
Abstract
The use of calcium phosphate (CaP)-based carriers in bone engineering is a promising approach to induce in vivo bone formation. However, the exact mechanism of osteoinduction by CaP is not known. Here, by mimicking the in vivo Ca2+ and Pi-enriched environment in an in vitro model, we assessed the effects of these ions on human periosteum-derived cells. We observed a significant Ca2+ and Pi-induced cell proliferation, which was found to be through the modulation of cell cycle progression, in a dose- and time-dependent manner. In addition, Ca2+, Pi, and combined Ca-P upregulated osteogenic gene expression in a dose- and time-dependent manner. Encouragingly, both ions administered individually or in combination persistently and dose dependently upregulated bone morphogenetic protein-2 gene expression. This suggested a potential osteoinductive effect through an autonomous activation of the bone morphogenetic protein signaling pathway by released Ca2+ and Pi, which may serve as an autocrine/paracrine osteoinduction loop that drives the cellularized CaP constructs toward effective bone formation in vivo. In conclusion, through an in vitro biomimetic model, we have partially probed the roles of the released Ca2+ and Pi on the osteoinductivity of CaP-based biomaterials.
Introduction
Ripamonti et al. have previously suggested that the entrapment of a threshold amount of endogenous BMP molecules by CaP implants possibly renders the material osteoinductive through the de novo activation of the BMP signaling cascade.16,17 These same investigators further reported that a specific geometry, porosity, and concavity promoted osteoinduction through biomimetising the osteonic remodeling processes.18,19 This is in line with the findings from other research groups20–22 that reported the direct biological effects 23 and physicochemical properties 24 of the macro- and microporosity on cellular focal adhesions, 25 mechano-transduction,26,27 osteogenicity, 28 growth factor adsorption and release, 29 vascularization, and final maturation of the bone tissue into a functional organ. 30 The presence of well-defined macropores, interparticle spaces, and micropores is beneficial for the penetration of bodily fluids and protein entrapment and provides a specific surface area 31 that influences the release kinetics of calcium (Ca2+) and phosphate (Pi) ions in the local vicinity.32–35 The release of these ions is believed to be the determinant of CaP bioactivity, encouraging the formation of a biological apatite layer and triggering local stem cells into osteogenic differentiation.20,36 A similar hypothesis was deducted from a study showing osteoinduction on chemical and heat-treated titanium implants, whereby specific microstructures caused the formation of a biological apatite layer on the implant surface, and therefore suggested the importance of Ca2+ and Pi on osteogenesis. 37 In addition, studies on tendon, 38 artery, 39 and polymer 40 calcification-induced bone formation have provided strong evidence that a Ca2+ and Pi-enriched environment is a prerequisite for osteoinduction. Indeed, a recent study reported by our group showed that chemical removal of CaP granules from the commercially available CaP-collagen composite (i.e., Collagraft™) seeded with human periosteum-derived cells (hPDCs) abrogated ectopic bone formation. 41
In this study, we have developed an in vitro Ca2+ and Pi system that allows us to probe the roles of Ca2+ and Pi individually and in combination, within a broader physiologically relevant concentration range. We believe that this system could sufficiently mimic a local environment that becomes enriched with different [Ca2+] and [Pi] due to dissolution from the implanted biomaterial. More specifically, beside gaining more insight on the mechanism of CaP-driven osteoinductivity, we would also like to identify the optimal concentration for [Ca2+] and [Pi] or in combination to effectively drive a clinically relevant mesenchymal stem cell (MSC) population—in this case hPDCs, along the axis of enhanced cell proliferation, cell aggregation, osteogenic differentiation, mineralization, and eventually enabling translation of this important fundamental biological know-how to the engineering of an intelligent osteoinductive scaffold for an effective bone engineering strategy.
Materials and Methods
Preparation of Ca2+- and Pi-supplemented growth media
About 100 × Ca2+ or Pi stock solutions of 200, 400, 600, 800, and 1000 mM were prepared by dissolving appropriate amounts of calcium chloride (CaCl2 · 6H2O; Applichem) or di-sodium hydrogen phosphate dihydrate (Na2HPO4 · 2H2O; Merck) and sodium di-hydrogen phosphate dihydrate (NaH2PO4 · 2H2O; Merck) in a ratio of 4:1 in Hepes Buffer Solution (25 mM Hepes [Sigma] + 140 mM NaCl + 390 mL demineralized water) 42 and sterilized by a syringe filter (0.2 μm). The Ca2+, Pi, and combined Ca-P supplemented growth media were prepared by diluting the stock solution to 1% (v/v) in growth media (GM) (Dulbecco's modified Eagle's medium + GlutaMax™-1 [Gibco] + 10% fetal bovine serum [Gibco] + 1% antibiotics + 1% sodium pyruvate), yielding a final concentration of 2, 4, 6, 8, and 10 mM either individually or in combination. Fluctuations in pH of the GM were not observed following Ca2+ and Pi addition (data not shown). For nomenclature purpose, Ca2+ is denoted as “C” and Pi is denoted as “P,” followed by a number indicating the final concentration in mM.
Cell culture of hPDCs
hPDCs were isolated from periosteal biopsies of different donors as described previously. 41 This procedure was approved by the ethics committee for Human Medical Research (Katholieke Universiteit Leuven) and with patient informed consent. Upon confluence, hPDCs were trypsin released and replated at a seeding density as specified in the experiments. After plating, the cells were cultured for 2 days and synchronized for 24 h in the serum-free medium (Dulbecco's modified Eagle's medium + 0.01% bovine serum albumin; BD Biosciences). Freshly prepared Ca2+, Pi, or combined Ca-P supplemented growth media was then added to the cell cultures and incubated for a defined experimental time frame (as mentioned in the following paragraphs) before harvesting for analysis. Cells treated with the growth medium were used as negative controls (GM), growth medium containing 1% (v/v) Hepes Buffer Solution as vehicle controls (Vh), and osteogenic medium (OM) (growth medium supplemented with 100 nM dexamethasone, 50 μg/mL ascorbic acid, and 10 mM β-glycerophosphate) as positive controls. The culture medium was refreshed twice a week; old medium was collected for Ca2+ and Pi analysis by inductively coupled plasma atomic emission spectroscopy (Varian Inc.) (n = 4), which indicated sustainable levels of [Ca2+] and [Pi] between each media refreshment (data not shown).
hPDC growth assay
hPDCs were seeded at 4500 cells/cm2 in 24-well plates and treated for 1, 3, 7, 14, 21, and 28 days (n = 4). At each time point, the cell cultures were harvested and lysed in 0.05% Triton-X100 (in phosphate buffered saline [PBS]) and the DNA content was measured using Quant-iT™ dsDNA HS Assay Kit (Invitrogen) according to the manufacturer's instructions. For metabolic activity measurement, the growth medium containing 10% Alamar Blue solution (AlamarBlue™ kit; Invitrogen) was added into the cultures and the samples were incubated for 2 h at 37°C. One hundred microliters of the solution from each sample was transferred into a 96-well plate and the fluorescence was measured with an excitation wavelength of 544 nm and emission wavelength of 590 nm. In addition, the living and dead cells after treatment were observed using a Live-Dead® viability/cytotoxicity staining (Invitrogen).
Cell cycle analysis
One hundred thousand cells were plated into six-well plates and treated for 1, 4, and 7 days (n = 3). At each time point, cells were trypsinized and single-cell suspensions were prepared, fixed with ice cold 70% ethanol, and stored at −20°C overnight to resolve sub-G1 phase. Subsequently, the fixed cells were resuspended in 0.5 mL PBS containing propidium iodide (40 μg/mL), RNase (100 μg/mL), and 0.025% Triton-X100, and incubated at 37°C for 45 min before analysis using a BD FACSCanto™ II flow cytometer. Approximately 10,000–20,000 events were recorded for day 1 samples and 50,000 events for days 4 and 7 samples. The percentages of G0/G1, S, G2/M, and apoptotic phases of each sample were determined using Modfit LT software (Verity Software House, Inc.)
Quantitative reverse transcription–polymerase chain reaction
hPDCs were seeded at 4500 cells/cm2 in six-well plates and treated for 7, 14, 21, and 28 days (n = 3). Total RNA of each sample was isolated using Nucleospin® RNA extraction kit (Macherey-Nagel) and quantified using Nanodrop ND-1000 spectrophotometer (Thermo Scientific). Complementary DNA was synthesized by reverse transcription of 250 ng total RNA using the RevertAid™ H Minus First Strand complementary DNA synthesis kit with Oligo(dT)18 as primer (Fermentas). Sybr green quantitative polymerase chain reaction primers were designed to span an intron so that only RNA-specific amplification was possible (BMP-2F, 5′-GTATCGCAGGCACTCAGGTC-3′; BMP-2R, 5′-TTTTCCCACTCGTTTCTGGT-3′; OPNF, 5′-ACTGATTTTCCCACGGACCT-3′; OPNR, 5′-TCAGGGTACTGGATGTCAGG-3′; OCNF, 5′-GTGCAGCCTTTGTGTCCAA-3′; OCNR, 5′-GCTCACACACCTCCCTCCT-3′; RUNX2F, 5′-CGCATTCCTCATCCCAGTAT-3′; RUNX2R, 5′-GCCTGGGGTCTGTAATCTGA-3′). The reverse transcription–polymerase chain reaction was cycled in a Rotor-Gene sequence detector as follows: 95°C for 3 min, 40 cycles of 95°C for 3 s, and 60°C for 60 s. Each sample was tested in duplicate and compared with β-Actin expression (βACTF, 5′-CCCAGATCATGTTTGAGACCT-3′; βACTR, 5′-CCTCGTAGATGGGCACAGT-3′) as an external control that allowed normalization of results. Relative differences in expression were calculated using the 2−ΔCT method, and statistical significances were calculated by comparing to the 2−ΔCT of the negative controls (GM).
Mineralization assay
hPDCs were seeded at 4500 cells/cm2 and treated for 21 days (n = 4). At day 21, the cultures were rinsed with PBS, fixed with 4% formaldehyde, and then rinsed with distilled water before staining with an alizarin red solution (pH 4.2). Nonspecific staining was rinsed off carefully with distilled water. Quantification of calcium mineral deposits was performed by dissolving the dye with 10% cetylpyridinium chloride (in demineralized water) for 60 min at room temperature. Absorbance was measured spectrophotometrically at 570 nm. The mineralized nodules were analyzed by scanning electron microscopy coupled with an energy dispersive X-ray (EDAX) system (FEI XL30-FEG). Briefly, the cultures were rinsed twice with PBS, fixed with 2.5% glutaraldehyde (in PBS) for 1 h, and postfixed in 1% osmium tetroxide for 2 h before dehydrated in 50%, 75%, 95%, and 100% ethanol solutions. Finally, the samples were chemically dried with hexamethyldisilazane 43 for 3 min and gold sputtered before scanning electron micrograph (SEM) analysis.
Statistical analysis
Data are expressed as mean ± standard deviation or mean ± standard error of the mean. One-way analysis of variance (ANOVA) was performed to evaluate the statistical difference of means of multiple independent groups, and the unpaired Student's t-test (two-tailed) was performed to compare two independent groups, both by establishing the statistical significance at α < 0.05.
Results
Enhancement of hPDCs growth and phenotypic change by Ca2+ or Pi supplemented growth media in a time- and dose-dependent manner
The growth of CaP-treated hPDCs was evaluated by comparing the DNA content of each sample to that of GM treatment for each time point (Fig. 1A). At day 1, no statistical differences in DNA content was observed in cultures treated with Pi-supplemented growth media and OM; however, significantly lower DNA content was obtained in cultures treated with Ca2+ containing growth media (p < 0.05). After 3 days, C2 and P8 caused a significant increase in DNA content, which was comparable to OM treatment (p > 0.05). At day 7, C2–C4 and P4–P8 produced significantly higher DNA content (ANOVA, p < 0.05) and again were comparable to OM treatment (ANOVA, p > 0.05). After 14, 21, and 28 days of treatment, cultures treated with C4, P4, C8, and P8 induced the greatest increase in DNA content (as shown by the solid and dashed lines in Fig. 1A), which were statistically the same as that of OM except for cultures treated with P4 for 14 days and P8 at 21 days. The same concentration for Ca2+ and Pi (i.e., C4 and P4 or C8 and P8) appeared as the optimal concentrations that produced the highest DNA content at the same time point, meaning that C4 and P4 were the highest at day 14, then C8 and P8 at day 21, and C4 and P4 again at day 28. These observations suggest a lag before additional phases of cell mitosis under Ca2+ and Pi-induced cell proliferation. In summary, low [Ca2+] (i.e., C2–C4) induced significant cell growth at early time points (≤7 days), which were then overwhelmed by higher [Ca2+] at the later time points, whereas similar ranges of [Pi] (i.e., P4–P8) induced significantly higher cell growth at both early and late time points. The effect of vehicle (Vh) treatment on cell growth was minimal as no significant increase in DNA content was observed for any of the time points except at 21 days (data not shown), confirming that this growth effect was induced specifically by Ca2+ and Pi.
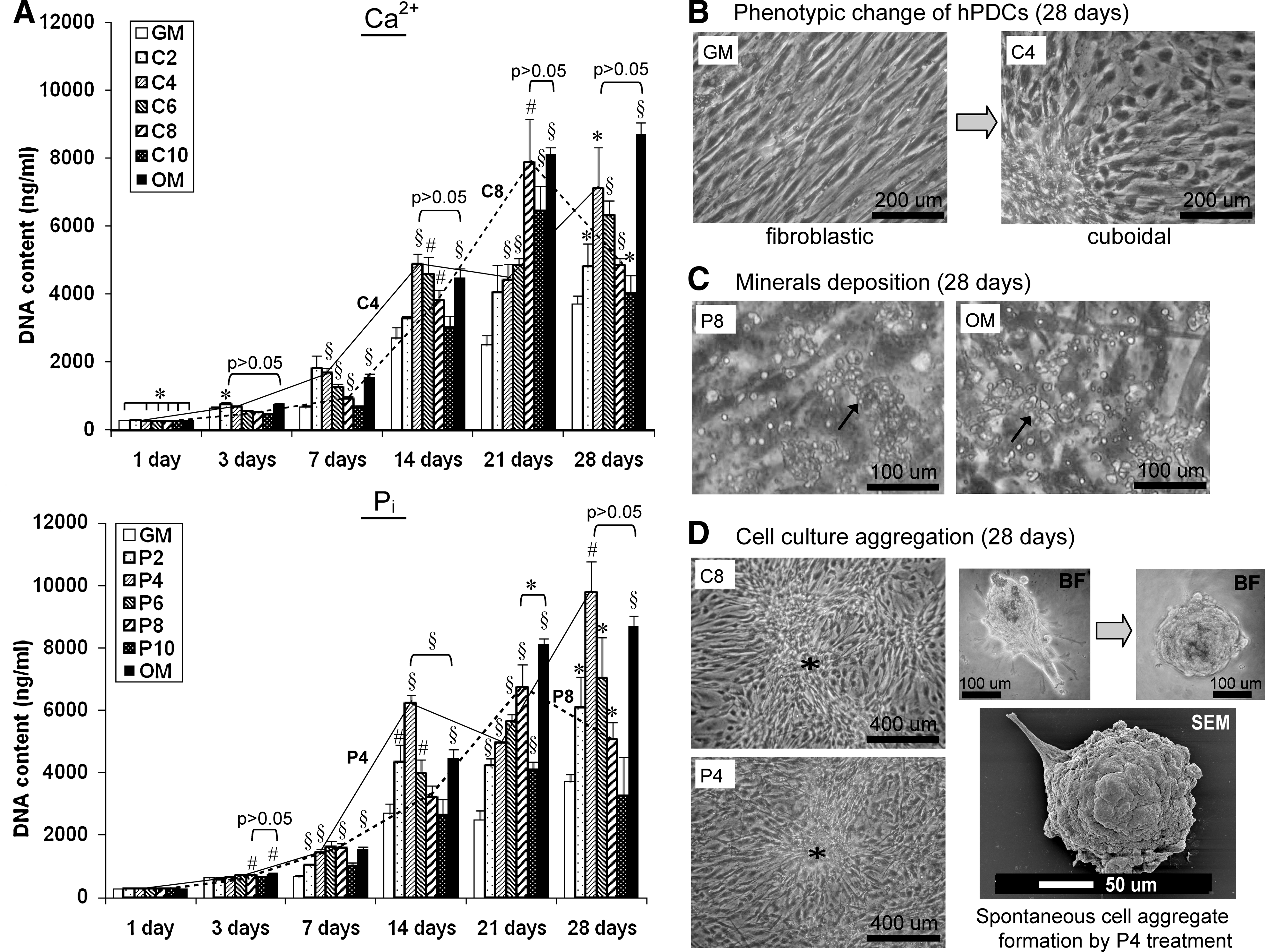
Treatment with various [Ca2+]- or [Pi]-induced
In addition, treatment with Ca2+ or Pi-supplemented growth media resulted in a dose- and time-dependent change in cell morphology from a fibroblastic to cuboidal phenotype, following treatment for 7 days and after (Fig. 1B and Supplementary Fig. S1; Supplementary Data are available online at www.liebertonline.com/tea). At 14 days, mineralization (arrows) was observed in cultures treated with P6 to P10 and in OM (Fig. 1C), whereas cell aggregation (*) was observed in cultures treated with various [Ca2+] as well as P4 (Fig. 1D). Spontaneous cell aggregate formation was also observed by P4 treatment at 28 days (as shown by brightfield and SEMs in Fig. 1D).
Short- and long-term synergistic effect of combined Ca-P treatments on hPDC growth
Upon simultaneous addition of Ca2+ and Pi supplemented growth media into cultures, nonphysiological precipitation of CaP occurred on the cell layer, which was proportionate to the concentration of ions used. Surprisingly, specific combinations of Ca-P at high concentrations, that is, C6P2, C8P4, C8P8, and C10P8, induced significantly higher DNA content than that of GM after 1 day of treatment and subsided at later time points (Fig. 2). This initial burst of cell growth was not observed in cultures treated with combined Ca-P solutions at lower concentrations. However, these lower concentrations, such as C2P2, C2P4, C4P2, C6P2, C8P2, and C10P2, resulted in a significantly higher DNA content compared to that of GM after 3 days of treatment and persisted to 28 days. Cultures treated with C4P10 induced significant cell growth only at day 3 and subsided at later time points. These findings indicate an inverse relationship between cell growth and the quantity of nonphysiological CaP precipitate formed upon the cell monolayer, and a high quantity of CaP precipitate may potentially jeopardize cell viability during long-term treatment. This is supported by additional data showing low metabolic activity in cultures treated with a high concentration of combined Ca-P-supplemented growth media (Supplementary Fig. S2).
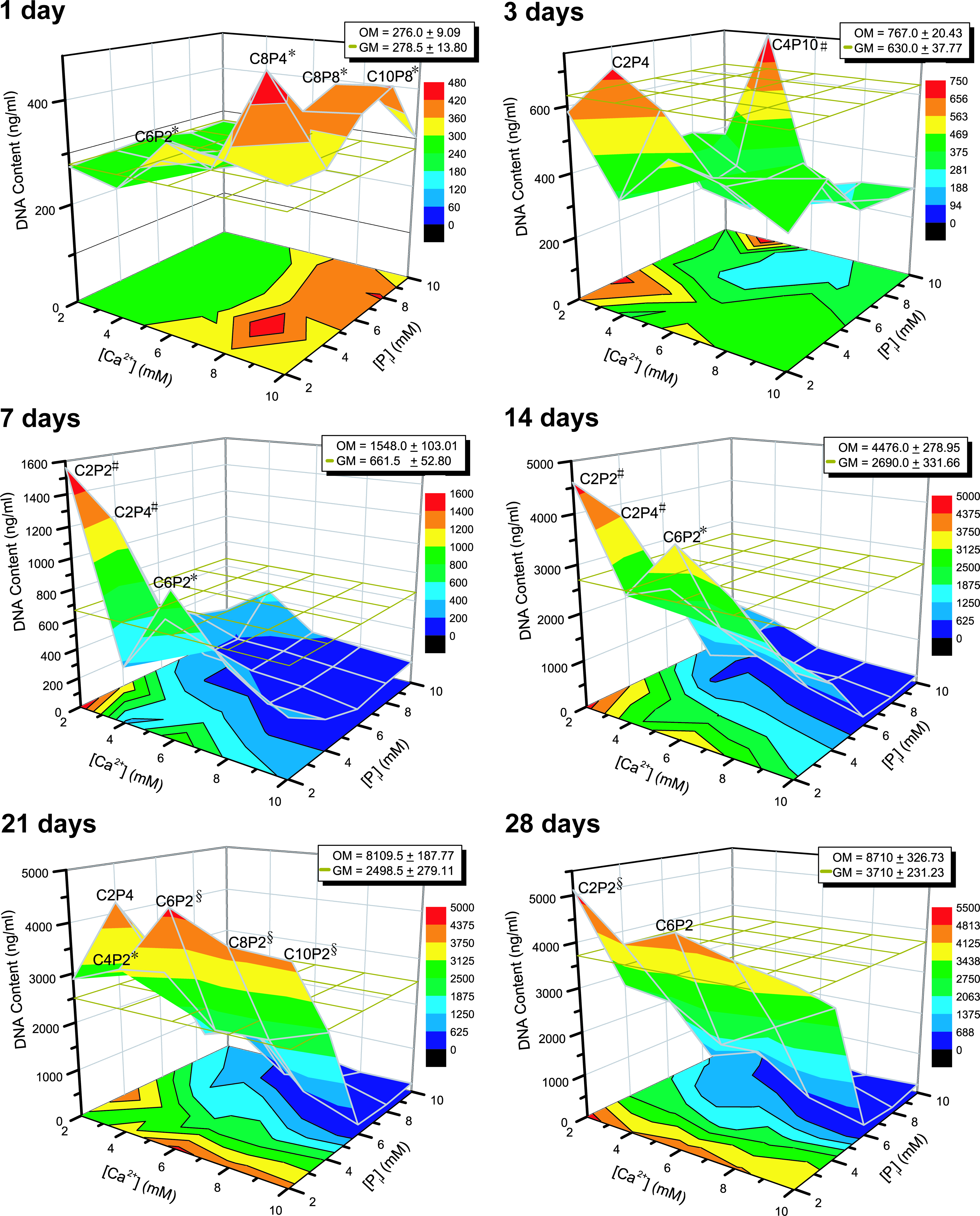
hPDCs growth in combined Ca-P solutions after treatment for 1, 3, 7, 14, 21, and 28 days. An initial burst of cell growth was observed for cultures treated with high concentrations of Ca-P, which subsided later on due to the adverse effect of the CaP precipitate formed on the cell monolayer. Lower concentrations of Ca-P, such as C2P2, C2P4, and C6P2, enhanced cell growth persistently during the entire culture period although in the presence of some amount of CaP precipitate. The mesh in the graphs indicates the DNA content (baseline levels) of the negative controls (GM). Mean ± SD (n = 4). Unpaired t-test (two-tailed): *p < 0.05, #p < 0.01, §p < 0.001. CaP, calcium phosphate. Color images available online at www.liebertonline.com/tea.
Encouragement of hPDC cell cycle progression in a time- and dose-dependent manner
Before synchronization, 90.6%, 5.1%, and 4.4% of hPDCs were found to be in the resting (G0/G1), synthesis (S), and mitotic phase (G2/M), respectively (Fig. 3A). After synchronization, 95.5% of the cells were in the G0/G1 (p < 0.01) with <4% of cells in the S and only 0.7% of cells in G2/M phases (p < 0.001). Morphologically, the synchronized cells had a shrunken cytoplasmic appearance. These results suggest that serum deprivation is an effective method in synchronizing cells into one similar phase in the cell cycle. Upon replenishing cells in a serum-containing medium (i.e., GM) for 1 day, significantly more cells were induced into the S (14.1%) and G2/M (12%) phases, leaving 74% of cells in the G0/G1 phase (Fig. 3B). This recovery of cell metabolism and progression to mitotic activity from serum deprived to serum-rich conditions was significantly retarded in a dose–response manner by C6–C10 after 1 day of treatment (p < 0.05), causing more cells to remain in the resting phase as compared to GM treatment. These results explained the earlier finding as to why a significantly lower DNA content was obtained (as compared to that of GM) when cells were treated with Ca2+ containing growth media for 1 day. No retardation of mitotic activity was observed for Pi-treated cultures after replenishment in serum-rich conditions for 1 day; indeed, P4 caused a significant higher percentage of cells in the S phase (20.6% ± 1.67%) as compared to that of GM (p < 0.05) and OM (p < 0.01) treatments. At day 4, a relatively higher percentage of cells were moving into the resting phase (Ca2+: 93.0% ± 1.17%; Pi: 92.7% ± 1.42%) as compared to day 1, but with a significant dose-dependent increase in the S and G2/M phases for both treatment types (Fig. 3C). This significant dose-dependent increase in the S and G2/M phases was observed again when cultures were treated with Ca2+ and Pi for 7 days, the effects of which were more prominent than that of OM (Fig. 3D).

Cell cycle analysis of hPDCs
On the basis of the DNA measurement data, the effects of specific combined Ca-P-supplemented growth media (i.e., C4P10, C8P4, and C10P8) on cell cycle progression were evaluated (Fig. 4). The results showed that these combined Ca-P solutions pushed a significantly higher number of cells into the G0/G1 phase (ANOVA, p < 0.001) at day 1, and moved the cells into a higher percentage of S (ANOVA, p < 0.05) and G2/M (ANOVA, p < 0.001) phases at day 4 (except for C10P8) (Fig. 4A). However, ∼40%–50% of the total analyzed cells were found to be apoptotic, which can be seen from the microscopic images showing apoptotic cells (Δ). When treating hPDCs with a gradual increase in [Ca2+] at constant 4 mM of Pi (P4) for 1 day, the percentage of cells in the S phase dropped dramatically from a high value (C2P4: 30.3% ± 1.91%) (p < 0.01) down to 9.5% ± 6.51% (for C10P4) (Fig. 4B). This was accompanied by a large increase in cell apoptosis from 58.8% ± 2.98% (for C2P4) to as high as 90.0% ± 3.27%, together with no cells in the G2/M phase, at the extreme case (i.e., C10P4). This was probably due to the formation of high amount of nonphysiological CaP precipitate (*) in the combined Ca-P treatments that had affected cell viability. At day 4, opposite responses were observed where the percentage of cells in the S phase increased fourfold (from 5.3% ± 3.84% to 23.0% ± 2.40%) with an increase in [Ca2+], and no significant changes in percentage of cells that were in the G2/M phase and apoptotic were observed. Subsequently, at day 7, C6P4 induced the highest percentage of cells in the S phase (45.0% ± 5.09%), which was threefold higher than other conditions with statistically the same percentage of apoptotic cells (41.7% ± 2.62%). These data indicate that Ca2+ and Pi individually or in combination induce cell proliferation by regulating cell cycle progression; additionally, the combined Ca-P treatment also induces cell apoptosis, which is attributed to the formation of nonphysiological CaP precipitate.

Cell cycle analysis of hPDCs after treatment with
Treatment of Ca2+ or Pi significantly upregulated osteocalcin, osteopontin, BMP-2, and Runx2 gene expression of hPDCs in a time- and dose-dependent manner
Cultures treated with C2, C4, C8, P4, P6, and P8 for 7, 14, 21, and 28 days were selected to assess the individual effect of Ca2+ and Pi on the dynamic expression of osteogenic gene markers (Fig. 5). Osteocalcin (OCN), an osteoblast-specific marker, was significantly upregulated by C8 as early as after 7 days of treatment (p < 0.05), and continued to rise significantly to approximately four to fivefold higher than that of GM and OM treatments at later time points [Fig. 5A (i)]. Cultures treated with C2 and C4 also displayed a significant upregulation of OCN gene expression starting from 14 till 28 days, but at relatively lower levels than that of C8 treatment. A significant dose–response and time-dependent upregulation of OCN gene expression was also obtained for cultures treated with P4–P8 as compared to that of GM (ANOVA, p < 0.001) and OM (ANOVA, p < 0.001) after 21 days of treatment [Fig. 5A (ii)]. Comparatively, treatment with C2 for 14 days and C4 for 28 days induced a relatively mild upregulation of osteopontin (OPN) gene expression (p < 0.05 in both cases) [Fig. 5B (i)], whereas treatment with P8 induced a significantly higher expression of OPN gene than that of GM and OM treatments at 14 (p < 0.05) and 21 days (p < 0.01) [Fig. 5B (ii)]. A persistent upregulation of BMP-2 gene expression by both Ca2+ and Pi treatments (in a dose- and time-dependent manner) was obtained as early as from 7 to 28 days, which the effect was superior to that of GM and OM treatments [Fig. 5C (i) and (ii)]. The effect of Ca2+ on Runx2 gene expression was relatively less prominent for the entire 28 days of treatment [Fig. 5D (i)]; however, Pi did upregulate Runx2 expression at later time points in a dose–response pattern especially when cells were treated with P8 for 14 (p < 0.05) and 28 days (p < 0.01) [Fig. 5D (ii)].

Osteogenic gene expression of hPDCs after treatment with (i) Ca2+- or (ii) Pi-supplemented growth media for 28 days. (
Long-term treatment with combined Ca-P significantly upregulated OCN, OPN, and BMP-2 but downregulated Runx2 gene expression of hPDCs in a time-dependent manner
The effect of combined Ca-P treatment on osteogenic gene expression of hPDCs was studied by treating the cells with the growth medium containing 6 mM of Ca2+ and 4 mM of Pi (i.e., C6P4) for 7, 14, and 21 days. This combined Ca-P condition was selected based on experimental data showing good cell viability, which was reflected by high metabolic activity and by live-dead staining over 28 days of treatment (Supplementary Figs. S2 and S3). A time-dependent increase in OCN, OPN, and BMP-2 gene expression was observed from 7 to 21 days in the presence of C6P4, which was significantly higher (12-fold higher for OCN; 20.6-fold for OPN; 13-fold for BMP-2) than that of the negative control (GM) after 21 days of treatment (Fig. 6). However, this combined Ca-P treatment had significantly downregulated the Runx2 gene expression (p < 0.05) when compared to the negative control at 21 days, although this effect was not statistically significant at earlier time points. These results indicate that during long-term combined Ca-P treatment, an environment that is enriched with Ca2+ and Pi (which includes its precipitation) has a promising effect on the regulation of hPDC osteogenic gene expression.
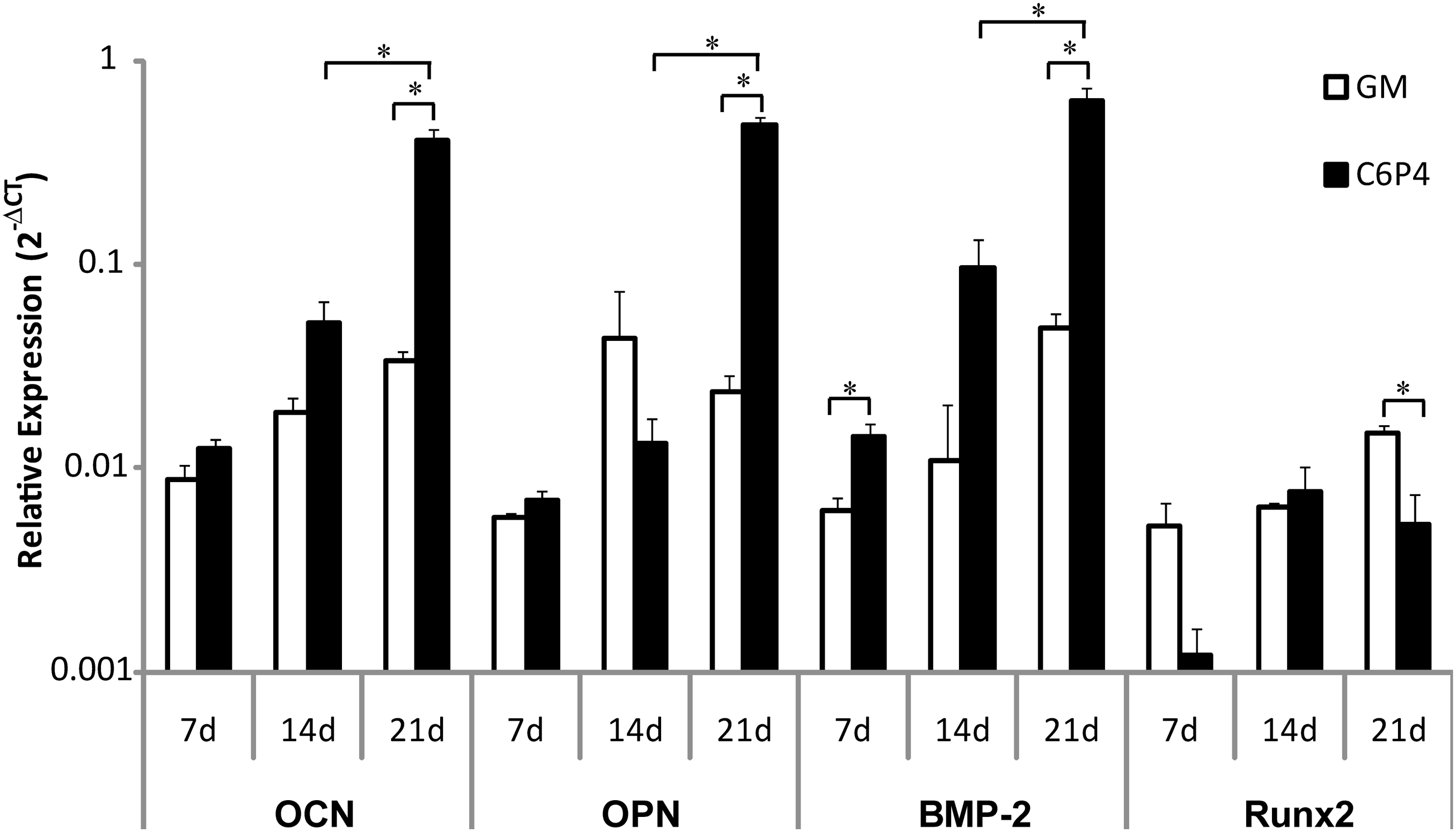
Osteogenic gene expression of hPDCs under combined Ca-P treatment with the growth medium containing 6 mM of Ca2+ and 4 mM of Pi (i.e., C6P4) for 7, 14, and 21 days. Expression of OCN, OPN, and BMP-2 was upregulated over the time (as compared to negative controls [GM]) and became statistically significant at 21 days. No significant effect of C6P4 treatment on Runx2 gene expression at earlier time points was found, but it was significantly downregulated at 21 days. Data shown are relative expression of the gene of interest normalized to β-Actin. Mean ± standard error of the mean (n = 3, in duplicate measurements). Unpaired t-test (two-tailed): *p < 0.05. BMP-2, bone morphogenetic protein-2; OCN, osteocalcin; OPN, osteopontin.
Mineralization of hPDCs culture was Pi-dependent and exhibited matrix vesicle-like structures as well as cell–CaP mineral interaction
Calcium mineral deposition by hPDCs after treatment with Ca2+, Pi, and combined Ca-P for 21 days was observed by alizarin red staining (Supplementary Fig. S4). The staining showed no mineral deposition for any culture treated with Ca2+ or with the Vh or GM treatments. However, positive staining was demonstrated in cultures treated with 2–10 mM of Pi, where the amount of deposited minerals was in a dose-dependent manner. These mineral depositions were formed mainly at the center of the well and had a different physical appearance than that of OM-treated samples, which the mineral deposition was homogenously distributed. Cultures treated with combined Ca-P supplemented growth media resulted in an intense staining of the mineral deposits that were proportionate to the concentration of the ions. These mineral deposits were believed to be physiological and cell-mediated, as nonphysiological Ca-P precipitate formed in the absence of cells could be rinsed off easily. Quantitatively, cultures treated with Pi caused a significantly higher mineral deposition as compared to GM-treated culture (Fig. 7A), whereas cultures treated with combined Ca-P solutions resulted in almost fivefold higher mineral deposition than that of GM treatment. However, this effect was only influenced by increasing [Pi], suggesting that calcium mineral deposition by hPDCs is Pi dependent.

Mineralization of hPDCs after treatment with Ca2+, Pi, or combined Ca-P supplemented growth media for 21 days.
SEMs showed no calcium mineral deposits in GM and Ca2+-treated cultures, except for the C10-treated culture, where some matrix vesicle-like structures (mv) of ∼400–600 nm in diameter were observed (Fig. 7B). Similar matrix vesicle-like structures (mv) were observed in abundance in cultures treated with OM and P4. However, EDAX analysis suggested that only matrix vesicle-like structures present in OM-treated cultures contained calcium (Ca2+) and phosphorus (P) elements. This was probably due to the presence of low amounts of Ca2+ elements in the C10 and P4-treated cultures that were masked by the high percentage of gold coating (Au). The mineral structures that were observed in cultures treated with P6, C6P4, and C10P4 were confirmed by EDAX analysis to be CaP minerals, where established cell–mineral deposit interactions were clearly visible. CaP precipitates formed in the C10P4 solution without the presence of cells were purely a nonphysiological mineral deposit, which had a different physical morphology and crystal structure than that of cell-containing treated samples. Trace amounts of Pi but not Ca2+ were detected in GM and C6-treated cultures. These results suggested that mineralization of hPDCs was enhanced by Pi but not Ca2+, and the mineral deposition process was cell mediated, provided that the threshold amount of CaP precipitate formed on the cell layer did not exceed or compromise the cell growth.
Discussion
The effect of extracellular Ca2+ and Pi on in vitro osteoblast behavior has been investigated with reports of enhanced cell proliferation, upregulation of osteogenic markers, and mineralization.44,45 However, these investigations were mainly carried out on differentiated osteoblasts from different species or osteoblastic cell lines (e.g., MC3T3-E1 and 2T3 cells), but experiments using human progenitor cells are still limited.46–48 In this study, we have performed the experiments on clinically relevant MSCs, that is, hPDCs, which have been shown to be multipotent 49 and a critical cell type for bone defect regeneration.50,51 Further, this cell type was reported to be physiologically responsive to an in vivo dynamic [Ca2+] and [Pi] fluctuating environment and involved in the homeostasis of these ions at the bone surface. 52
In the event of direct osteogenesis (membranous bone formation) by MSCs the following processes occur: (1) cell proliferation, (2) cell aggregation, (3) cell differentiation, (4) angiogenesis, (5) mineralization, and (6) bone formation, with the dynamic expression of osteogenic transcription and growth factors in a tightly regulated spatial and temporal pattern.53,54 In our study, we have shown that Ca2+ and Pi stimulated cell growth in a time- and dose-dependent manner to a level comparable to the mitogenic effect of a generally used standard osteogenic medium. We observed phenotypic changes of hPDCs, including a phenotypic change from a fibroblastic to cuboidal morphology as well as cell aggregation. These findings suggested a potential role of Ca2+ and Pi in stimulating cell proliferation and cell aggregation, which is important for the production of a critical cell mass that is required to initiate bone tissue formation. 55 This mitogenic effect by Ca2+ and Pi was corroborated by cell cycle analysis. At appropriate concentrations and treatment time, Ca2+ and Pi dynamically influence the cell cycle progression, mainly by inducing cells to enter into DNA synthesis and the phase of cell mitosis.
At the molecular level, Ca2+ has been shown to affect cell cycle progression, 56 osteoblast proliferation, 57 and differentiation 58 through the calcium/calmodulin signaling pathway. 59 Additionally, Ca2+ has been shown to increase DNA synthesis with concurrent production and secretion of insulin-like growth factor-1, which was found to be independent of Ca2+ channel signaling. 60 Similar stimulatory effects on osteoblastic cell proliferation by 2–4 mM of Pi was also reported. 61 These investigations clearly support our data on Ca2+ and Pi-enhanced hPDCs growth, and they were within the same range of concentrations. However, high [Pi] (>5 mM) has been associated with osteoblast apoptosis due to the failure of mitochondrial function, which could be abolished by inhibiting Pi transport using Na-Pi cotransporter (Pit-1) inhibitors.62–64 However, a critical function of Pit-1 in cellular proliferation, which is independent of its Pi transport activity, has been identified. 65
Alkaline phosphatase (ALP) plays an important role in Pi homeostasis by converting extracellular pyrophosphate (ePPi, an inhibitor of mineralization) to Pi. 66 hPDCs contain osteoprogenitor cells that inherently show no ALP activity until exposure to osteogenic stimuli such as OM. 49 In this study, we have measured the ALP activity of each sample over 28 days but no significant increase in ALP secretion was found in Ca2+, Pi, or in combined Ca-P treatments when compared to the negative controls (GM) (data not shown).
OPN has been described as a determinant of carbonated hydroxyapatite crystal formation for mineralization as well as facilitating osteoclast attachment to the matrix, whereas OCN is involved in the binding of calcium to the extracellular matrix for the regulation of the growth and size of mineral crystals 67 and is also a marker for osteogenic differentiation. 68 In vivo data on OPN and OCN knockout mice support these findings.69,70 In our study, we have shown a significant upregulation of OCN and OPN expression in Ca2+, Pi, and combined Ca-P-treated cultures in a specific time- and dose-dependent fashion. These findings indicate that Ca2+ and Pi, individually or in combination, induce the expression of OCN and OPN, which are essential in the regulation of extracellular matrix mineralization. The regulation of OPN expression by Pi has previously been reported, where OPN gene transcription in cementoblasts by extracellular Pi requires a functional glucocorticoid receptor. 71 Indeed, osteoblasts have been shown to have an autonomous Pi handling system (the STC1-Pi-1-Pi-sensing system) that stimulates OPN expression and mineralization. 72 Further, our data show an upregulation of the master gene of bone formation, Runx2, in cultures treated with Pi and OM for >14 days but not by Ca2+ or combined Ca-P treatments. This suggests that Pi may play a role in differentiating hPDCs from a MSC into the osteochondroprogenitor lineage. This observation can be linked to a study reporting a Pi-induced nuclear export system for Runx2 in osteoblasts. 73
Previous studies published by our group have shown that implantation of collagraft seeded with hPDCs induced robust bone formation ectopically, 51 which required the presence of CaP and the activation of BMP and Wnt signaling. 74 In this study, we have shown a strong and persistent upregulation of BMP-2 expression (in a dose- and time-dependent manner) in both Ca2+ and Pi as well as in combined Ca-P-treated cultures. These findings are in line with our previous in vivo findings and other studies that reported a similar phenomenon.75,76 Indeed, this is a very encouraging finding that potentially reveals a possible mechanism of the osteoinductive response triggered by CaP. Although entrapment of endogenous BMPs onto CaP-based implants has been suggested as the mechanism by which bone formed within a CaP biomaterial persists without resorption, 15 compelling evidence for this is still lacking. Here, we would like to provide a hypothesis that osteoinduction by CaP could potentially be attributed to the direct effect of the released Ca2+ and Pi on BMP-2 expression. This may lead to the generation of BMP-2 secreting cells, which serves as a BMP-2 autocrine/paracrine osteoinduction model that drives a cell-CaP carrier toward effective ectopic bone formation. How Ca2+ and Pi enhance BMP-2 gene expression is unknown, but recent studies suggest that BMP-2 protein is necessary to stimulate Pi transport by enhancing expression of Pit-1. 39 Second, it was reported that Ca2+ reciprocally regulates BMP-2 and the BMP antagonist Noggin secretion via calcium-sensing receptor, although this was only demonstrated in myofibroblasts. 77
In this study, we have provided evidence on the osteoinductive effect of CaP. This effect encompasses the key stages of in vitro osteoblastic differentiation, including proliferation, cell aggregation, osteogenic gene expression, and matrix mineralization, as detailed in Figure 8. Additionally, BMP-2 expression is induced in a time- and dose-dependent manner during treatment with Ca2+ and Pi. This has led to the hypothesis that CaP-induced bone formation may be governed by the activation of BMP signaling, which could potentially serve as an autocrine/paracrine osteoinduction loop (Fig. 8). If correct, this hypothesis would reduce the need to functionalize CaP-based biomaterials with osteoinductive molecules, which is an area that has previously been highlighted as a challenge in developing biomimetic matrices capable of inducing bone formation. 78 After all, we believe that the osteoinductivity of CaP is not solely a Ca2+ and Pi-dependent process, but is rather a network of in vivo processes that coherently fine-tune the cellular response, both spatially and temporally, toward coordinated cell proliferation, differentiation, mineralization, and eventually bone formation.
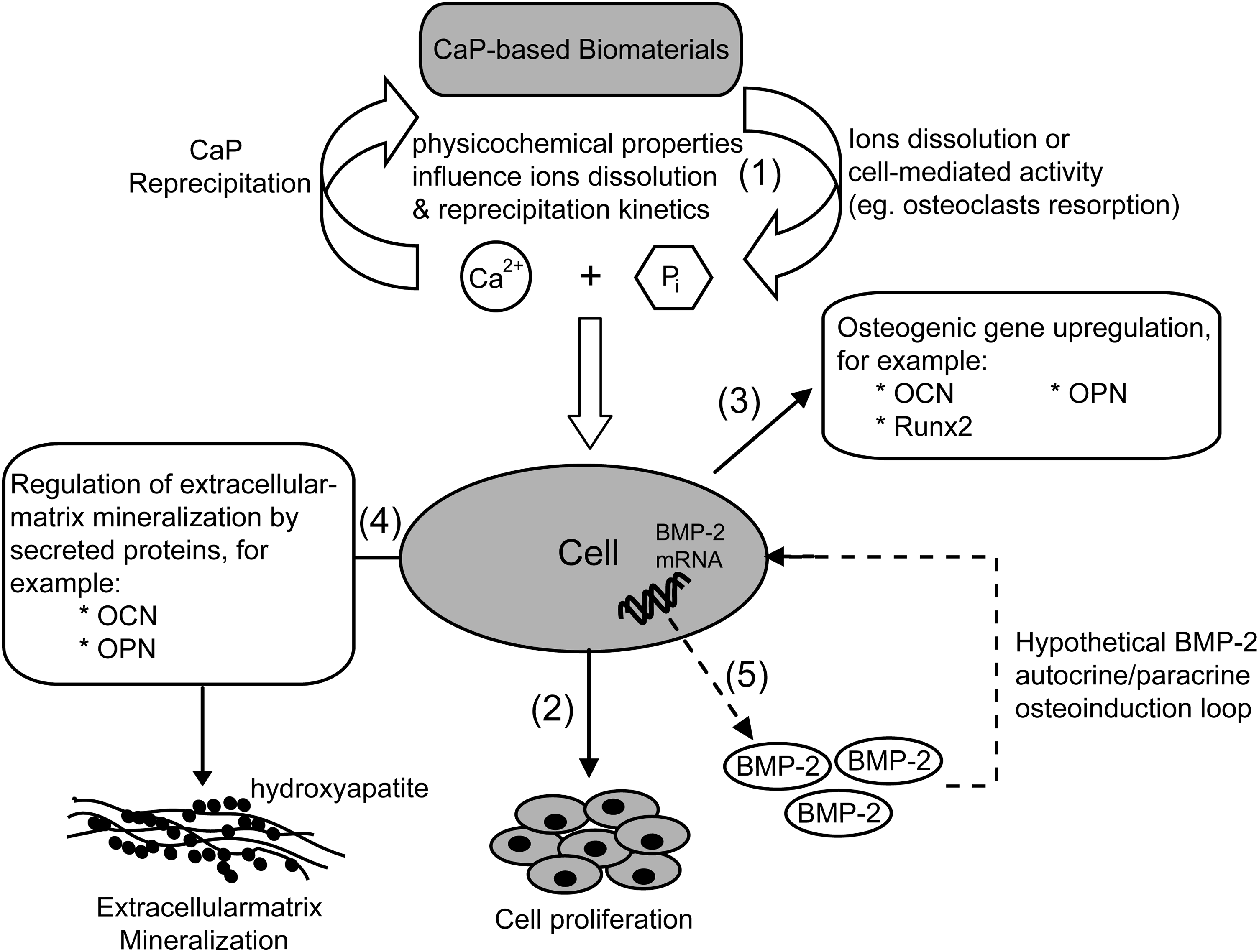
Proposed hypothetical mechanism of CaP osteoinductivity. (1) Depending on the physicochemical properties of a CaP-based biomaterial, as well as the surrounding cell-mediated events, appropriate [Ca2+] and [Pi] are released from the bulk material into the extracellular compartment and reprecipitate partly onto the material surface to achieve equilibrium. (2) At appropriate ion concentrations, the released Ca2+ and Pi stimulate cell proliferation through modulation of cell cycle progression. (3) Upregulation of osteogenic gene expression, such as OCN, OPN, and Runx2, by the released Ca2+ and Pi. (4) Physiological mineralization of the extracellular matrix is tightly regulated by secreted proteins such as OCN, OPN, and collagen type-1. (5) The released Ca2+ and Pi upregulate the BMP-2 gene expression and may lead to the secretion of BMP-2 proteins, which in turn serves as an autocrine/paracrine osteoinduction loop that activates the BMP signaling pathway and drives the cell-CaP construct toward effective bone formation.
Conclusion
Through an in vitro biomimetic model using Ca2+ and Pi solutions tested on hPDCs, we have partially probed the roles of each ion on the osteoinductivity of CaP-based biomaterials. We have shown that Ca2+ and Pi both induced cell proliferation by pushing cells into S-synthesis and M-mitotic phases in a dose- and time-dependent manner. This mitogenic effect was believed to be essential for achieving a critical cell mass that would initiate bone tissue formation. Osteogenic genes that are important for the regulation of physiological mineralization of the extracellular matrix (e.g., OCN and OPN) were upregulated by Ca2+ and Pi individually, or in combination, in a dose- and time-dependent manner. Relatively, Pi showed a more pronounced effect on hPDCs differentiation into the osteochondroprogenitor lineage than the effect of Ca2+ or combined Ca-P treatments. Most encouragingly, both Ca2+ and Pi strongly and persistently upregulated BMP-2 gene expression, thus potentially leading to the generation of BMP-2 secreting cells, which we believe could serve as an autocrine/paracrine osteoinduction loop that autonomously activates the BMP signaling pathway and drives the cell-CaP construct toward effective bone formation.
Footnotes
Acknowledgments
The authors are grateful to Carla Geeroms, Kathleen Bosmans, and Isabelle De Wit for the excellent technical assistance. The authors also would like to thank Viktor Van Duppen for the technical and data analysis assistance on the cell cycle experiment. This work is funded by the K.U.Leuven IDO project 05/009—QuEST and is part of Prometheus, the Leuven Research & Development Division of Skeletal Tissue Engineering of the Katholieke Universiteit Leuven: ![]() .
.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.