
Abstract
The mechanisms driving bone marrow stem cell mobilization are poorly understood. A recent murine study found that circulating bone marrow-derived osteoprogenitor cells (MOPCs) were recruited to the site of recombinant human bone morphogenetic protein-2 (BMP-2)-induced bone formation. Stromal cell-derived factor-1α (SDF-1α) and its cellular receptor CXCR4 have been shown to mediate the homing of stem cells to injured tissues. We hypothesized that chemokines, such as SDF-1, are also involved with mobilization of bone marrow cells. The CD45− fraction is a major source of MOPCs. In this report we determined that the addition of BMP-2 or SDF-1 to collagen implants increased the number of MOPCs in the peripheral blood. BMP-2-induced mobilization was blocked by CXCR4 antibody, confirming the role of SDF-1 in mobilization. We determined for the first time that addition of SDF-1 to implants containing BMP-2 enhances mobilization, homing of MOPCs to the implant, and ectopic bone formation induced by suboptimal BMP-2 doses. These results suggest that SDF-1 increases the number of osteoprogenitor cells that are mobilized from the bone marrow and then home to the implant. Thus, addition of SDF-1 to BMP-2 may improve the efficiency of BMPs in vivo, making their routine use for orthopaedic applications more affordable and available to more patients.
Introduction
In adult mammals, most hematopoetic stem cells (HSCs), endothelial progenitor cells (EPCs), and MSCs reside in the bone marrow, with a few HSCs and EPCs circulating in the peripheral blood. Circulating MSCs are usually not detectable. 4 Osteoblast-lineage cells have been reported to be present in the circulating blood of humans. 5 However, the origin and function of the circulating osteoblastic cells are uncertain. A recent murine study found that circulating bone marrow-derived osteoprogenitor cells (MOPCs) were recruited to the site of bone formation induced by BMP-2. 6 These authors found that 50% of the osteoblasts generating bone in response to BMP-2 were derived from the endogenous circulating MOPCs with the remaining cells probably derived from tissue-based or perivascular progenitors. 7 The CD45− fraction of mononuclear cells in bone marrow was mobilized within 7 days and represented the major source of circulating MOPCs that provide mature osteoblasts to peripheral tissues. The specific mechanism of the cell mobilization was not identified.
The factors that drive bone marrow stem cell mobilization have been unclear, but one of the earliest consequences of a bone fracture or surgical incision is local tissue hypoxia. Multipotential MSCs are mobilized into peripheral blood by experimentally induced hypoxia and myocardial infarction.8,9 The chemokine stromal cell-derived factor-1α (SDF-1α), although most studied for its role in cell homing, has also been linked to mobilization of EPCs and HSCs in rodents.10,11 SDF-1 is a chemokine that entraps progenitor cells expressing its receptor, CXCR4, in a variety of tissues. 12 Changes in circulating CD45−/CD34− MSCs in patients after myocardial infarction or burns have been associated with elevated levels of plasma SDF-1 within 3 days.13,14 Recently, it has been demonstrated that CD18, a protein known to exist only on hematopoetic cell surfaces, is also highly expressed on the progenitor cells and plays a critical role in the osteogenesis of bone marrow stromal stem cells. 15 Although it has not been reported if MSCs mobilized by SDF-1 also have osteogenic potential, these findings prompted us to determine if SDF-1 plays a role in mobilization of MOPCs from bone marrow.
In this study, we test the hypothesis that chemokines, such as SDF-1, are critical for in vivo responsiveness to BMP-2 because they both mobilize bone marrow precursor cells from the marrow into the peripheral blood and exert homing effects that direct the osteoblast progenitors to the site of new BMP-2 induced bone formation. Chemokines, while increasingly studied in cardiac, liver, and dermal tissue regeneration, represent a new focus for BMP-induced bone formation.
Materials and Methods
Animals and ectopic bone formation assay
After approval by the Institutional Animal Care and Use Committee, male athymic rats were obtained from Harlan Laboratories (Indianapolis, IN) at 6–10 weeks of age and housed under sterile conditions. The rats were anesthetized using an appropriately sized nose cone to deliver 1%–2% isoflurane mixed with oxygen at a flow rate of 0.5–1 L/min and maintained during surgery with the same dose. Surgery was performed with the animal on a circulating water heating pad. For mobilization and homing experiments one 1–2 cm incision was made on the chest of each rat, and subcutaneous pockets were created. Implants (collagen pellets) contained 100 μL of recombinant human BMP-2 (rhBMP-2; 5 μg; Medtronic, Minneapolis, MN) alone or combined with 100 μL SDF-1 (10 μg; R&D Systems, Minneapolis, MN). BMP-2 and/or SDF-1 were loaded using a pipette (Rainin, Oakland, CA) onto the sterile collagen disc in a biosafety cabinet and taken to the surgical suite in a sterile container. Each animal received one subcutaneous implant on the chest. Rats awakened from anesthesia rapidly were housed as two/cage, and were fed ad libitum. For ectopic bone formation experiments, all procedures were as described above except that four subcutaneous implants were placed on the chest of each rat. Rats were euthanized 4 weeks later and the implants were removed for palpation, radiography, and histology. The amount of bone in each implant was scored blinded based on the volume of an implant that on X-ray is mineralized (0 = no bone; 1 = less than 25% of the implant contains bone; 2 = 25%–49% of implant contains bone; 3 = 50%–74% of the implant contains bone; 4 = 75%–99% of implant contains bone; 5 = 100% or greater of the original implant size contains bone). Specimens analyzed by histology were fixed in formalin, embedded in methyl methacrylate, and stained with Gomori's one-step Trichrome.
Isolation of peripheral blood mononuclear cells
Peripheral blood was taken by cardiac puncture from the heart of athymic rats. Light-density mononuclear cells were isolated by Ficoll-Paque premium centrifugation (GE Healthcare, Uppsala, Sweden) according to the manufacture's protocol. Red blood cells were removed by re-suspending in ACK lysing buffer and centrifuging at 1000 rpm for 10 min. The supernatant was carefully removed and the cell pellet resuspended in magnetic cell sorting (MACS) bovine serum albumin (BSA) buffer supplemented with 2 mM ethylenediaminetetraacetic acid (Miltenyi Biotec-GmbH, Auburn, CA). The cells were washed by centrifuging at 1000 rpm for 10 min and resuspended in MACS BSA buffer.
Measurement of the mobilized CD45− population within peripheral blood mononuclear cells by flow cytometry
The isolated peripheral blood mononuclear cells (PBMNCs) were incubated with fluorescein isothiocyanate (FITC)-conjugated mouse anti-rat CD45 monoclonal antibody (BD Biosciences Pharmingen, San Jose, CA) for 30 min at 4°C in the dark. A conjugate control tube contained the FITC-conjugated mouse immunoglobulin G isotype control. After incubation, cells were washed with 10 volumes of phosphate-buffered saline (PBS) and centrifuged at 1000 rpm for 5 min. The supernatant was removed and the cell pellet was resuspended in 500 μL of PBS. Fluorescence-assisted cell sorting (FACS) analysis was performed with a FACS Calibur (Becton Dickinson, San Diego, CA) using Cell Quest software to determine the percentage of CD45− cells contained within the PBMNC population.
Enrichment of circulating MOPCs
To enrich the PBMNC population for CD45− cells, which contain predominately MOPCs, we used MACS Indirect Microbeads (Miltenyi). First, the PBMNCs were stained with the CD45 FITC-conjugated antibody according to the manufacturer's recommendation. Ten million cells were re-suspended in 100 μL degassed buffer (PBS, 0.5% BSA, and 2 mM ethylenediaminetetraacetic acid), mixed well, and incubated for 10 min in the dark at 4°C. Cells were washed to remove unbound primary antibody. Second, 10 million cells were resuspended in 90 μL of buffer and incubated for 15 min at 4°C with 10 μL of anti-FITC microbeads. The cells were washed, the supernatant was completely removed, and the cells were resuspended in 500 μL of buffer. Third, MACS LD columns were used on a MidiMACS separator (Miltenyi) to enrich the fraction depleted of CD45+ cells. Five hundred microliters of cell suspension was applied onto the prewashed LD column; the column was washed with 2 mL degassed buffer and the total effluent was collected.
Differentiation of MOPCs to mature osteoblasts
PBMNCs from animals implanted for 5 days with collagen pellets containing 5 μg BMP-2 were enriched for CD45− cells and plated at 1 × 104 cells/cm2 in Dulbecco's modified Eagle's medium supplemented with heat inactivated10% fetal bovine serum (Hyclone, Logan, UT) on Fibronectin-coated plates (BD Biosciences, Bedford, MA). Cells were cultured at 37°C in 5% CO2 under humidified conditions. To induce osteoblast differentiation the cells were treated with 100 ng/mL rhBMP-2, 50 μg/mL ascorbic acid (Sigma-Aldrich, St. Louis, MO), and 10 mM beta glycerophosphate (Sigma-Aldrich) for 21 days. Control cultures were grown in the same medium, but without BMP-2. The medium was changed twice weekly. Alkaline phosphatase mRNA levels were assessed on day 3. After 21 days, cultures were fixed and stained by Von Kossa staining; nodules were counted using a semi-automated computerized video image analysis system (Optimax 5; Optimax, Hollis, VT).
Chemokine in vitro migration assay
To demonstrate that the MOPCs mobilized by BMP-2 into the peripheral blood were also capable of migrating toward chemotactic agents, we collected peripheral blood from rats 5 days after BMP-2 (5 μg) implantation and generated a CD45− enriched population of MOPCs as described above. About 6 × 105 cells (3 million cells/mL stock) in 200 μL medium were loaded onto transwell filters (Corning, Corning, NY). Filters were carefully transferred to another well containing 2 mL of medium with 100 ng/mL BMP-2 or 1000 ng/mL of SDF-1. After a 4 h incubation at 37°C the upper chamber was removed and the cells in the bottom chamber were collected, washed, and re-suspended for counting using a hemocytometer.
Differentiation of pleuripotent C2C12 cells
C2C12 cells were seeded at 2 × 104 cells/cm2 and grown overnight in Dulbecco's modified Eagle's medium containing 10% nonheat inactivated fetal bovine serum (Hyclone, Thermo Scientific, Waltham, MA) at 37°C under humidified conditions in 5% CO2. Cells were then treated with 50 μg/mL ascorbic acid, 5 mM beta-glycerophosphate, and 100 ng/mL BMP-2 alone or in the presence of 50 ng/mL SDF-1 for 24 h. RNA was harvested on day 1 for analysis of osterix mRNA levels by real-time reverse transcription (RT)-polymerase chain reaction (PCR).
Real-time RT-PCR
Total RNA from each in vitro sample was extracted using the RNeasy kit as specified by the manufacturer (Qiagen, Valencia, CA). The isolated total RNA was treated with DNAase 1 (Qiagen) to remove DNA contamination and the concentration of the isolated RNA was spectrophotometrically determined. Two micrograms of total RNA was reverse transcribed using Mastermix containing 40 units RNase Inhibitor and 125 units MultiScribe (Applied Biosystems, Foster City, CA). Real-time PCR was then performed using the 7500 PCR System from Applied Biosystems on a 1/800 dilution of the cDNA with osterix and 18S primers (Applied Biosystems). PCR conditions were as follows: 50°C for 2 min, 95°C for 10 min and 95°C for 15 s, 62°C for 1 min for 32 cycles. Data were normalized to endogenous 18S rRNA levels using the Applied Biosystems ΔΔCt Method.
Filling and priming of osmotic pumps
Alzet (Cupertino, CA) micro-osmotic pumps (model 1007D) were used to deliver SDF-1 in athymic rats; the reservoir volume of this model is 90 μL and the pumping rate is 12 μL/day. Each Alzet pump was filled according to the manufacturer's protocol.
Immunohistochemistry and analysis
Five days after surgical implantation of collagen pellets containing PBS, BMP-2 (2.5 μg), or BMP-2 plus SDF-1 (0.5 μg), rats were euthanized and the subcutaneously implanted collagen discs were removed and analyzed by immunohistochemistry. The specimens were fixed for 24 h in 10% buffered neutral formalin. Each specimen was embedded in paraffin and cut into 5-μm-thick sections. The sections were then stained with hematoxylin and eosin or subjected to immunihistochemical staining procedures using the Vectastain® ABC Kit (Vector Laboratories, Burlingame, CA) with antibodies specific for CXCR4 (Abcam, Cambridge, United Kingdom) or CD18 (LifeSpan Biosciences, Seattle, WA) at a dilution of 1:100. For quantitative analysis, 0.25 mm2 regions of interest (ROI) were selected using digital software (Olympus, Tokyo, Japan). Four ROIs were selected at the edges of each implant and the number of CD18-positive cells were counted within the ROI.
Statistical analysis
Results are reported as the mean of determinations with error bars representing the standard error of the mean. Statistical significance was calculated with a one-way analysis of variance and subsequently with the Holm–Sidak post hoc test. When multiple comparisons were not made, statistical significance was calculated using the Student's t-test. Analysis was performed using Statistical Products for Social Sciences version 13.0 (SPSS 13.0) for Windows (SPSS, Chicago, IL). Statistical probability of p < 0.05 was considered significant.
Results
BMP-2 implantation on the chest of an athymic rat results in mobilization of MOPCs into the peripheral blood
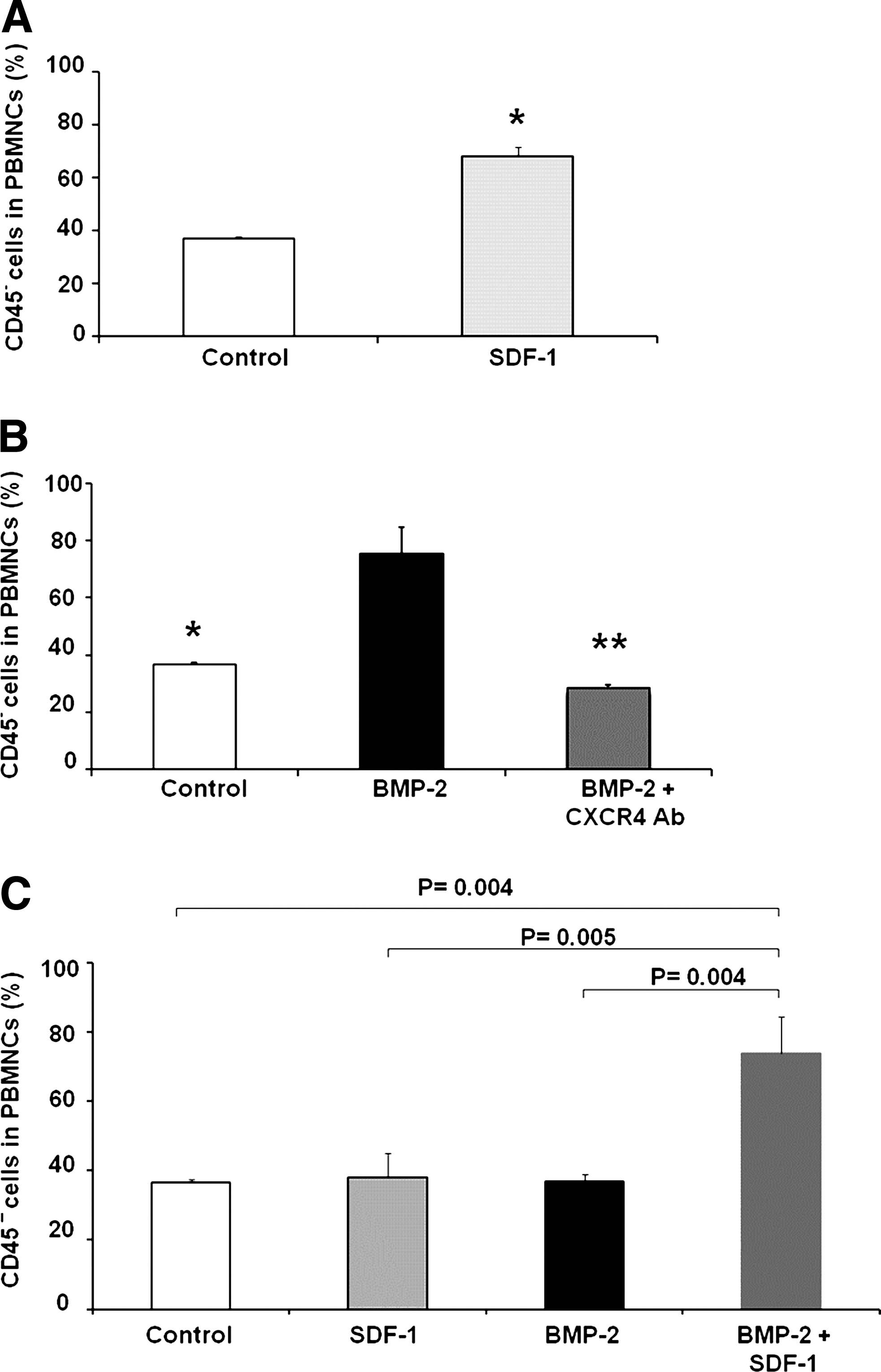
Flow cytometry demonstrated that the CD45− population increased in the peripheral blood after subcutaneous BMP-2 (5 μg) implantation in athymic rats (Fig. 1A). Implantation of a collagen pellet containing PBS caused a small increase in the circulating CD45− population as a response to the surgical injury, which was maximal on day 5. This increase in circulating CD45− cells is expected in response to surgical injury. 6 The addition of BMP-2 to the collagen pellet resulted in a statistically significant increase of CD45− cells as compared to the surgical injury alone at day 5 (p = 0.002) and a trend for an increase at day 3 (p = 0.09). The percentage of the CD45− population in the PBMNC fraction rose from 10% in animals implanted with pellets containing PBS to 83% in animals implanted with BMP-2-containing pellets (p < 0.001). The increase in the CD45− population peaked at day 5 and dropped significantly by day 7 to 19%.

BMP-2 implantation results in increased mobilization of MOPCs to the peripheral blood.
The CD45− cell population can be induced to the mature osteoblast phenotype
To demonstrate that the CD45− cell population isolated from PBMNCs contained osteoprogenitor cells, we cultured CD45− cells under osteogenic conditions. After 3 days, alkaline phosphatase mRNA levels were dramatically enhanced compared with cells cultured under nonosteogenic conditions (p < 0.001, Fig. 1B). After 21 days of culture under osteogenic conditions, the CD45− cell population produced mineralized nodules. Those cultured in the presence of BMP-2 produced 147 ± 0.9 nodules per six wells (n = 4). In multiple experiments these cells produced <5 nodules per six wells when cultured without BMP-2. This result confirms that the CD45− cell population from PBMNCs contains osteoprogenitor cells capable of responding to BMP-2 and producing mature osteoblasts with mineralized bone matrix.
SDF-1 induces mobilization of CD45− MOPCs
SDF-1 is thought to be involved with mobilization of CD34+ HSCs, but its role in mobilization of osteoprogenitor cells has not been studied. To demonstrate the role of SDF-1 in mobilization of MOPCs, we delivered 10 μg SDF-1 by gradual release subcutaneously either from a collagen pellet or from a subcutaneous mini-osmotic pump for 5 days. FACS analysis showed that the percentage of MOPCs in the PBMNC fraction on day 5 increased from 37% in rats implanted with either pellet or pump alone to 68% in rats that received SDF-1 on the pellet (p = 0.001, Fig. 2A) or 64% in rats that received SDF-1 via osmotic pumps (data not shown). This finding suggests that SDF-1 can increase mobilization of MOPCs into the circulation.
SDF-1 induces mobilization of MOPCs and potentiates their mobilization in response to BMP-2.
Antibody to CXCR4 inhibits mobilization of MOPCs
To determine whether SDF-1 was important in mobilization of MOPCs from the bone marrow to the peripheral blood, we implanted athymic rats with a collagen disc containing either 5 μg BMP-2 alone or combined with antibody to the SDF-1 receptor, CXCR4 (0.2 mg). PBMNCs were harvested after 5 days for measuring CD45− MOPCs by FACS analysis. CXCR4 antibody inhibited mobilization of CD45− cells from the bone marrow that in response to local tissue injury and BMP-2. The animals that received CXCR4 antibody and BMP-2 had fewer circulating CD45− PBMNCs (28%) compared to the animals that received BMP-2 alone (75%) (p < 0.001, Fig. 2B). This result suggests that SDF-1, in addition to its role as a homing signal, may be a key signal for the mobilization of MOPCs in peripheral blood.
SDF-1 potentiates BMP-2 mobilization of MOPCs in vivo
We have shown that injury plus BMP-2 at an optimal dose for bone formation (5 μg) mobilizes about 81% of the CD45− cell population by day 5 and that injury plus 10 μg of SDF-1 mobilizes about 68% of the CD45− cell population in athymic rats at day 5. In this study we examined the possible SDF-1 potentiation of BMP-2-induced mobilization of MOPCs by using suboptimal doses of each agent. A low dose of SDF-1 (0.5 μg) enhanced mobilization of CD45− MOPCs by BMP-2 (2.5 μg) on day 5 from 37% with BMP-2 alone to 74% in the combined presence of BMP-2 and SDF-1(p = 0.004, Fig. 2C). Although SDF-1 mobilized cells have not been reported to have osteogenic potential, our findings led us to hypothesize that SDF-1 may be a key signal for mobilization of MOPCs from the bone marrow. These data establish a novel role of SDF-1 in the BMP-2 induced mobilization of MOPCs.
CD45− MOPCs migrate in response to SDF-1 and BMP-2 in vitro
To establish that the MOPCs migrate in response to chemotactic agents, the CD45− population of cells was seeded on transwell filters and incubated with SDF-1 or BMP-2 in the lower chamber of the plates. Approximately three times more cells migrated from the transwell filter toward either SDF-1 (1000 ng/mL) or BMP-2 (100 ng/mL) compared with the number of cells that migrated toward medium alone controls (p < 0.001, Fig. 3). This in vitro assay suggests that the CD45− cell population may not only mobilize in response to these agents in vivo, but also migrate (home) toward areas where SDF-1 or BMP-2 is concentrated.
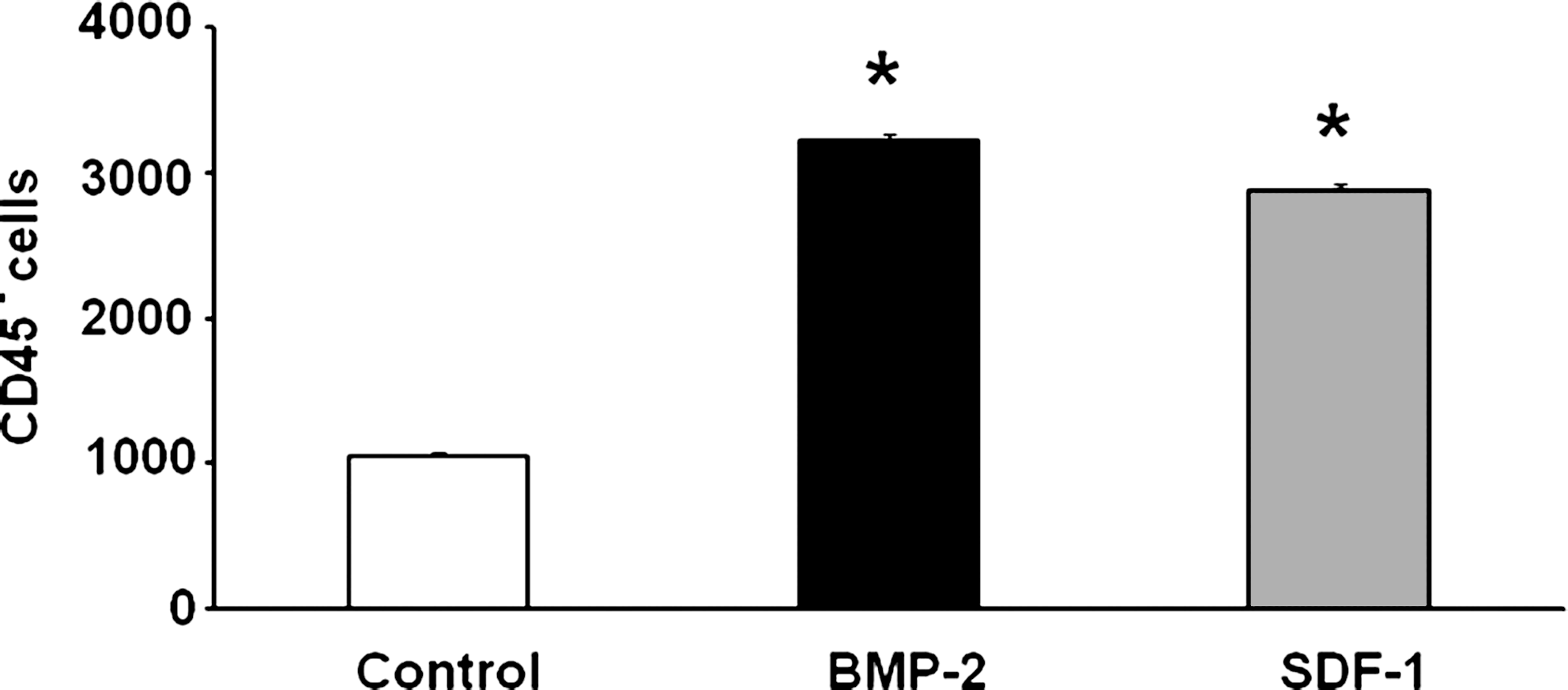
CD45− MOPCs migrate in response to BMP-2 and SDF-1 in vitro. CD45− cells were harvested from PBMNCs of rat after implantation of BMP-2 and plated in the upper chamber of transwell plates whose lower chamber contained either BMP-2 or SDF-1. The migration of cells from the upper chamber to the lower chamber was compared to the migration of cells toward PBS (control). Approximately three times more cells migrated toward BMP-2 or SDF-1 as migrated toward PBS. *p < 0.001 compared to control.
SDF-1 enhances homing of MOPCs in response to BMP-2 in vivo
To demonstrate that local application of SDF-1 synergizes with BMP-2 to increase homing of MOPCs to the collagen disc, immunohistochemistry was performed to detect cells expressing CD18. CD18-positive cells play a critical role in osteogenesis of the bone marrow stromal stem cells. 15 We also confirmed the presence of the SDF-1 receptor, CXCR4, a progenitor cell surface marker. Representative sections from each group are shown in Figure 4. Homing of MOPCs in response to BMP-2 was confirmed in these samples as more CXCR4 and CD18-positive cells were observed in the implants containing BMP-2 than in control implants. In addition, more CD18 or CXCR4-positive cells were observed in implants containing BMP-2 plus SDF-1 (Fig 4A). Results obtained from counting CD18-positive cells in four sections from each of four implants/group are shown in Figure 4B. There was a trend for the BMP-2 + SDF-1 group to increase CD18-positive cells compared to BMP-2 alone (p = 0.06). Thus, SDF-1 can enhance cellular homing to BMP-2.
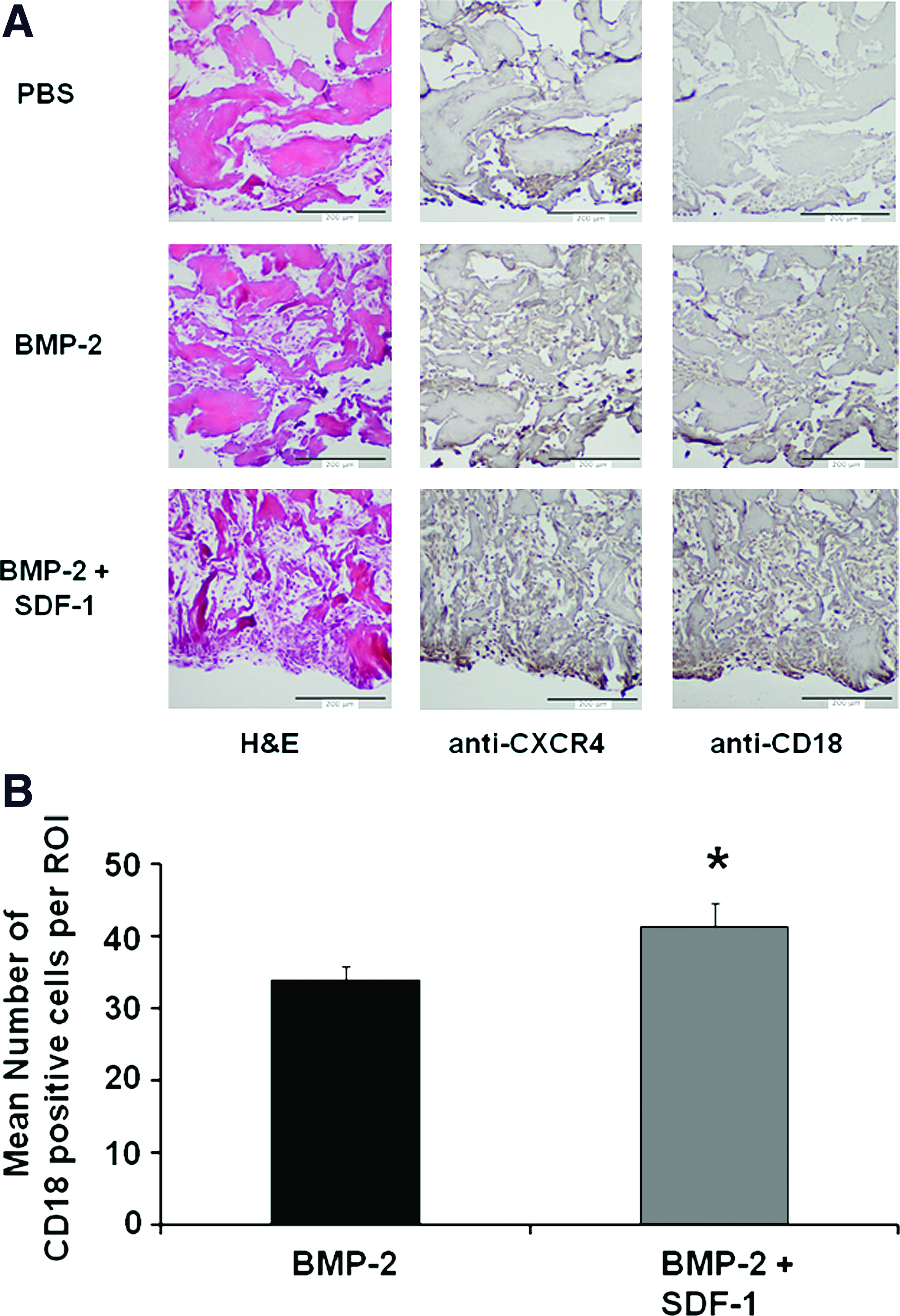
SDF-1 enhances the homing of MOPCs to sites containing BMP-2. Collagen pellets containing PBS, BMP-2 (2.5 μg), or BMP-2 plus SDF-1 (0.5 μg) were harvested 5 days after implantation, processed for microscopy, and representative samples stained with H&E (left), anti-CXCR4 (middle), or anti-CD18 (right).
A suboptimal dose of BMP-2 fails to make consistent bone in the athymic rat model of ectopic bone formation
To establish a dose–response relationship and determine a minimum threshold dose of rhBMP-2 in the athymic rat ectopic bone formation model, we implanted collagen pellets with 2.5–40 μg BMP-2. Faxitron X-rays of explants 28 days after implantation demonstrate decreasing bone formation as the dose of BMP-2 decreases (Fig. 5A). When 2.5 μg BMP-2 was used, the results became inconsistent and some explants failed to make any bone. This suboptimal dose of BMP-2 was selected for subsequent experiments investigating the ability of SDF-1 to enhance the efficacy of BMP-2-induction of bone formation.
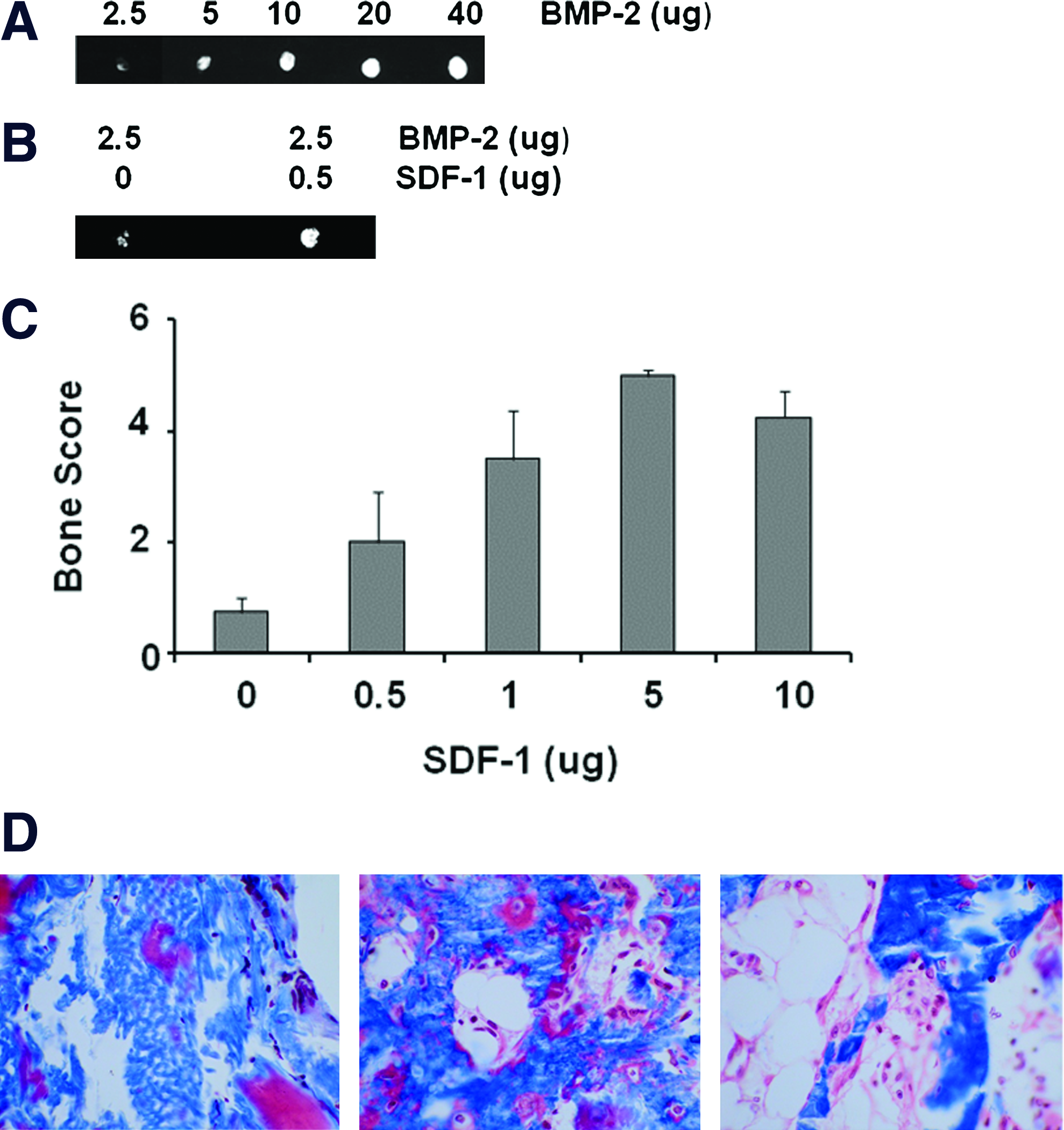
SDF-1 potentiates BMP-2 responsiveness in vivo
We placed a suboptimal dose of rhBMP-2 (2.5 μg) plus various doses of the chemokine, SDF-1 (0–10 μg), on a collagen pellet and implanted it subcutaneously on the chest of athymic rats. Pellets containing the intentionally low dose of BMP-2 contained some bone, but were not mineralized throughout the pellet; the addition of SDF-1 produced an increase in the amount of bone induced by BMP-2 (Fig. 5B). This demonstrates that SDF-1 enhances the efficacy of BMP-2 to induce bone formation in vivo. The amount of bone in each X-ray was scored and the scores were pooled to calculate the mean bone score for each group (Fig. 5C). There was a dose-dependent effect of adding SDF-1 to BMP-2 (analysis of variance, F = 7.88, p = 0.001) and 5 μg SDF-1maximally enhanced BMP-2 induced ectopic bone formation. Gomori's Trichrome staining of sections taken from implants treated with 2.5 μg BMP-2 without SDF-1 (Fig. 5D, left) revealed little or no bone with residual collagen bundles still evident. Sections taken from implants with BMP-2 plus 5 mg SDF-1 revealed (Fig. 5D, center) new woven bone with active surfaces of osteoid (color) and (Fig. 5D, right) mature bone with formation of a marrow cavity, as is often seen with higher doses of BMP-2. This confirms our X-ray observations that SDF-1 enhances induction of bone formation by a suboptimal dose of BMP-2.
SDF-1 enhances BMP-2-induced differentiation of the osteoblast phenotype in pleuripotent C2C12 cells
A recent study reported that SDF-1 was required for BMP-2-induced differentiation of the osteoblast phenotype in C2C12 cells. 16 We extended that observation and show in C2C12 cells that 50 ng/mL SDF-1 enhanced BMP-2 (25 ng/mL)-induced mRNA levels of osterix (an early marker of the osteoblast phenotype) 2.8-fold on day 1(p = 0.009) (Fig 6).
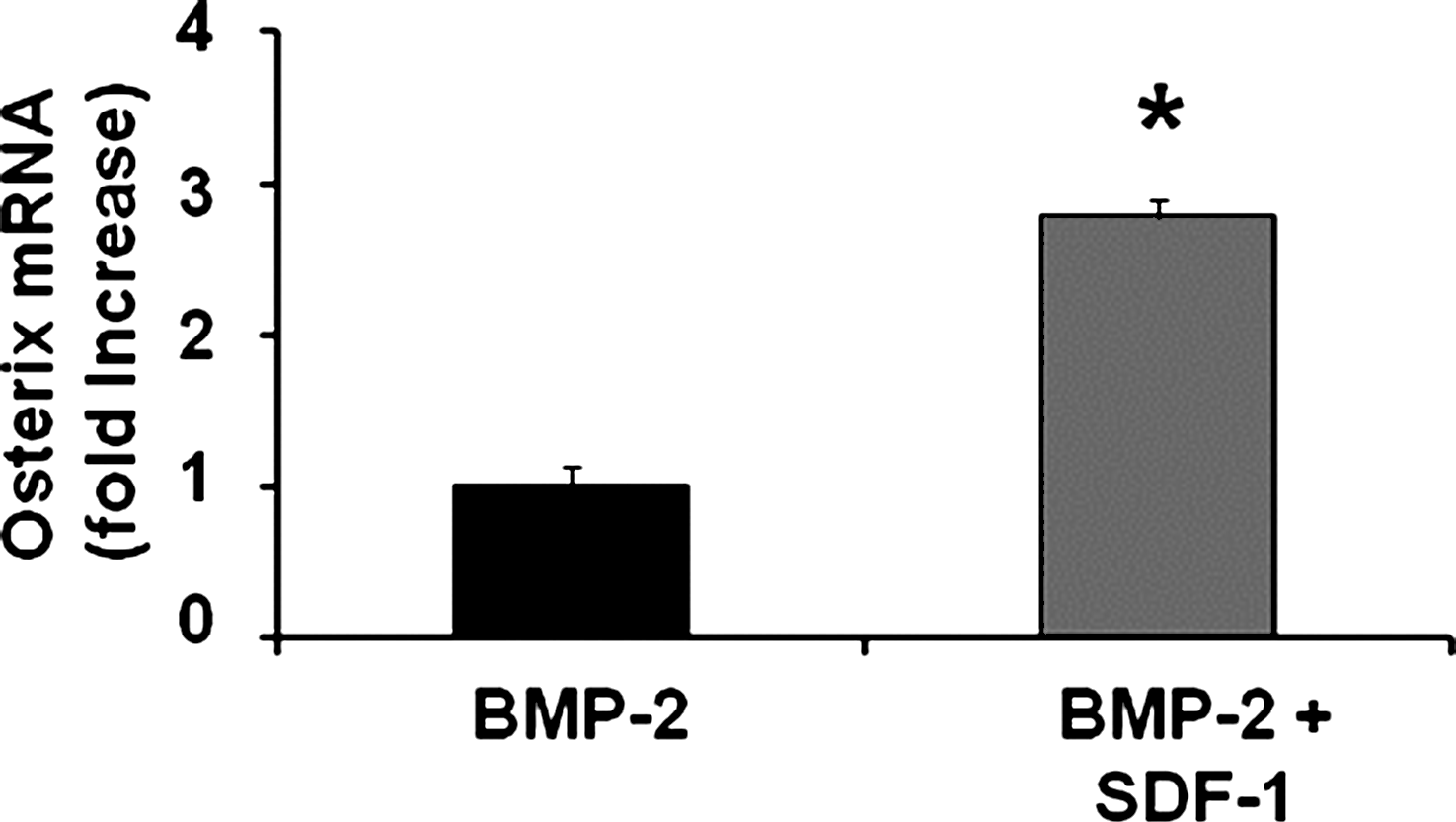
SDF-1 enhances BMP-2-induced differentiation of pleuripotent C2C12 cells to the osteoblast phenotype. C2C12 cells were treated with 25 ng/mL BMP-2 ± 50 ng/mL SDF-1. The level of osterix mRNA was measured on day 1. The level of BMP-2-induced osterix was increased by SDF-1 treatment. *p = 0.009 compared to BMP-2.
Discussion
Although BMP-2 is approved by the U.S. Food and Drug Administration (FDA) for clinical application as a bone graft substitute in interbody spine fusions and for healing open tibial fractures, the need for large doses of expensive recombinant protein to achieve therapeutic goals makes its use prohibitive for many patients. One approach to improving the efficiency of BMP-2 to induce bone formation has been to implant exogenous bone marrow or culture-expanded MSCs concurrently with BMP-2 to provide immediately available BMP-2 responsive cells rather than wait for chemotaxis of osteoprogenitor cells in response to BMP-2. However, neither approach has rendered adequate results in primates.17–19 Herein we report the novel approach of increasing the mobilization and homing of endogenous MOPCs to the site of BMP-2-induced bone formation. The CD45− cells isolated from peripheral blood in these studies have been differentiated in culture to express the osteoblast phenotype and produce mineralized nodules, proving that they are truly osteoprogenitors.
MSCs are capable of local regeneration of skeletal tissues, including bone. Progenitor cells exist in the bone marrow and in peripheral tissues. Although initially difficult to identify, there is now strong evidence that circulating osteoblast-lineage progenitor cells exist in humans.5,20 Research in this field has been hampered by the lack of a single reliable surface antigen marker for skeletal progenitor cells and because surface marker expression can change in vitro. 21 There is now some agreement that osteoblast progenitor cells are CD45−, CD34−, CD44+, Sca-1+, lin−, and CD90+.20,22 A recent murine study found that the CD45− fraction of mononuclear cells in bone marrow was mobilized within 7 days to the site of BMP-2-induced bone formation. 7 Our observations reported herein have confirmed the murine observation in rats and have used the chemokine, SDF-1, to enhance the mobilization of these cells and their homing to the site of BMP-2 induced bone formation. We have also shown that the CD45− cells mobilized into peripheral blood migrate in vitro in response to both BMP-2 and SDF-1 and confirmed that the mobilized MOPCs are capable of differentiating into osteoblastic cells.
The role of SDF-1 has been most focused on cell homing and maintenance of cellular niches, but SDF-1 has also been linked to mobilization of nonosteogenic progenitor cells from bone marrow.10,11 In the present investigation we have clearly demonstrated a role for SDF-1 as a signal to trigger mobilization of osteoprogenitor cells from the bone marrow. Although we did not investigate whether other signals were involved with MOPC mobilization, we did demonstrate the requirement of SDF-1 for mobilization, by demonstrating that antibody to CXCR4, the SDF receptor, significantly reduced BMP-2-induced mobilization of MOPCs.
A recent study reported that SDF-1 was required for BMP-2-induced differentiation in pleuripotent C2C12 cells. 16 This led us to hypothesize that SDF-1 synergizes with BMP-2 to enhance osteoblast differentiation both in vitro and in vivo. We demonstrated in this study that treatment of pleuripotent C2C12 cells with SDF-1 enhanced BMP-2-induced mRNA levels of the osteoblastic marker, osterix.
Taken together, these findings raise the possibility that, in vivo, SDF-1 could contribute to enhancing the efficiency bone BMP-2-induced bone induction via three mechanisms: mobilization of MOPCs, homing of MOPCs to the bone formation site, and enhancement of differentiation of the osteoprogenitor cells. Our in vivo experiments demonstrated SDF-1-stimulated enhancement of BMP-2-induced mobilization, homing, and osteoprogenitor differentiation to increased ectopic bone formation. In these studies, placement of a single dose of SDF-1 dose dependently enhanced bone formation on a collagen disc containing a deliberately suboptimal dose of BMP-2.
Our study has several limitations. With the current design, the relative contribution of each SDF-1 effect (increased mobilization, homing, and differentiation of osteoprogenitor cells) was not able to be assessed. In addition, we do not know how the release profile of SDF-1 from the collagen discs compares with the release rate of BMP-2 that has a residence half-life of 3.5 days at the site. 23 Since a plain collagen delivery matrix, while optimal for small rodents, resorbs too quickly in primates, a detailed study of SDF-1 release kinetics in this model would be of very limited value and was not performed. The importance of our results might also be limited if any/all of these effects observed in rodents do not translate to primates that may have different thresholds for progenitor cell mobilization, homing, and impact on differentiation.
Notwithstanding these limitations, our findings strongly suggest that addition of SDF-1 to BMP-2 may enhance the efficiency of BMP-2 and, if translatable to primates, could make recombinant BMPs more clinically accessible to patients. To our knowledge, this is the first demonstration of enhanced efficiency of BMP-2 to induce bone formation by chemokine-stimulated mobilization, homing, and differentiation of osteoprogenitor cells. Future studies will apply SDF-1 with BMP-2 to induce bone formation in a more challenging model such as spine fusion.
Footnotes
Acknowledgments
The authors are grateful to Colleen Oliver, D.V.M., for her help and support in animal surgery and take downs. The authors would like to thank Dr. Sreedhara Sangadala for critical discussions and for performing statistical analyses, Mesfin Tekelmarian, M.S., for his technical support, and Ludmilla Friedman for support with histology and immunohistochemistry. This work was supported by a VA merit award to Dr. Boden. Hui Liu is partially supported by the International Program of Project 985, Sun Yat-sen University.
Disclosure Statement
Dr. Boden has received compensation in the past as a consultant for Medtronic Sofamor Danek and for intellectual property. The terms of this arrangement were reviewed and approved by Emory University in accordance with its conflict of interest policies. The other authors have no conflict of interest.
This work was performed at the Atlanta Veterans Affairs Medical Center.
This work was presented in part at the 31st annual meeting of the American Society for Bone and Mineral Research in 2009.