
Abstract
Myoblast-based therapy can improve cardiac function after infarction and is conventionally performed by direct injection. A scaffold-based transfer could overcome injection-associated problems. In upgrading this approach we transplanted skeletal myoblasts (SkM) overexpressing the prosurvival gene Akt1. SkM were transfected with pcDNA3-huda-Akt1 and seeded on polyurethane scaffolds. These scaffolds were transplanted in rats 2 weeks after myocardial infarction. Hemodynamics were analyzed before therapy and 6 weeks later. Infarction size and capillary density were performed thereafter. Additional groups received injections of Akt1-transfected or untransfected myoblasts, scaffolds seeded with untransfected myoblasts, or sham operation. Deterioration of global systolic left ventricular function could be inhibited by all therapeutic approaches. In addition, transplantation of Akt1-transfected cells, either scaffold-based or injected, was superior with regard to systolic properties of the left ventricular wall. This effect was accompanied by smaller infarction sizes and angiogenesis. Scaffolds with untransfected myoblasts yielded also smaller infarctions than injections of untransfected myoblasts. Both Akt groups profited with regard to dP/dt(min). In contrast, other diastolic parameters pointed at impaired relaxation and stiffer myocardium especially in the Akt1-scaffold group. In conclusion, SkM overexpressing Akt1 can maintain myocardial function after infarction, reduce infarction size, and induce neovascularization. Scaffold-based cell transfer does not augment this reverse remodeling capacity.
Introduction
A major drawback for studies using direct injection of stem cells is significant cell loss during the actual injection due to the physical trauma to the cells and the myocardial infarction zone's unfavorable target environment. We have previously suggested a scaffold-based transfer of the cells using SkM attached to long-term degradable polyurethane. The cells remained undifferentiated inside the scaffold, but the functional benefit was the same as with direct injection of the myoblasts, which gave us another indirect indication of the paracrine mechanisms involved. At the very least, we proved that the myocardial infarction is exposed to a large number of living myoblasts via this scaffold-based transfer.7,8
Genetic modulation of stem cells has been suggested for therapeutic purposes on one hand, and to study the effect of certain factors on the other. Akt1 has been used for transfection of MSCs to preserve their viability after the injection process. 9
Akt1 is an intracellular signaling molecule and one of three isoforms of the serine/threonine kinase Akt (or protein kinase B). It mediates its action by phosphorylating an array of target genes. Akt regulates survival, migration, and endothelial nitric oxide synthetase activity of endothelial cells 10 and is hereby involved in angiogenesis. 11 In addition, it inhibits apoptosis by phosphorylating several proapoptotic molecules (like BAD and caspase 9) and by activating transcription of antiapoptotic factors. 12 In myocardium, Akt is involved in cardiomyocyte survival, glucose metabolism, 13 and hypertrophy, both adaptive and compensational. 14 In a rat model of myocardial infarction, cells transfected with Akt1 normalized metabolism 15 and cardiac function. 9 This effect was also present when only the cell medium of the Akt1-MSC was injected. Therefore, other soluble factors that support cardiac function may be secreted by Akt1-modulated cells. 16
We have previously reported a strategy to combine the scaffold-based application of cells with genetically modified cells. 17 This study showed that scaffolds seeded with cells that secrete large amounts of soluble factors can act as a kind of cytokine factories. In the current study, we have tested the functional effect of this approach on the heart after myocardial infarction in the rat model.
Materials and Methods
Isolation and cell culture of primary SkM
SkM were isolated from male newborn Lewis rats as reported previously. 6 In brief, limb muscles were minced and enzymatically digested with 1.5 mg/mL collagenase IA (Sigma Chemical Company) and 0.25% trypsin/ethylenediaminetetraacetic acid (Gibco). Following passages through a 100-μm and a 40-μm filter (Falcon, BD), cells were expanded in the culture medium consisting of 57% minimum essential medium (MEM) (Gibco), 27% M199 (Gibco), 15% fetal bovine serum (HyClone), 1% penicillin/streptomycin (Gibco), and 5 ng/mL beta-fibroblast growth factor (Sigma). After 48 h, cells were harvested by trypsinization, counted, and frozen in fetal bovine serum containing 10% dimethyl sulfoxide (Sigma) until they were needed for scaffold preparation. We usually obtained an average of 1.5 ± 0.7 × 107 cells per newborn Lewis rat. Anti-desmin immunofluorescent staining (Sigma, clone DE-U-10) was performed to calculate the percentage of SkM in the preparation as described previously. 18 Culture purity achieved 46.8% ± 12.1% of desmin-positive myoblasts.
Preparation of polyurethane scaffolds and cell seeding
The polyurethane-urea (PU) matrix used in this study was obtained from Artimplant (Artelon®). Artelon is a soft, elastic material that degrades by hydrolysis over ∼5 years. The manufacture of Artelon membranes has been described. 19 For our purpose, the biomaterial was processed into a highly porous scaffold (90% ± 3% volume porosity) with interconnected pores measuring 30–300 μm.
For scaffold seeding, Artelon membranes were cut to a scaffold size of 1 × 7.5 × 7.5 mm and precoated with 10 μg/mL laminin (Invitrogen) for 45 min at room temperature. SkM were thawed, diluted, centrifuged, and counted. Preliminary viability tests showed that cells were unaffected by the freeze–thaw process (data not shown). Thirty microliters of cell suspension containing 5.0 × 106 cells was applied to the scaffold. This amount of cell suspension disperses into the matrix and thereby distributes the cells to the polyurethane as shown in detail previously. 8 The seeded scaffolds were incubated for 30 min at room temperature to allow cell attachment. Afterward, the culture medium was added and seeded PU scaffolds were incubated further at 37°C (5% CO2) for 48 h. This seeding protocol was identified as the optimal way to facilitate cell attachment in our preceding study. We found that almost 50% of cells attach. 8 After incubation, the quantification of attached cells yielded 3.2 ± 0.6 × 106 cells in the scaffolds. For control, unseeded scaffolds were prepared in the same way using 30 μL of the culture medium instead of cell suspension.
Transfection of primary SkM
Cell transfection was performed as described previously. 17 In brief, the plasmid pcDNA3-huda-Akt (expressing a myristinylated dominant-active form of Akt1) was delivered into SkM using the liposome-based transfection reagent Metafectene™ Pro (Biontex). According to the manufacturer's protocol, we incubated 5 μg plasmid DNA (diluted in 100 μL MEM) with 12 μL Metafectene Pro solution (diluted in 100 μL MEM) for 25 min at room temperature. After attachment of the cells, the culture medium (2 mL) of the cell-seeded PU scaffolds was renewed and transfection mix was added. After 24 h, the culture medium was changed again and the scaffolds were further incubated at 37°C (5% CO2). The plasmid was a generous gift from Robert Freeman (University of Rochester). Effective transfection of the cells was confirmed by real-time (RT)-polymerase chain reaction and Western blot as reported in our previous study. 17
Animal model
The animals received human care in compliance with the Principles of Laboratory Animal Care formulated by the National Society for Medical Research and the Guide for the Care and Use of Laboratory Animals prepared by the National Institute of Health (NIH Publication No. 86-23, revised 1985). All procedures were performed in accordance with the German Law on the Protection of Animals after obtaining permission from the Regional Administrative Authority (Freiburg, Germany, G-05/56).
The animal procedure has been described before. 7 In brief, female Lewis rats were anesthetized with isoflurane (5% of oxygen for induction and 2.5% of oxygen for maintenance) and additive buprenorphine (10 μg/kg subcutaneously), placed on a warming pad (37°C), and tracheally ventilated at 80 cycles/min (14-Gauge IV cannula, Abbocath®; Abbott; Small Animal Ventilator 683; Harvard Apparatus Inc.). A myocardial infarct (MI) was then induced through a left lateral thoracotomy by ligating the left anterior descending coronary artery. Two weeks later, a sternotomy allowed access to the infarction area for baseline catheterization and intervention. The animals were randomized to undergo either a sham operation (group sham, sternotomy, and adhesiolysis only, n = 10), the implantation of a scaffold seeded with transfected myoblasts (PU-Akt1-SkM, n = 9) or untreated myoblasts (PU-SkM, n = 9), or direct intramyocardial injection of transfected (Inj-Akt1-SkM, n = 10) or untreated (Inj-SkM, n = 9) myoblasts. In the PU-Akt1-SkM and PU-SkM groups, the scaffolds were placed on the epicardial surface of the visually identified infarction area and surgically fixated by four single epicardial sutures at the corners of the quadratic matrix (7/0 polypropylene). In the Inj-Akt1-SkM and Inj-SkM groups, 5.0 × 106 cells in 150 μL medium (containing 15% fetal bovine serum) were distributed within the scar via four to six intramyocardial injection sites. Six weeks after the intervention, the rats were anesthetized for left ventricular (LV) catheterization before the hearts were removed for histological analysis.
Hemodynamic assessment
Before therapeutic intervention and 6 weeks thereafter, LV function was assessed by online LV pressure–volume signals obtained by a 1.9 French pressure–volume catheter (FT212; Scisense Inc.). After sternotomy, a purse-string suture was placed in the apical region of the left ventricle, where the catheter was inserted and placed along the LV long axis using an IV cannula (Abbocath®) as a sheath. The catheter was connected to a Sigma-SA signal processor (CD Leycom) for display and acquisition of pressure–volume loops. We registered 3 cycles of 20 beats under respiratory arrest to measure the LV function, quantified by heart rate, LV end-diastolic and end-systolic pressure, the maximum rate of LV pressure increase [dP/dt(max)] and decrease [dP/dt(min)], LV ejection fraction (EF), and pressure half time (PHT).
Further, the isovolumic relaxation time constant (Tau) was determined with phase-plot analysis. In addition, we determined the isovolumic contraction (PISO) and, in single beat estimation, the end-systolic elastance (EES) and the diastolic chamber stiffness constant (KED).
Data were transferred to the data analysis software package CIRCLAB, which was a generous gift from Dr. Paul Steendijk (Leiden, University Medical Center, The Netherlands).
Table 1 lists the main hemodynamic parameters measured pre- and postinterventionally, that is, before therapeutic procedure and after 6 weeks.
Infarction size and angiogenesis
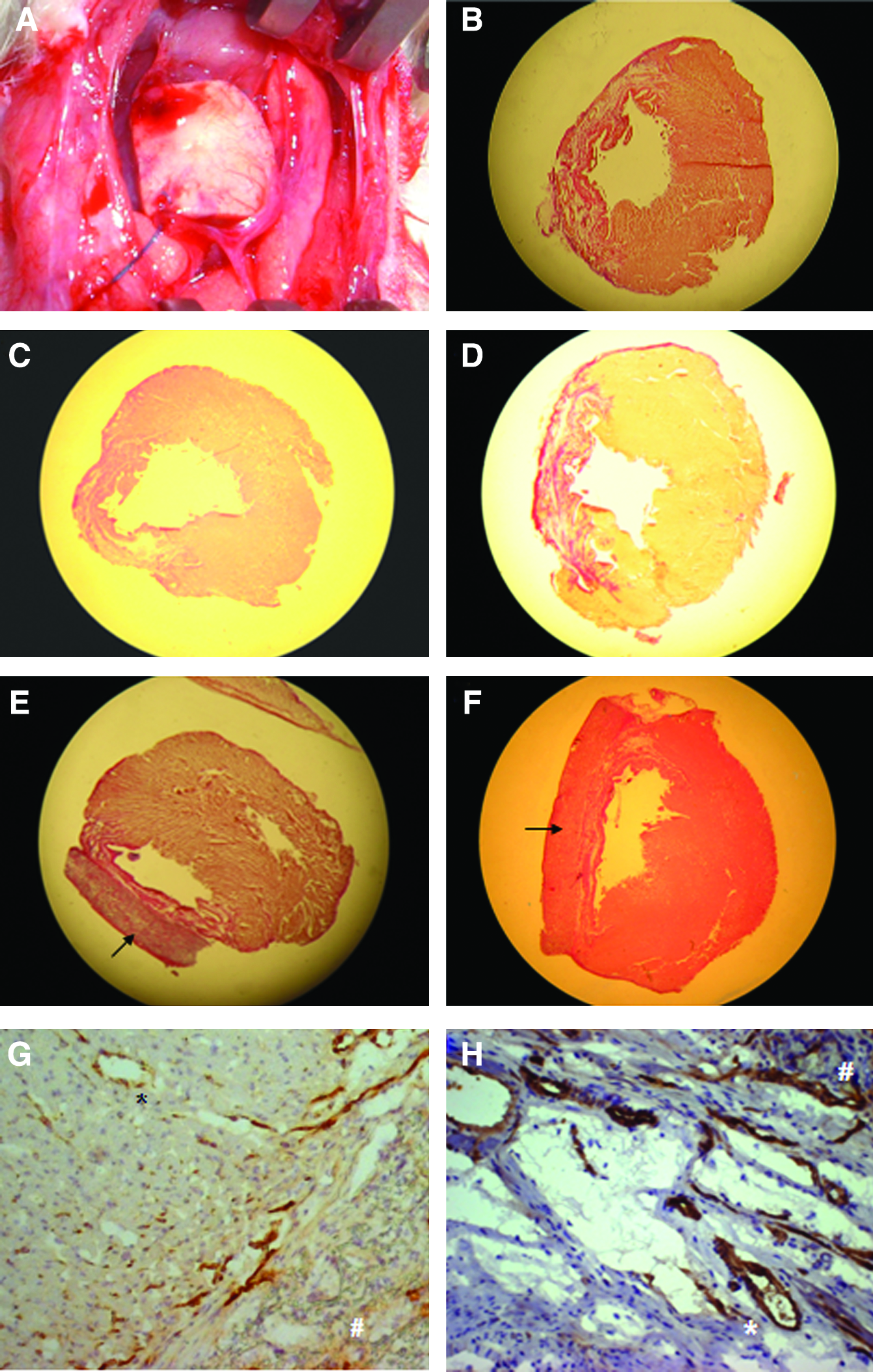
Hearts were explanted and sliced into three parts of equal thickness orthogonally to the heart axis. The pieces were frozen in Jung Tissue Freezing Medium (Leica Microsystems) and stored at −20°C. Cryosections of 5 μm were prepared and fixed with ice-cold acetone. An Elastika van Gieson staining kit (Merck) was used to observe the infarcted myocardium in each of the three slices. Staining was performed according to the manufacturer's instructions. The sections were then photographed with a digital camera. We calculated the living and infarcted areas, and then integrated the values of all sections to estimate the infarction size of the three slices using an image-analysis program (ImageTool2.0; Microsoft Corporation).
For anti–von Willebrand factor staining, endogenous peroxidase activity was quenched by peroxidase-blocking reagent (Dako). The sections were then blocked with 10% goat serum and subsequently incubated with rabbit anti-human von Willebrand factor antibody (Dako, 1:250) for 1 h at room temperature. All stainings were developed with horseradish peroxidase-goat anti-rabbit secondary antibody (Dako) followed by DAB chromogen (Dako) and counterstained with hematoxylin (Merck). We counted the positively stained capillaries in 10 visual fields (magnification 400×) for each heart in the infarction zone, the remote myocardium, and the infarction border zone. Capillaries were identified by a one-layered vessel wall, a present or collapsed lumen, and a diameter of up to about 10 μm. Arterioles, arteries, and veins were excluded.
Statistical analysis
Data are expressed as mean ± standard deviation. The statistical significance of differences between pre- and postinterventional data within one group was calculated using Student's t-test. One-way analysis of variance (ANOVA) was used to identify differences among the groups in capillary density and infarction size at the end of the investigation. Two-way ANOVA for repeated measurements was used to compare hemodynamic pre- and postinterventional variables among groups. Both ANOVA tests were followed by Holm-Sidak as a post-hoc test. Differences were considered significant if p < 0.05.
Results
Morbidity and mortality
Of a total of 93 operated animals, 47 could be included in this study (51%). Twenty-eight rats (30%) died during the study—mainly of ventricular arrhythmia after the infarction (eight animals) or during the first reoperation with hemodynamic assessment and treatment (eight animals). Another 12 rats died between the treatment and the examination without revealing significant group-specific differences (four sham, two Inj-SkM, three PU-SkM, one Inj-Akt1-SkM, and two PU-Akt1-SkM). We excluded further 18 animals (19%) on the basis of the hemodynamic assessment (13 rats) before randomization because of severely impaired LV function or due to technical problems with the hemodynamic measurement setup (5 rats).
Hemodynamic assessment
At randomization, none of the hemodynamic values differed among all groups. Heart rates were not affected by the treatment and were slightly higher at the study end, without revealing group differences (data not shown). The changes in percent from baseline to study end of the main hemodynamic parameters are highlighted in Figure 1a (systolic LV function) and Figure 1b (diastolic function).

Systolic
In the sham group, we noticed a drop of systolic LV function with significantly reduced EF (from 25% ± 14% to 11% ± 8%, p = 0.032) and dP/dt(max) (from 2180 ± 693 at baseline to 1709 ± 337 mmHg/s at the study end, p = 0.048).
In the groups that received SkM without transfection of Akt1, we observed preserved systolic LV function with unchanged or insignificantly improved hemodynamic variables from baseline to the study end. In the intergroup comparison, dP/dt(max) was higher in groups PU-SkM and Inj-SkM than in the sham group (p = 0.034 and p = 0.037, respectively) at the study end. In addition, dP/dt(min) improved in group PU-SkM but only compared to sham (p = 0.024).
In group Inj-Akt1-SkM, we observed a rise from baseline to the end of the study in the following systolic and diastolic parameters: dP/dt(max) (from 2043 ± 1394 to 3556 ± 1817 mmHg/s, p = 0.009), PISO (from 82.4 ± 43.4 to 142.2 ± 70.1 mmHg, p = 0.004), Ees (from 263 ± 143 to 561 ± 448 mmHg/μL, p = 0.029), dP/dt(min) (from −1826 ± 1558 to −3821 ±2340 mmHg/s, p = 0.047), and Ked (from 3.3 ± 2.0 to 6.2 ± 3.5, p = 0.023). The values for dP/dt(max) and dP/dt(min) at the study end were significantly higher in the Inj-Akt1-SkM group than in the sham group (p = 0.001 and p = 0.001, respectively).
Various significant changes occurred in the PU-Akt1-SkM group. The following systolic and diastolic parameters were significantly higher at the study end when compared to the baseline hemodynamics dP/dt(max) (from 2224 ± 891 to 3203 ± 1196 mmHg/s, p = 0.039), PISO (from 89.5 ± 43.4 to 147.6 ± 34.2 mmHg, p < 0.001), Ees (from 310 ± 141 to 704 ±410 mmHg/μL, p = 0.036), dP/dt(min) (from −2213 ± 1006 to −3223 ± 1188 mmHg/s, p = 0.008), Tau (from 20.2 ± 2.6 to 26.4 ± 5.9 ms, p = 0.006), PHT (from 8.3 ± 1.9 to 13.8 ± 2.7 ms, p = 0.002), and Ked (from 3.7 ± 1.4 to 7.6 ± 1.0, p = 0.031). In the inter-group comparison, the dP/dt(max) and dP/dt(min) values at the study end were higher than those of the sham group (p = 0.006 and p = 0.014, respectively), and the PHT values were higher than those of the sham, PU-SkM, and Inj-Akt1-SkM groups (p = 0.016, p = 0.018, and p = 0.025, respectively).
Infarction size and capillary density
As highlighted in Figure 2, we measured significantly larger myocardial infarction areas in the sham group than in the PU-SkM (p = 0.027) and Inj-Akt1-SkM groups (p =0.010); they were highly significantly larger than in the PU-Akt1-SkM group (p = 0.004) (Fig. 4B–F).
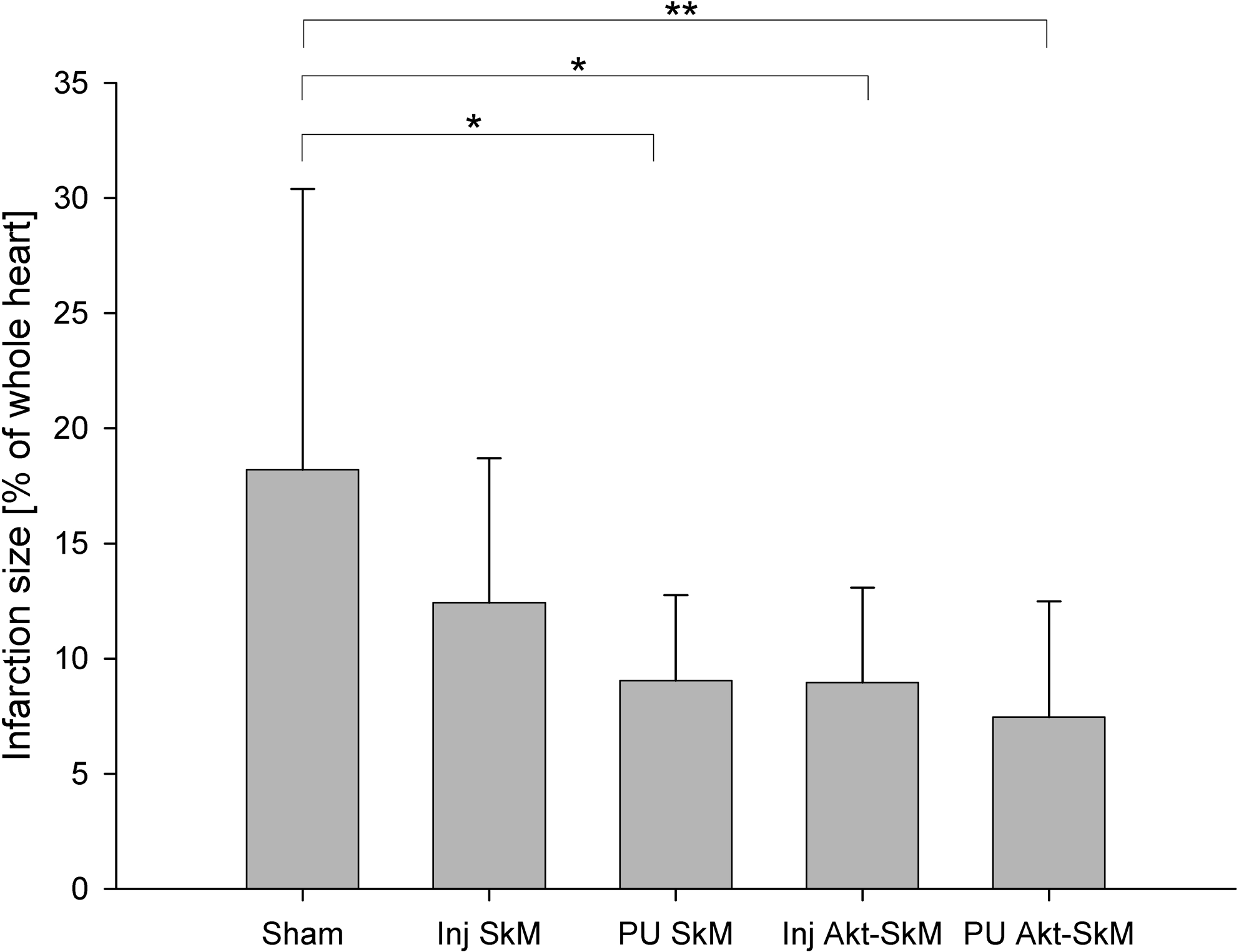
Infarction size in percent of the whole heart. Three parts of the excised hearts were stained with Elastika van Gieson staining to observe the infarcted myocardium. From the photographed heart slices, we calculated the living and infarcted areas using an image analysis program. Mean values ± standard deviation are given (*p < 0.05 and **p < 0.01 between groups).
We observed multiple capillaries entering the seeded patches at the study end (Fig. 4A). However, the density of spreading vessels did not differ between the PU groups.
Capillary density was measured in three different compartments of the heart (Table 2). Capillary density in the viable myocardium was much higher in all groups compared to the infarction zone. However, we observed no intergroup differences in the capillary density in the infarction zone and in the remote myocardium (Table 1). In the border zone, we observed significantly higher capillary density in the Inj-Akt1-SkM and PU-Akt1-SkM groups than in the sham and Inj-SkM groups (Fig. 3). Figure 4 illustrates sections of lower capillary density in the infarction border zone of a PU-SkM treated animal (panel G) compared to PU-Akt1-SkM (panel H).
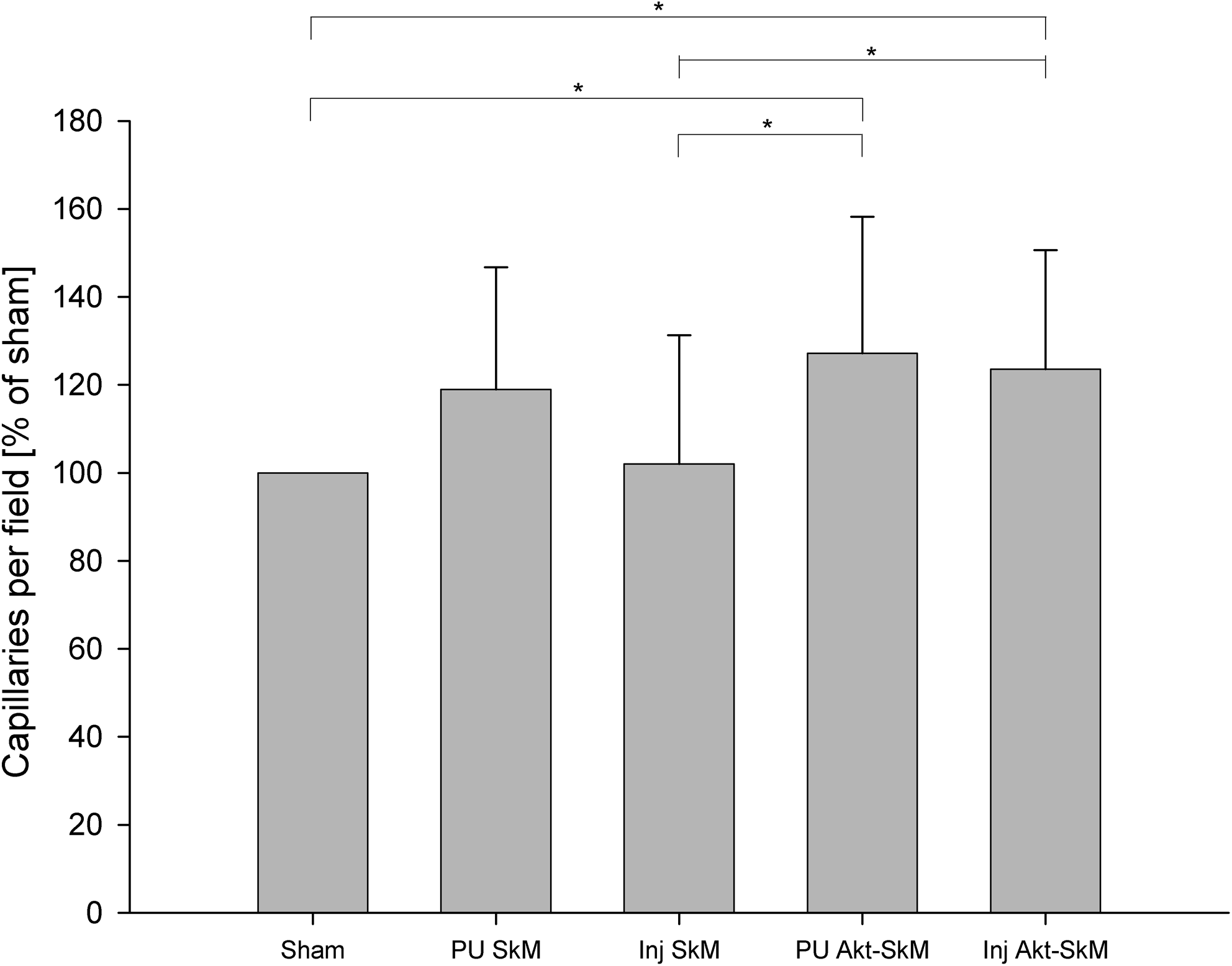
Capillary density in the infarction border zone. Capillaries were stained with anti–von Willebrand factor and counted in 10 visual fields (400 × magnification). Values are given in percent of sham group, which was normalized to 100% (*p < 0.05 between groups).
Capillaries were identified by anti–von Willebrand factor staining and counted in the infarction zone, the remote myocardium, and the infarction border zone in 10 visual fields (magnification 400 × ) for each heart.
SkM, skeletal myoblasts; n.s., not significant; PU, polyurethane.
Discussion
The present study examines the effect of Akt-transfected SkM on infarcted myocardium in rats. Two weeks after ligation of the left anterior descending coronary artery, hemodynamics were assessed at baseline and Akt-transfected myoblasts were transplanted by injection or attachment of a cell-seeded polyurethane scaffold. Untransfected myoblasts, for injection or seeded on scaffolds, were used for control purposes. Six weeks later, the therapy groups were compared to sham-operated animals with regard to hemodynamics, infarction size, and capillary density.
The deterioration of global systolic LV function [LVEF and dP/dt(max)] that follows myocardial infarction could be inhibited in comparison to the sham animals by transplanting SkM, both scaffold-based or after intramyocardial injection and Akt1-transfected or not. In addition, transplantation of Akt1-transfected cells, either scaffold-based or injected, was superior compared to untransfected cells with regard to systolic properties of the LV wall (Ees, PISO). This effect was accompanied by smaller infarction size and angiogenesis. Relating to the extent of infarction, scaffolds with untransfected myoblasts yielded also better results than injections of untransfected myoblasts. Both Akt groups profited with regard to dP/dt(min). In contrast, other diastolic parameters pointed at impaired relaxation (PHT, Tau) and higher stiffness of the myocardium (Ked) especially in the PU-Akt1 group, while the other groups underwent no change.
Effects of transplanting SkM
In numerous preclinical studies, SkM were shown to be capable to regenerate myocardial function. That effect was observed in small3,7 and large animals. 20 The functional endpoints of most preclinical studies were usually assessed using echo, 3 LV catheter, 7 or magnet-resonance-tomography (MRT). 21
Remarkable improvements in functional parameters have been reported, which sometimes reached values similar to those of normal hearts without myocardial infarction. Ott et al. 22 treated rats with multiple microinjections of SkM for myocardial infarction and achieved an EF of around 70%. In our previous study, 7 scaffold-based transfer of untransfected SkM stopped the functional decline after myocardial infarction, which was observed in the sham group. However, we noted no significant improvement compared to baseline values. In the experiments presented here, we failed again to achieve a clear improvement in LV function by untransfected SkM. In accordance, Menasché et al. observed almost no effect from SkM transplantation in failing hearts when compared to the control group in a randomized clinical phase II trial. 23
Transplantation of Akt1-transfected SkM
The Akt1-SkM groups rendered significantly higher values of some hemodynamic parameters than the sham group. MSCs transfected with Akt1 were used by other investigators. They showed that the functional effect of Akt1-MSC in small 9 and large animals 24 is superior to classic MSC transplantation. The gene transfer was initiated primarily to overcome frequent cell death after injection 9 since Akt1 should lead to more resistant cells for the application process. However, Dzau's group later discovered that functional results of Akt1-MSC can also be achieved by injecting the medium in which Akt1-MSC were cultured. 16 Hereby, this study proofed the actual paracrine effect of this approach.
We speculate that the same cause for functional benefit can be claimed for the effects observed in our study with Akt1-modified SkM. Previously, no difference in Akt activity between SkM and bone marrow cells was found by Shintani et al. when they compared the paracrine profiles. 25 To the best of our knowledge, ours is the first study that used Akt1-overexpressing SkM;, therefore, we can only infer the underlying mechanisms.
Angiogenesis in the infarction border zone
We observed that the hemodynamical superiority of the therapies with Akt1-modified SkM was accompanied by higher capillary density in the infarction bordering zone compared to the sham and myoblast injection groups.
We have previously described that Akt1-overexpressing myoblasts seeded on scaffolds have elevated vascular endothelial growth factor (VEGF) secretion levels, an effect that we used to optimize the transfection protocol. 17 A close inter-relationship between VEGF signaling and Akt1-activation has been proposed to play a critical role in angiogenesis.26,27 However, our results permit us only to describe the coincidental finding of increased capillary density and improved function in animals that received Akt1-modified cells and not to infer about the pathway to augmented angiogenesis.
The notion of smaller infarction zone requires similar consideration as angiogenesis. The positive effects of Akt on cell survival in general are well known. 28 Specifically, Dzau's group revealed that the preservation of metabolism in the surviving myocardium may be responsible for positive effects on the failing heart. 15 In correlation with our own finding, we presume that hibernating myocardium in the infarction bordering zone could be salvaged by transplanted Akt1-overexpressing cells. In this process, Akt-dependent VEGF upregulation could play a role. VEGF has been shown to be part of the genetic program that regulates hibernation and subsequent return of function. 29 Other effects on the extracellular matrix cannot be excluded.
Scaffold-based application of SkM
In a recent study, Chachques et al. employed bone marrow cells on a collagen scaffold and applied them on the myocardial infarction zone of patients. 30 They found an additive effect on diastolic function. In our experiments, the most pronounced decrease in infarction size was found in the group with Akt1-SkM-seeded scaffolds. However, the present approach leads to higher Tau and PHT as well as to higher Ked. These data point to impaired relaxation and higher myocardial stiffness, respectively. In contrast, there was no difference to the baseline when we used scaffolds with untransfected myoblasts. These observations could indicate that scaffolds seeded with Akt-transfected or untransfected myoblasts develop different physical properties when transplanted on infarcted myocardium. It has to be taken into consideration that this is an effect of Akt on the SkM themselves since Akt supports the differentiation into myotubes. 31
Study limitations
The main reason for the small differences among groups is the high standard deviation. The larger differences within the groups could be due to varying severity of the myocardial infarction, even though the MI procedure was always performed in the same manner. The means of data acquisition might also have caused our parameter to vary. Serial catheterization is very difficult in small animals. We found it particularly helpful to use the LV apex as access route for both catheterizations to allow for comparison of parameters at different time-points. We recommend this technique because both carotids remain untouched and penetration of the aortic valve is not necessary. However, some animals might have suffered from this apical insertion more than others, which may explain the variability to a certain extent.
Our study could be subject to criticism for the use of SkM, which are unlikely to play a major role in future cell therapy approaches after the disappointing Magic-II trial results. 23 We chose this approach mainly because we wanted to investigate the effect of Akt1-overexpressing cells seeded on a scaffold on myocardial infarction.
In our previous study, we had observed no beneficial effect on LV function by the scaffold alone. 7 The group that received an empty scaffold behaved functionally exactly like the sham control group. We used the same animal instrumentation in the current investigation and therefore refrained from including an additional control group with an empty scaffold.
In conclusion, SkM overexpressing Akt1 can restore cardiac function after myocardial infarction in some aspects. Scaffold-based cell transfer does not enhance this reverse remodeling capacity but may increase LV wall stiffness. The beneficial functional effects were accompanied by a reduced infarction size and higher capillary density in the animals that received Akt1-modified cells.
Footnotes
Acknowledgments
This work was supported by the Deutsche Forschungsgemeinschaft (SI 858/3-1) and Deutsche Stiftung für Herzforschung (F/10/08). We thank Heide Marniga for technical assistance and Youhua Liu, Kensuke Egashira, and Robert Freeman for their collaboration on this study. Dr. Paul Steendijk has provided data analysis software and was a great help in data interpretation. We would like to thank Carole Cuerten and Con Lehane for editorial assistance.
Disclosure Statement
No competing financial interests exist.