. They were transferred to polyvinylidine difluoride membranes, and nonspecific binding to membranes was blocked by incubation with 5% (w/v) milk dissolved in 1 × Tris-buffered saline-Tween (TBS-T) (20 mM Tris-base, 0.14 M NaCl, 0.1% Tween®-20; pH 7.6). Membranes were incubated with mouse monoclonal anti-K3 and anti-GAPDH primary antibodies (1 μgmL−1) diluted in 2% (w/v) milk dissolved in 1 × TBS-T at 4°C overnight. Blots were washed for 45 min in 1 × TBS-T before incubation with an anti-mouse HRP-conjugated secondary antibody (1:6000 dilutions) for 2 h at room temperature. Proteins were detected on X-ray film using an enhanced chemiluminescence system.
Statistical analysis
Student's t-tests (unpaired) were performed using Microsoft Excel. Results are presented as the mean of three individual experiments with standard error of mean, and p-value ≤0.05 was considered significant.
Results
Scanning electron microscopy of iAM and dAM
Scanning electron microscopy was performed on iAM and dAM before its use as a limbal stem cell substrate. Clear differences in the surface structure were seen between dAM and iAM (Fig. 1). The native epithelial cells were closely packed on the iAM surface (Fig. 1A, C). After manual removal of the native AM epithelial cells and basement membrane, collagen fibers were exposed (Fig. 1B, D).

Scanning electron microscopy micrographs of AM (iAM: left column; dAM right column). The native devitalized AM epithelium on the iAM sample remains attached. The AM epithelial cells were closely packed forming a barrier covering the AM surface (
Morphology of limbal-cultured suspended epithelial cells
Bovine limbal epithelial cells were successfully cultured on both dAM and iAM for 3 weeks (2 week before air-lifting plus 1 week after air-lifting) (Fig. 2). Within the resultant culture, two cell types were observed on iAM and dAM. Before air-lifting (Fig. 2A, B), large spindle-shaped cells dominated the surface layer of both iAM (Fig. 2A) and dAM (Fig. 2B), with a limited number of small round epithelial cells located among them. After air-lifting (Fig. 2C, D), less spindle-shaped cells were visible and the epithelial cells were spread over iAM and dAM with a cobblestone appearance.
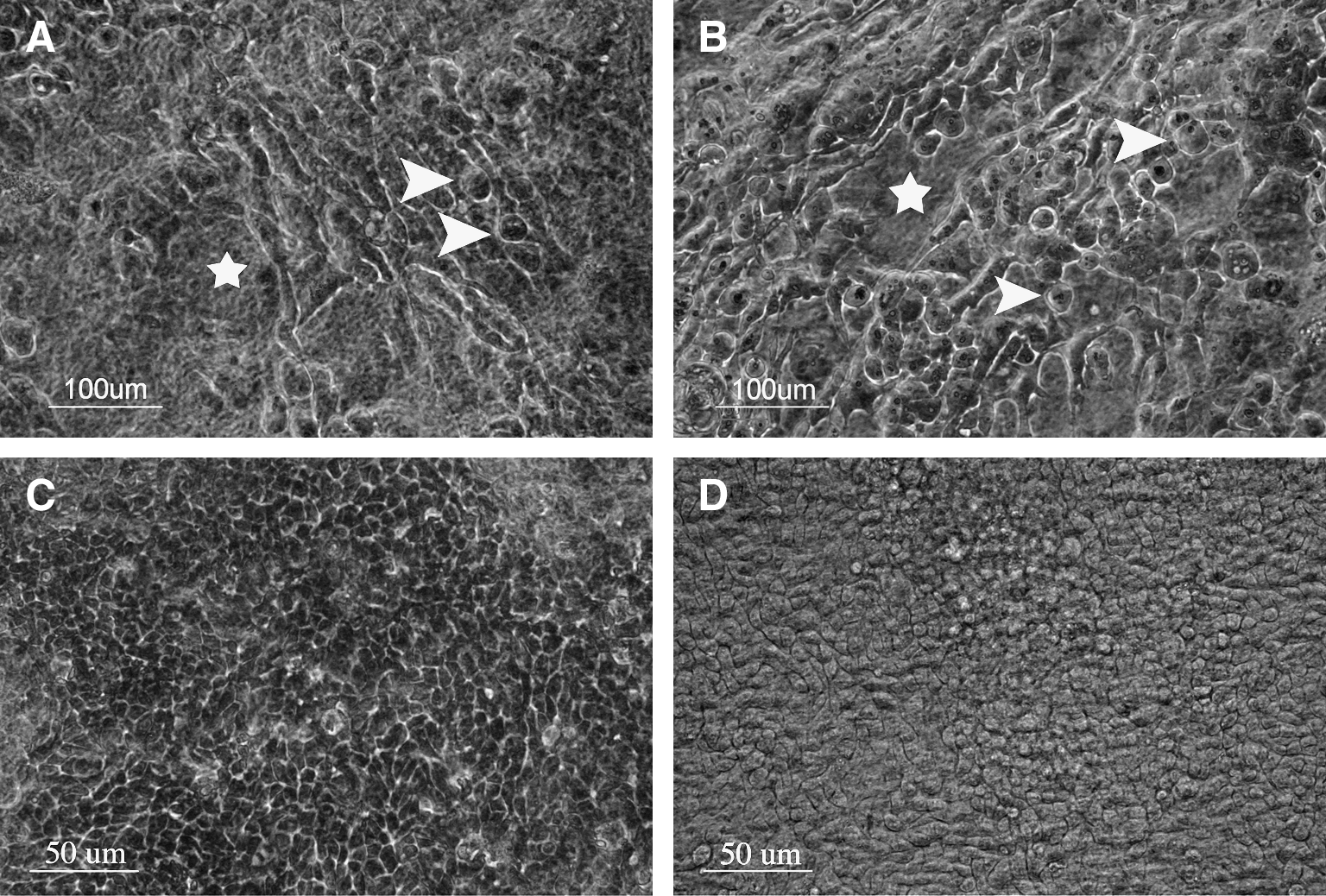
Epithelial cells grown on iAM (left column) and dAM (right column) before air-lifting (
Expression of differentiation markers in limbal epithelial cells expanded on AM
To assess relative expression of differentiated corneal epithelial cells, K3/12 and or K14 levels were evaluated by real-time PCR, Western blotting (Fig. 3A, B respectively), and immunostaining (Fig. 4).
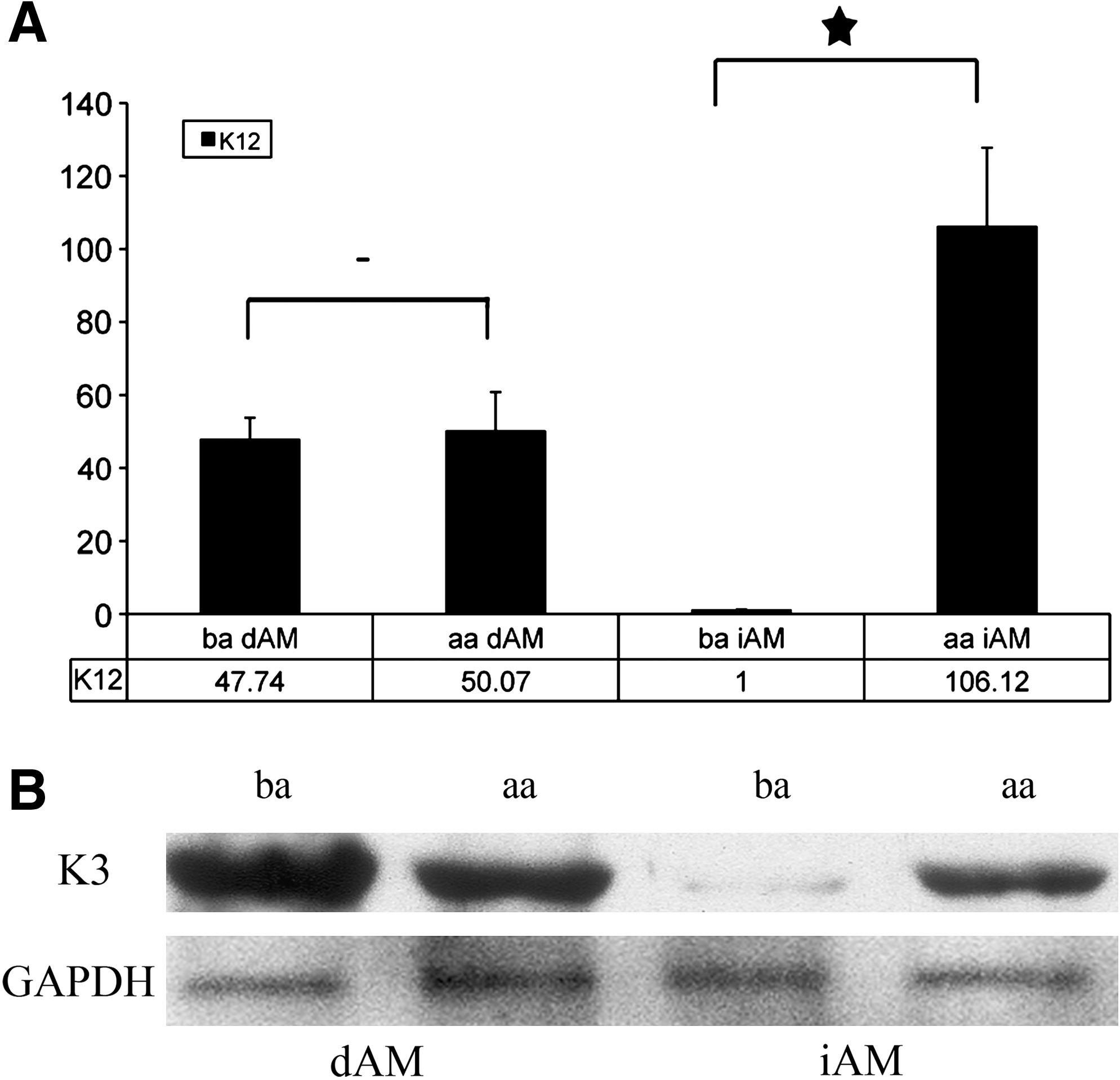
K12 mRNA and K3 protein expression in cultured epithelial cells on iAM and dAM, before air-lifting (ba; 14 days) and after air-lifting (aa; 21 days). Relative K12 mRNA levels did not change in cells cultured on dAM, but a significant increase in K12 mRNA was observed in cells cultured on iAM after air-lifting (−not significant; *p < 0.01) (
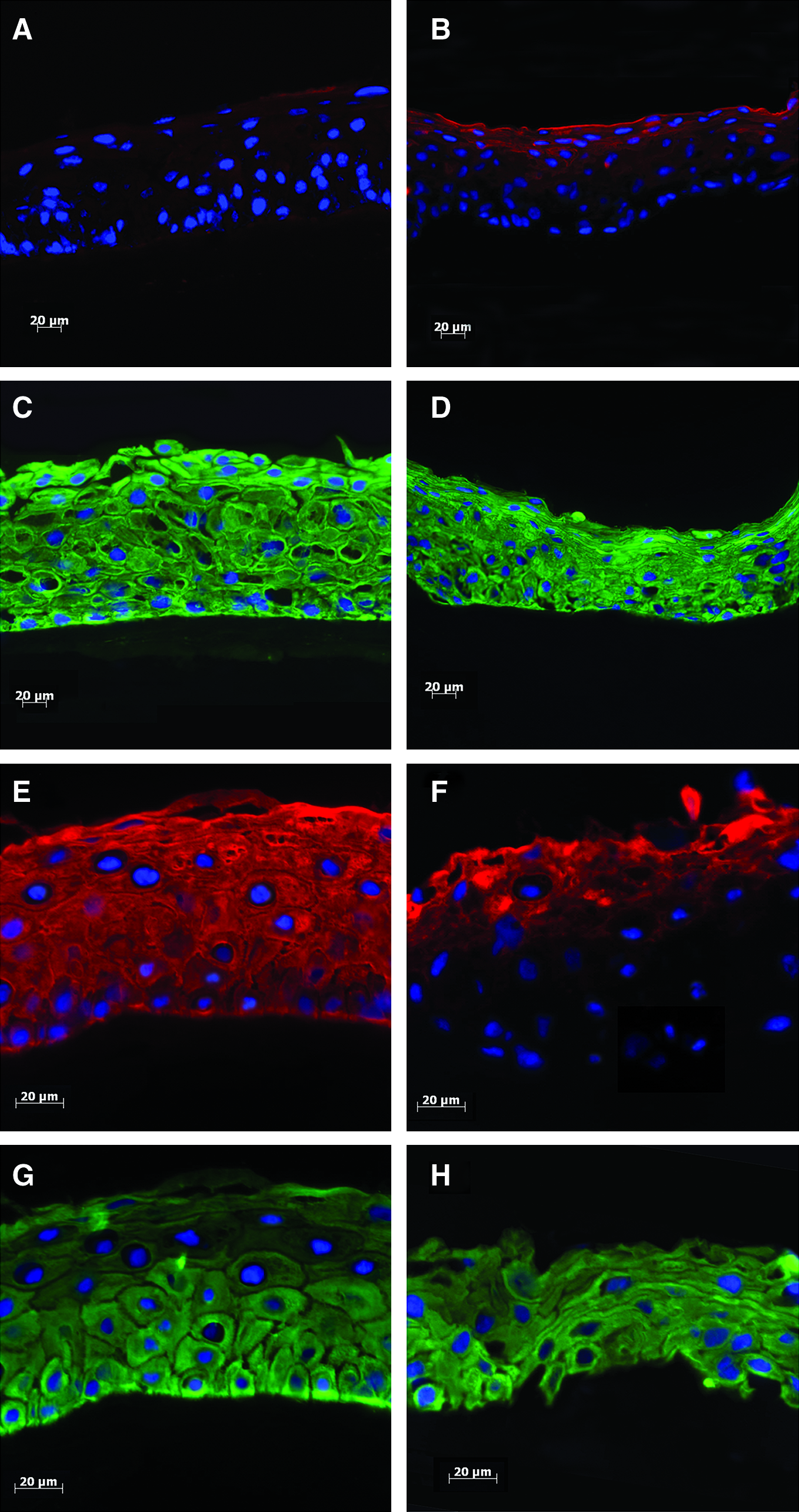
K3 (red) and K14 (green) expression in epithelial cells cultured on iAM and dAM before air-lifting at 14 days and after air-lifting at 21 days. Before air-lifting, a high proportion of the bovine limbal epithelial cells grown on iAM were K3 negative (
Using the housekeeping gene GAPDH and SDHA as internal controls, the real-time PCR results demonstrated that the K12 mRNA expression level in limbal epithelial cells cultured on iAM (Fig. 3A, ba iAM) increased significantly after air-lifting (Fig. 3A, aa iAM). By contrast, K12 mRNA levels in epithelial cells cultured on dAM are not significantly different (Fig. 3A, ba and aa AM). K3 and K12 are an associated pair of cytoskeletal filaments; therefore, the levels of K12 detected by real-time PCR are comparable with levels of K3 detected by Western blotting as seen in Figure 3.
K3/12 protein levels assessed by Western blotting almost mirrored the pattern of gene expression observed by real-time PCR. K3/12 protein (K3 associates with K12 in bovine cornea epithelium) 24 expression levels increased followed air-lifting in cells cultured on iAM, but K3/12 expression level before and after air-lifting remained similar in cells cultured on dAM (Fig. 3B).
Immunohistochemistry
K3 and K14 (markers for differentiated and undifferentiated corneal epithelial cells, respectively) 24 expression in epithelial cells cultured on iAM and dAM was observed by immunohistochemistry (Fig. 4). K3 and K14 are not expressed in native AM epithelial cells. 1
After a further 7 days in culture (air-lifted), there was no increase in the number of cell layers compared with 14 days cultured cells on AM (Fig. 4), but K3-positive cell layers had increased markedly in cells grown on iAM (Fig. 4E) compared with cultured cells without air-lifting (Fig. 4A). K3 expression did not change before and after air-lifting in cells grown on dAM, supporting real-time PCR and Western blotting results showing no difference in mRNA and protein expression of K3 in cells grown under the same conditions (Fig. 3). K3/12 results are summarized in Table 1. Keratin 14 expression remained at a similar level in corneal epithelial cells cultured on iAM and dAM after 1-week air-lifting (Fig. 4G, H).
Protein expression and mRNA expression of K3/12 are summarized as levels quantified or observed within limbal epithelial cells grown on iAM and dAM either before or after air-lifting.
iAM, intact amniotic membrane; dAM, denuded AM; Q-PCR, quantitative polymerase chain reaction; IHC, immunohistochemistry.
Integrin α6 expression was observed on both iAM and dAM before (1 week) and after (3 weeks) air-lifting (Fig. 5A, B, E, F). In contrast, laminin expression was initially absent on dAM, before air-lifting (Fig. 5G), but reappeared after air-lifting (Fig. 5H), with a similar expression level to iAM (Fig. 5C, D).

Integrin α6 (red) and laminin (green) expression in cultures of bovine corneal epithelial cells on iAM and dAM. Epithelial cells before air-lifting cultured for 7 days (
Discussion
AM as a culture substrate has previously been shown to provide excellent support for ex vivo limbal stem cell expansion.9,42 Further, by retaining the AM epithelium before limbal stem cell expansion (iAM), the expanding limbal cells will remain in an undifferentiated state. 31 Conversely, by culturing limbal epithelial cells under air-lifting conditions, differentiation and cell–cell attachment are promoted, 37 creating a superficial epithelial cell barrier. 43
In this study we have demonstrated that the manner in which the AM is prepared, before ex vivo limbal epithelial cell expansion, can strongly influence cell differentiation and that this can be affected by air-lifting. Our results are consistent with the previous findings that show, after 2 weeks culturing before air-lifting, the majority of limbal epithelial cells cultured on iAM remain in an undifferentiated state9,31 (i.e., K14 positive, and K3/12 limited expression) and that after 1-week air-lifting there is a change in limbal cell morphology (from spindle to tightly packed cells) and an increase in the level of differentiation, demonstrated by increased expression of K3/12.33–36 We have also observed K3 expression when bovine cornel epithelial cells are cultured on iAM for three or more weeks without air-lifting; however, K3-positive cells only occurred in superbasal layers (data not shown). Importantly, however, our results build significantly on these previous studies as we further demonstrate that significantly increased differentiation, after air-lifting, occurs only when the cells are cultured on iAM. A proportion of the limbal epithelial cells, when cultivated on dAM, are differentiated even before air-lifting, suggesting that air-lifting has a limited effect on cell differentiation when dAM is used (as the amount of K3/K12 remained the same). Specifically, using multiple lines of evidence, this article reports that after air-lifting the proportion of K3/K12-positive (differentiated) limbal epithelial cells cultured on iAM increases significantly, whereas it remains unchanged on dAM (Table 1).
We have clearly demonstrated, by performing an ultrastructural investigation, that retention of the native AM epithelium forms a physical barrier covering the underlying AM stroma. This study also shows that conventional mechanical removal of native AM epithelium reveals a mesh-work of stromal collagen fibers, together with others, evidence that this removal process causes basement membrane damage. 38 Mechanical removal of the AM's native epithelium has previously been shown to disturb the composition and structure of its basement membrane, 38 and this denuding process is believed to influence subsequent corneal epithelial cell growth.
Limbal epithelial cells cultured on dAM resemble corneal epithelia produced in response to a wound-healing stimulus. 31 The mechanism for this is not well understood, but in the normal eye, after corneal wounding, the basement membrane is destroyed in a controlled manner exposing the migrating epithelial cells to the underlying stroma. 44 Damage to the basement membrane facilitates direct epithelial–stromal interaction resulting in proliferation and differentiation of the migrating cells.22,45 These processes may well occur in vitro using dAM, as in the current study, in which the basement membrane of AM is similarly damaged exposing the cultured corneal epithelial cells to the stroma. Therefore, dAM is a suitable substrate to enhance cell–cell and cell–surface attachment, 43 which has subsequently been shown to influence cell differentiation. 46
To investigate further the potential for dAM to influence differentiation, we undertook a study of integrin α6 and AM basement membrane protein laminin expression between expanded limbal epithelial cells before and after air-lifting on iAM and dAM. Our results demonstrate that dAM did not noticeably affect (after 1–4 weeks in culture) integrin α6 expression, but AM basement proteins were absent before air-lifting, suggesting that cultured cells directly attached to the underlying collagen. Further, transmission electron microscopy demonstrated that corneal epithelial cells cultured on dAM showed considerably more desmosomal junctions and attachment complexes (hemidesmosomes) than the cells cultured on iAM. 11 Therefore, these observations may help to explain the measured difference in differentiation (K3/K12 expression) within limbal epithelial cells cultured on iAM and dAM. Limbal epithelial cells (which contain a high proportion of relatively undifferentiated cells) cultured on dAM come into direct contact with stromal collagens, which then associate with integrin α6 forming cell–matrix attachments. The formation of these cell–matrix protein complexes leads to direct epithelial–stromal interaction, which then triggers cell differentiation.43,47,48 This may occur almost immediately after cells are seeded onto the membrane, as in the present study limbal epithelial cells were observed to differentiate on dAM before air-lifting. We have shown a similar response when corneal epithelial cells are cultured directly on compressed collagen gel. 39 Therefore, limbal epithelial cells seeded onto iAM may not be able to differentiate before air-lifting, because the resident basement membrane together with native AM epithelium acts as a barrier preventing exposure of integrin α6 to the underlying stroma.
Conclusions
We believe that the findings reported here have significant and practical implications in terms of improving the surgical technique of limbal stem cell transplantation. Clear evidence has been produced that shows that to prevent limbal epithelial cells turning toward a differentiated state, at the early stages of cell culture, these cells should be cultivated on iAM and should not be air-lifted. The benefits, however, of using dAM cannot be dismissed as it can produce epithelial constructs with improved barrier function11,43 while still maintaining a population of undifferentiated cells, and this more mature construct may play a significant part in the successful use of dAM in limbal stem cell transplantations.14,16,49,50 Together, these findings could serve as a warning to the surgeon highlighting the fact that the effect of AM on transplantable cells is still not fully understood. Further, the use of limbal stem cells expanded upon either iAM after air-lifting or dAM before air-lifting could form the basis of a model system to investigate limbal stem cell growth and/or an oculotoxicity test to measure the toxic effect of drugs or chemicals on the surface of the eye.
Footnotes
Acknowledgments
The authors thank Cattleya Setio Leksmono for support in this study. This study was financially supported by the Biotechnology and Biological Sciences Research Council of United Kingdom (Grant Ref: BB/F019742/1), Cardiff University (School of Optometry and Vision Science), and University of Reading (RETF).
Disclosure Statement
No competing financial interests exist.