
Abstract
Previous studies have reported that well-defined culture conditions can improve keratinocytes terminal differentiation and reproducibility. The aim of our study was to compare skin substitutes cultured in a complete medium with those cultured in a serum-free medium at the air–liquid interface to optimize the self-assembly method. Skin substitutes, cultured in a serum-free medium over 7, 14, and 21 days, were compared with others cultured in a complete medium (5% serum) over the complete culture period. Masson's Trichrome staining showed that the substitutes cultured in a serum-free medium generated a well-developed and differentiated epidermis. Immunolabeling analyses between the substitutes cultured without serum and those cultured in complete serum showed similar expression of epidermal differentiation markers, dermo-epidermal junction, and dermal extracellular matrix components. On the basis of our Attenuated Total Reflectance-Fourier Transform Infrared (ATR-FTIR) results, the skin substitutes cultured in serum-free condition over 21 days of culture at the air–liquid interface showed lower frequencies of the CH2 symmetric mode of vibrations, which means a better lipid organization of the stratum corneum. No significant difference in hydrocortisone penetration was observed between serum-free medium substitutes and the controls. Results demonstrate that the absence of serum does not compromise the characteristics of the skin substitutes observed in this study.
Introduction
In most tissue-engineering studies, the medium provides the essential nutrients such as amino acids, fatty acids, sugars, ions, trace elements, vitamins, cofactors, and molecules necessary to maintain the proper chemical environment for the cells to proliferate accordingly. 7 To produce skin substitutes, the addition of animal or human serum is required in the culture medium because serum contains various growth factors, cytokines, hormones, vitamins, and proteins that increase cell growth and cell proliferation.8,9
Serum is an undefined mixture composed of over 1000 different components, 10 some of whose functions are known, whereas others are not. The primary functions of serum in cell culture are to stimulate cell growth and proliferation by hormonal factors; to carry hormones, minerals, trace elements, and lipids by transport proteins; and to allow the attachment and spreading of cells by the formation of extracellular matrix components.8,11 Despite the fact that serum is essential for cell proliferation, it also has many non-negligible disadvantages. First, serum is an ill-defined medium supplement and its composition is inconsistent from one lot to the next, which can increase results variability. 12 If the serum sample is consistent across the experiment, this should improve the reproducibility. 13 Second, serum production is costly and unefficient. Effectively, about 44–144 cows must be slaughtered to obtain 1 L of fetal bovine serum. 14 Third, fetal calf serum can contain different amounts of endotoxins, hemoglobin, and infectious agents (fungi, bacteria, viruses, and prions).11,15 Finally, high levels of proteins are present in the serum, which can lead to the overgrowth of some unwanted cell types and downstream protein purification. 16
Previous studies have shown that small changes in the concentration of hormones 17 and cholesterol, 18 or large changes in the concentration of glutamate, 19 which is present in the serum, affect cell culture. Taking into account the results obtained by these groups and others, the present study aimed to test if a partial or total reduction of serum, at strategic moments over the course of keratinocytes culture, can improve reproducibility and avoid interference from ill-defined supplements. To optimize our self-assembly method, experiments were performed in which skin substitutes were cultured at the air–liquid interface, either in a complete medium or in a serum-free medium.
Materials and Methods
Patients
The subjects were Caucasian females aged between 18 and 57 years (cell populations: ♀18a, ♀18b, ♀26, ♀29, and ♀57). Skin samples used to generate primary human cell banks were derived from breast reduction surgeries. This study was conducted in agreement with the Helsinki declaration and performed under the guidelines of the research ethics committee of the “Centre hospitalier affilié universitaire de Québec.” All patients were given adequate information to provide written consent.
Cell culture media
Fibroblasts were cultured in the high-glucose Dulbecco-Vogt modification of Eagle's medium (DMEM) supplemented with 10% fetal calf serum (Invitrogen, Burlington, Ontario, Canada), 100 IU/mL penicillin G (Sigma, Oakville, Ontario, Canada), and 25 μg/mL gentamicin (Schering, Pointe-Claire, Québec, Canada). Keratinocytes were cultured in a combination of high-glucose DMEM with Ham's F12 (3:1) supplemented with 5% Fetal Clone II serum (Hyclone, Scarborough, Ontario, Canada), 5 μg/mL insulin (Sigma, Oakville, Canada), 0.4 μg/mL hydrocortisone (Calbiochem, EMD Biosciences, Gibbstown, NJ), 10−10 M cholera toxin (MP Biomedicals, Montréal, Québec, Canada), 10 ng/mL human epidermal growth factor (Austral Biological, San Ramon, CA), 100 IU/mL penicillin G (Sigma), and 25 μg/mL gentamicin (Schering).
Cell culture
Keratinocytes were extracted using the isolation method with thermolysin and trypsin. 20 According to this method, keratinocytes (passage 1) were seeded at 8 × 103 cells/cm2 on a feeder layer of irradiated 3T3 mouse fibroblasts. Fibroblasts were extracted using the isolation method with thermolysin and collagenase.21,22 They were seeded at 4 × 103 cells/cm2 and used at passage 6 for skin substitute's production. All cultures were incubated at 37°C in an 8% CO2 air atmosphere and changed three times a week with the media previously described in the cell culture media section. Cells were frozen at −150°C until needed.
Production of tissue-engineered substitutes
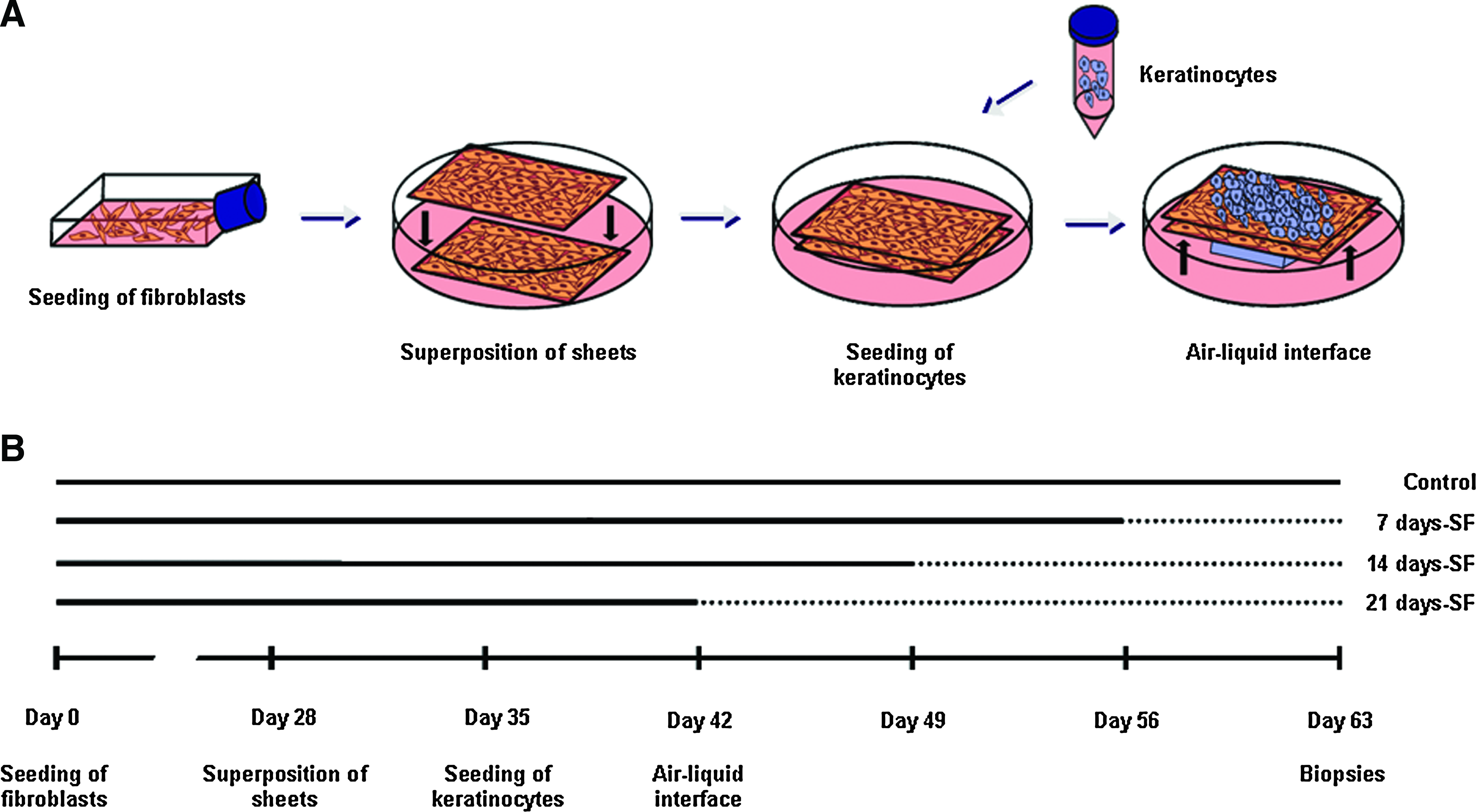
Skin substitutes were produced using the self-assembly method, as previously described. 6 Briefly, the fibroblasts were cultured in the presence of ascorbic acid at a concentration of 50 μg/mL (Sigma, Mississauga, Ontario, Canada) to form manipulatable sheets. After 28 days, dermal sheets were carefully detached of flasks with curved forceps. Then, dermal sheets were superimposed and a Merocel sponge was placed on the top of the substitutes to facilitate the natural fusion of dermal sheets leading to the formation of a new dermal layer. 23 After a week, the keratinocytes (passage 2) were seeded on the dermal equivalent to form a new epidermal layer. After 7 days of growth, the substitutes were raised to the air–liquid interface (Fig. 1A). Starvation of serum was begun at day 0 of the air–liquid interface culture before the beginning of the differentiation process. 24 Substitutes were cultured in the serum-free medium (DMEM with Ham's F12) over 7, 14, or 21 days and compared with the controls, which were cultured in the presence of serum for the complete experiment (Fig. 1B). After 21 days of culture at the air–liquid interface, biopsies of the skin substitutes were analyzed.
Histological and morphometric analysis
Two biopsies of each combination were fixed in HistoChoice® solution and embedded in paraffin wax. Five-micrometer-thick sections were cut and stained with Masson's Trichrome. The thickness of the epidermis was measured with MetaVue Imaging System software version 7.1.2.0 (Molecular Devices, Sunnyvale, CA). Ten measurements in four different sections of each biopsy were performed (n = 80 for each combination).
Immunolabeling analyses
After 21 days of culture at the air–liquid interface, biopsies were taken and the samples were embedded in Tissue-Tek O.C.T. Compound (Somagen Diagnostics Inc., Edmonton, Alberta, Canada) and stored at −80°C until needed. Indirect immunofluorescence analyses were performed on acetone-fixed cryosections (5 μm thick). Frozen sections of human skin were used as positive controls. Immunolabeling studies were performed with monoclonal and polyclonal antibodies. The primary antibodies were: rabbit anti-involucrin (dilution 1:200; Sigma, Oakville, Ontario, Canada), mouse monoclonal anti-keratinocyte transglutaminase (dilution 1:50; BTI, Stoughton, MA), rabbit polyclonal anti-mouse loricrin (dilution 1:750; Babco [Covance], Berkeley, CA), mouse monoclonal (IgG1) anti-human filaggrin (dilution 1:800; BTI), rabbit anti-keratin 5 (dilution 1:200; Babco [Covance]), mouse anti-keratin 10 (dilution 1:50; Cederlane, Burlington, Ontario, Canada), guinea-pig anti-keratin 6 (dilution 1:1000), 25 mouse anti-keratin 16 (dilution 1:50; ICN Biomedicals Inc., Aurora, OH), mouse monoclonal (IgG1) anti-laminin V (γ2-chain) (dilution 1:200; Chemicon, Temecula, CA), rabbit anti-collagen IV (dilution 1:50; Abcam, Cambridge, MA), mouse anti-collagen VII (dilution 1:400; Chemicon), mouse anti-collagen I (dilution 1:200; Calbiochem, Gibbstown, NJ), rabbit anti-collagen III (dilution 1:200; Cederlane), rabbit anti-elastin (dilution 1:200; Institut Pasteur, Lyon, France) and mouse anti-fibronectin (dilution 1:25; ATCC, Rockville, MD). The second antibody was goat anti-rabbit IgG (H + L) Alexa 594 (dilution 1:400; Molecular Probes, Eugene, OR) or goat anti-mouse IgG (H + L) Alexa 594 (dilution 1:800; Molecular Probes), goat anti-rabbit Alexa 594 (dilution 1:400; Molecular Probes). The nuclei were labeled with Hoechst reagent 33258 (dilution 1:100; Sigma), added in the secondary antibody solution.
ATR-FTIR experiments
Infrared spectra of the stratum corneum of the skin substitutes were obtained using a Golden Gate single reflection-attenuated total reflection system (Specac, Pleasantville, NY) fitted with a diamond crystal. The infrared spectra were recorded with a Nicolet Magna 860 Fourier transform spectrometer (Thermo-Nicolet, Madison, WI) equipped with a narrow band mercury–cadmium–telluride detector and a germanium-coated KBr beam splitter. At each temperature, a total of 128 interferograms were acquired, coadded, and Fourier transformed, using a Happ–Genzel apodization function to give a spectral resolution of 4 cm−1 in the spectral range of 4000–750 cm−1. All data were processed with the Grams 386 software (Galactic Industries Corporation, Salem, MA). The spectral region corresponding to the CH2 stretching band vibrations was baseline-corrected using a cubic function and the frequency peak position was determined using the center of gravity at the top 10% of the bands.
Percutaneous absorption
Percutaneous absorption was measured by the standard Franz diffusion cell technique as described by Franz.26–28 Briefly, samples were clamped tightly between the two glass chambers of the diffusion cell (FDC-100 Standard, 0.63 cm2 surface area O-ring; Crown Glass, Somerville, NJ). Receiver compartment was filled with media (DMEM supplemented with penicillin G [Sigma], gentamicin [Schering], and 0.5 μg/mL of fungizone [Bristol-Myers Squibb Canada, Montréal, Québec, Canada]) and maintained at 37°C with a water jacket. One hundred microliters of 3 H-hydrocortisone (1 mCi/100 μL/chamber; NEN-Dupont, Mississauga, Ontario, Canada) was deposited on the substitute. Samples were taken at selected intervals (1, 2, 4, 6, 8, and 24 h) with a 5 mL syringe prolonged by a catheter (3½ FR Tom Cat Length 4½) and were conserved at 4°C until needed. Radioactivity was determined with a scintillation counter Beckman LS 6000 SC (Beckman Instruments Inc., Fullerton, CA) by adding 0.5 mL of sample to 4.5 mL of scintillation fluid Scintisafe™ 30% (Fischer Scientific Ltd., Québec, Québec, Canada).
Statistical analysis
Data were analyzed using the analysis of variance test and expressed as means ± standard deviation. A p-value <0.05 was selected to confer statistical significance. Percutaneous absorption results were analyzed with the PROC MIXED program of the SAS 9.1 (SAS Institute, Cary, NC).
Results
Macroscopic aspect of skin substitutes
Here, it is interesting to note the presence of a complete proliferation of the keratinocytes with a seamless epidermal layer. This wide surface can be seen in all conditions, suggesting, in a macroscopical way, that proliferation is not affected by serum-free culture. Further, substitutes cultured in the serum-free medium for 21 days (Fig. 2D) showed a uniform and smooth surface as observed with the control substitutes (Fig. 2A). The same macroscopic aspect was observed after serum had been removed over 7 (Fig. 2B) and 14 days (Fig. 2C) before the completion of the air–liquid culture period.
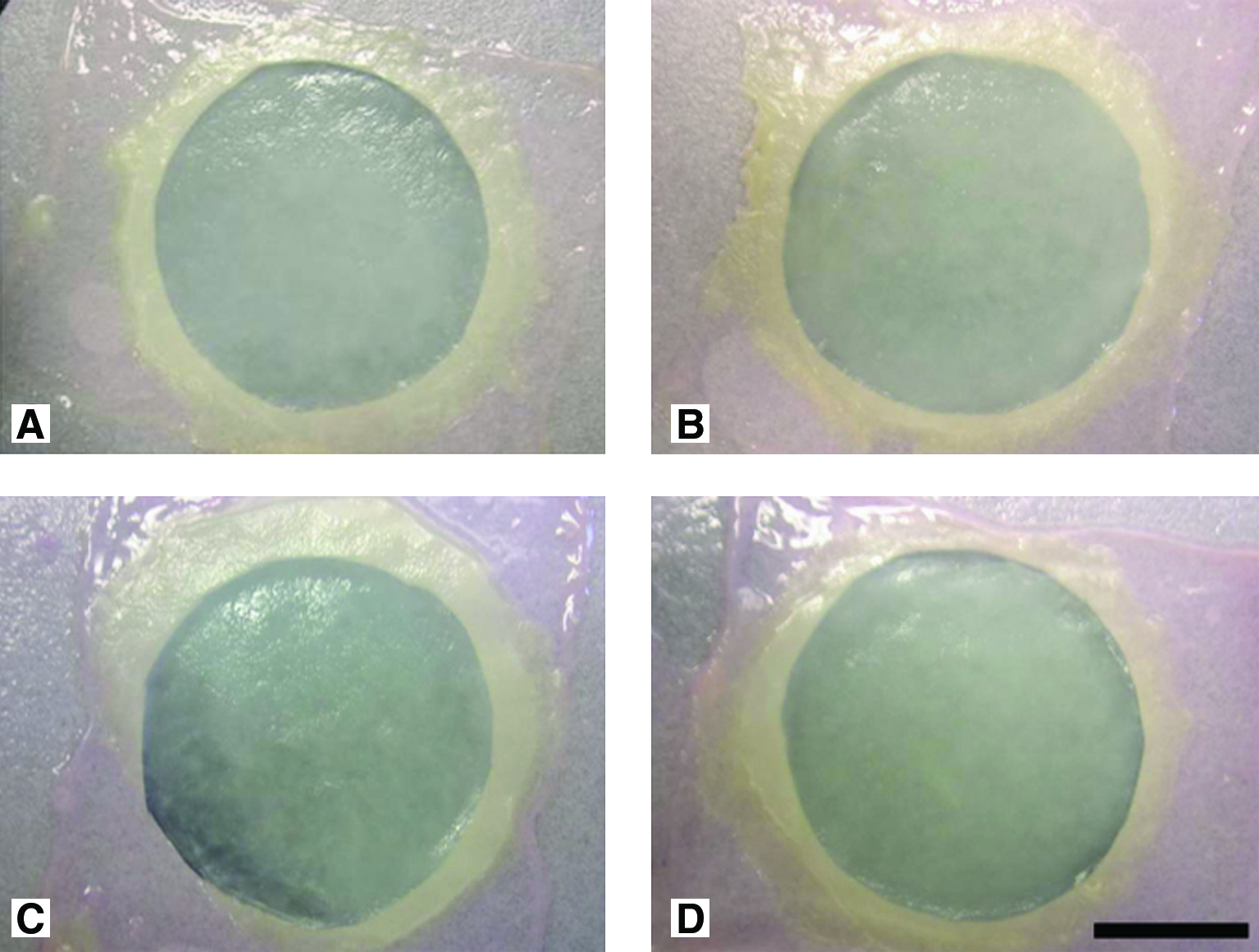
Macroscopic aspects of the substitutes cultured in
Histological aspect
Masson's Trichrome staining revealed that substitutes cultured in a serum-free medium over 21 days (Fig. 3D) showed a well-developed and differentiated epidermis such as observed in control substitutes (Fig. 3A). A differentiation pattern similar to the controls was also observed for the other conditions (Fig. 3B, C). In the dermis, despite the absence of serum at the air–liquid interface, fibroblasts were still present and alive (Fig. 3B–D).
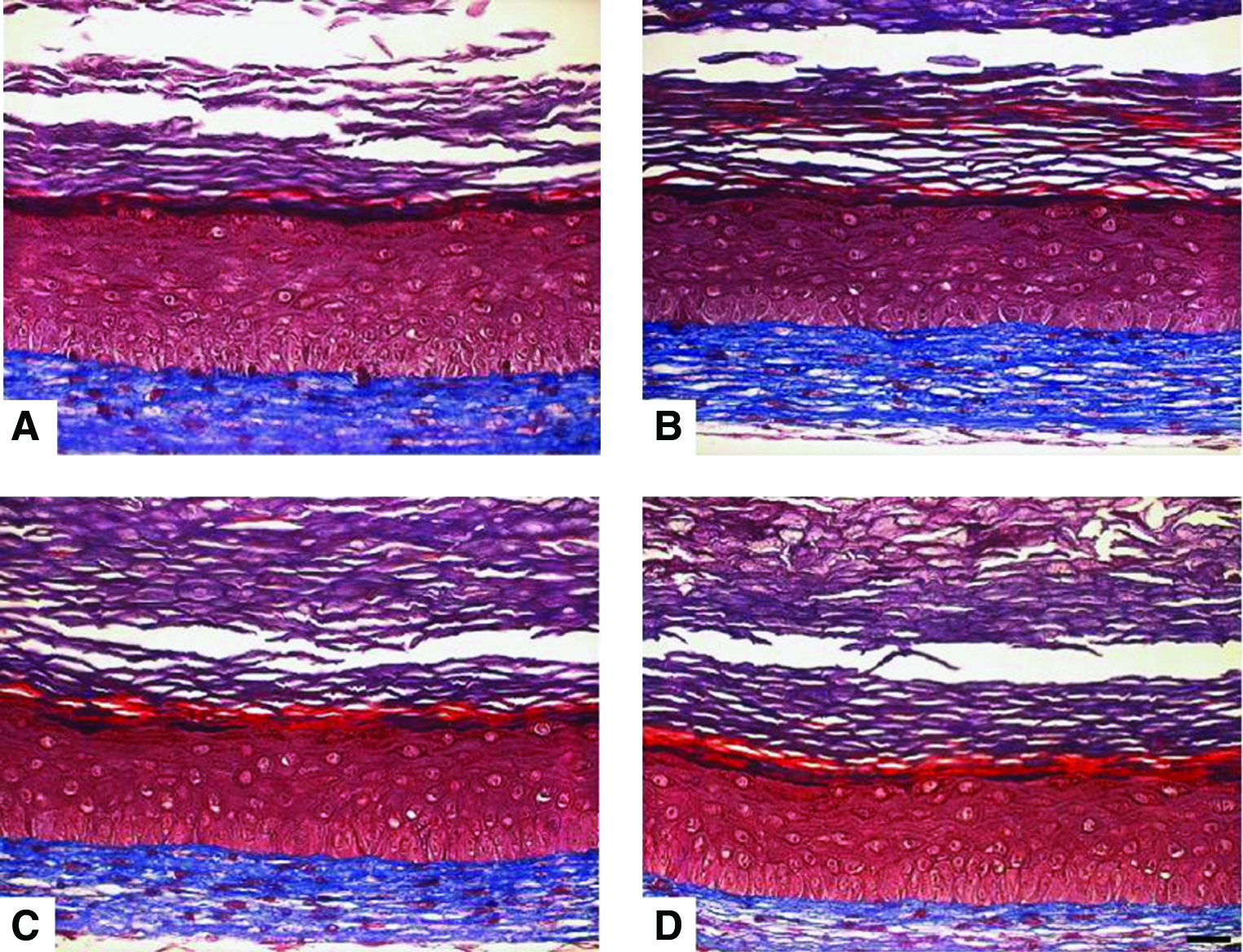
Masson's trichrome staining obtained from substitutes cultured in
Epidermal thickness
Measurements of the epidermis were done with the MetaVue Imaging software. No significant difference was observed in epidermal thickness between substitutes cultured with or without serum: 7 days-SF (231.3 ± 52.5 μm), 14 days-SF (312.3 ± 32.3 μm), and 21 days-SF (274.6 ± 27.1 μm), compared to the controls (246.4 ± 46.8 μm) (Fig. 4). The same observation was made for all cell populations (n = 5, where two substitutes were observed for each cell population).
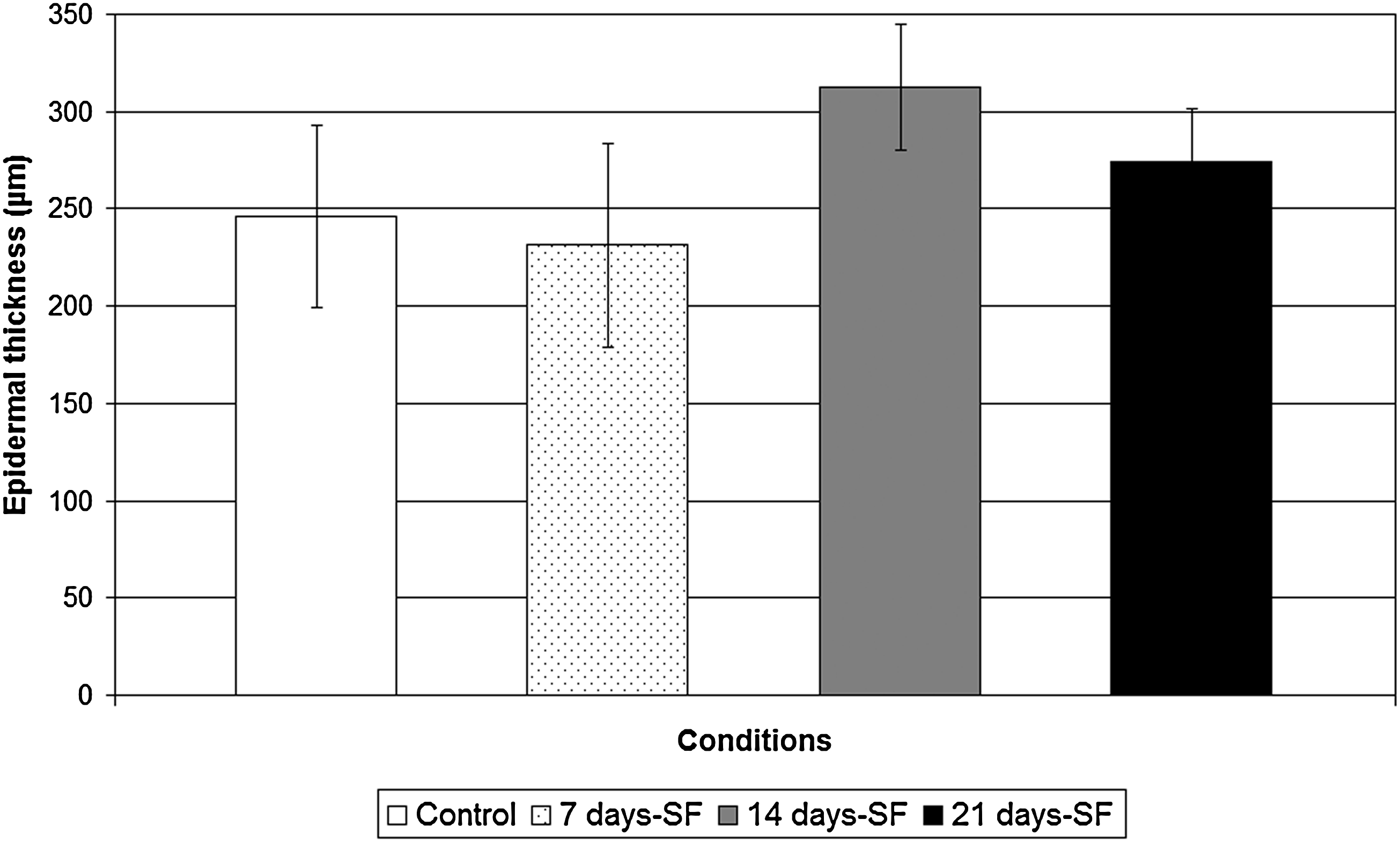
Epidermal thickness of substitutes cultured in 5% serum for the complete experiment (controls; white bar), serum-free medium for the last 7 days of the experiment (dotted bar), serum-free medium for the last 14 days of the experiment (gray bar), and serum-free medium for the last 21 days of the experiment (black bar). Statistical significance was determined using analysis of variance test (p < 0.05, n = 80). All data presented are the means ± standard deviation of one cell population. The results were confirmed for the five cell populations tested.
Immunolabeling analyses
In both normal human skin (NHS) and experimental controls, expression of involucrin and transglutaminase appeared in the spinous layer of the epidermis, whereas the filaggrin and loricrin showed a staining in the granular layer. The same pattern was obtained with our skin substitutes cultured in a serum-free medium over the complete period of culture at the air–liquid interface (21 days-SF) (Fig. 5A, B). Moreover, in NHS and in control substitutes, expression of basal keratins 5 (Fig. 6A) and 14 (data not shown) appeared in the basal layer, whereas the suprabasal keratin 10 was expressed in the suprabasal layers (Fig. 6A). Same observation was noted with the serum-free substitutes. Hyperproliferative keratins 6 and 16 were expressed in the suprabasal layers of control and serum-free substitutes in a similar way, but no labeling was observed in the NHS (Fig. 6B). Markers of the dermo-epidermal junction, such as laminin V, collagen type IV, and human collagen type VII, showed a staining, which appeared as a thin and continuous line in both NHS and our controls (Fig. 7). Substitutes cultured in a serum-free medium over the complete culture period at the air–liquid interface (21 days-SF) showed a similar appearance after these same stainings. No disruption in the dermo-epidermal junction was observed even if serum was removed during this long period. In serum-free medium conditions, collagen I and III, elastin, and fibronectin were expressed in the dermis of the skin substitutes as observed in NHS and in our controls (Fig. 8). However, expression of fibronectin seemed a little underexpressed in substitutes cultured without serum over 21 days (21 days-SF). For all differentiation markers tested, similar results were obtained with the two other conditions: 7 and 14 days-SF (data not shown).
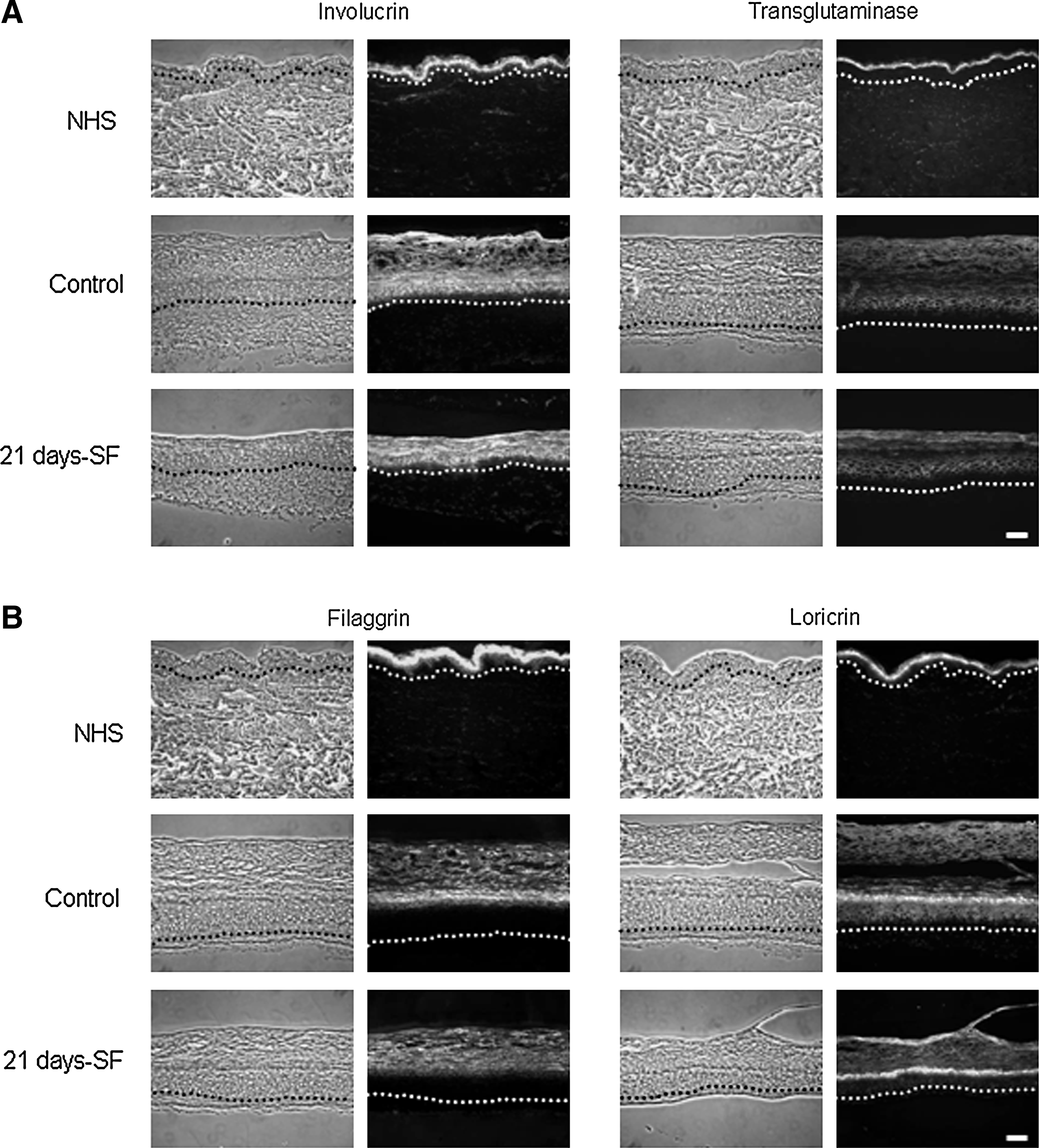
Immunolabeling of differentiation markers:
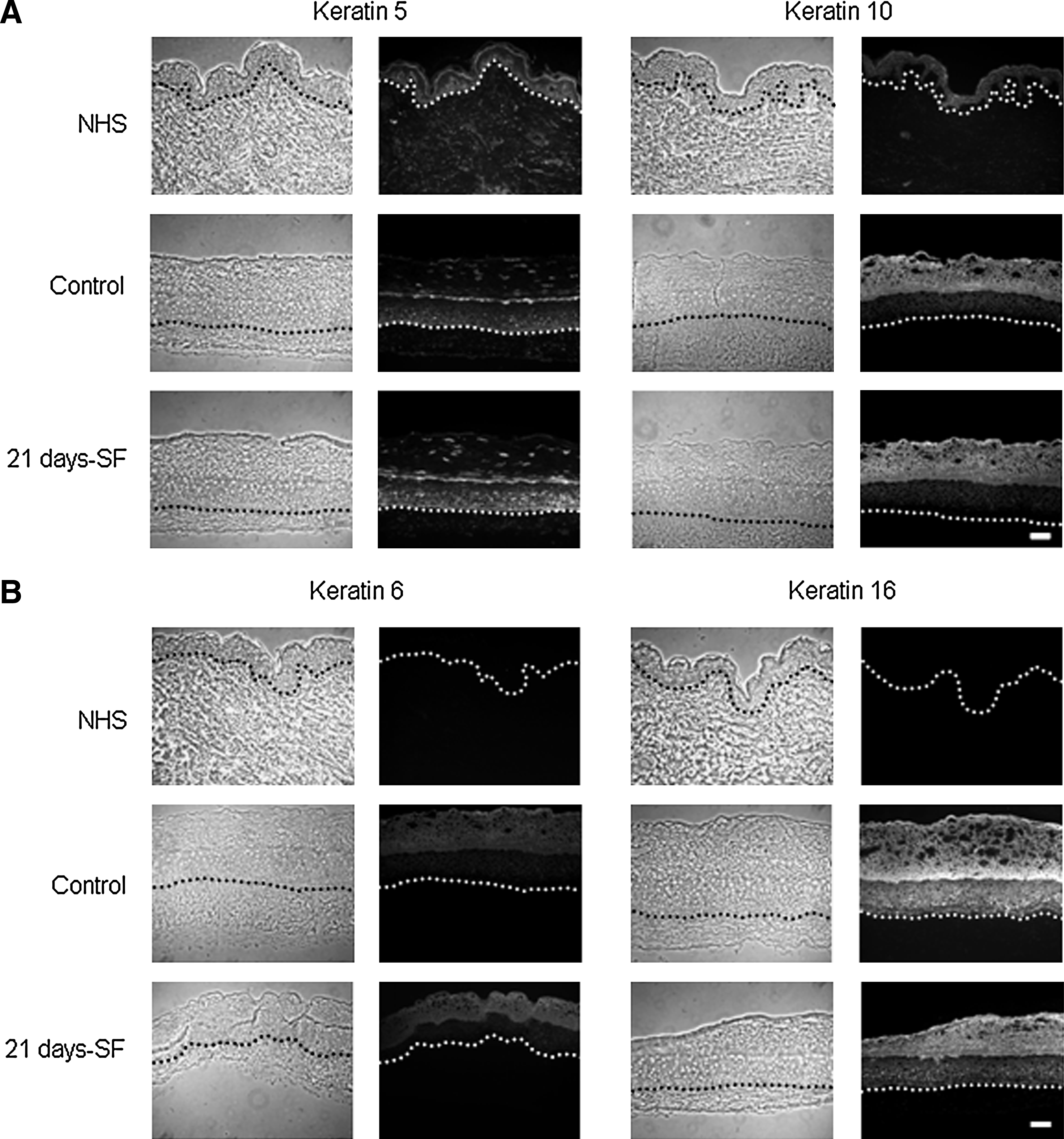
Immunolabeling of keratins:
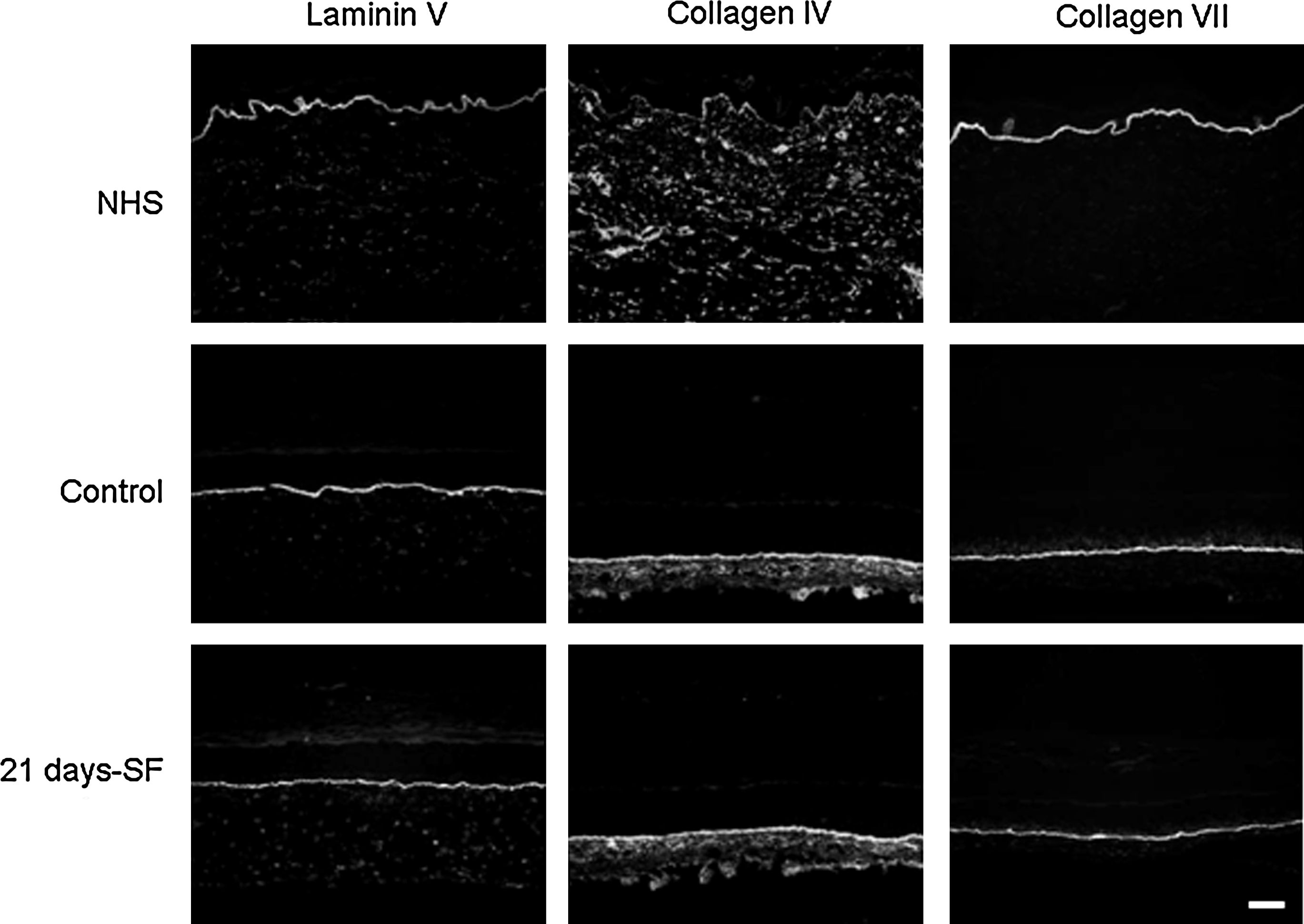
Immunolabeling of the dermo-epidermal junction: laminin V, collagen IV, and collagen VII. Results obtained from skin substitutes made with cells from a 26-year-old woman. Observations were confirmed with cells from four other patients. The nuclei were stained with Hoechst (n = 4 substitutes for each condition). Similar results were obtained with the two other culture conditions, that is, 7 and 14 days-SF (data not shown). scale bar = 50 μm. NHS, normal human skin; Control, condition with 5% serum for 21 days; 21 days-SF, serum-free medium for the last 21 days of the experiment.
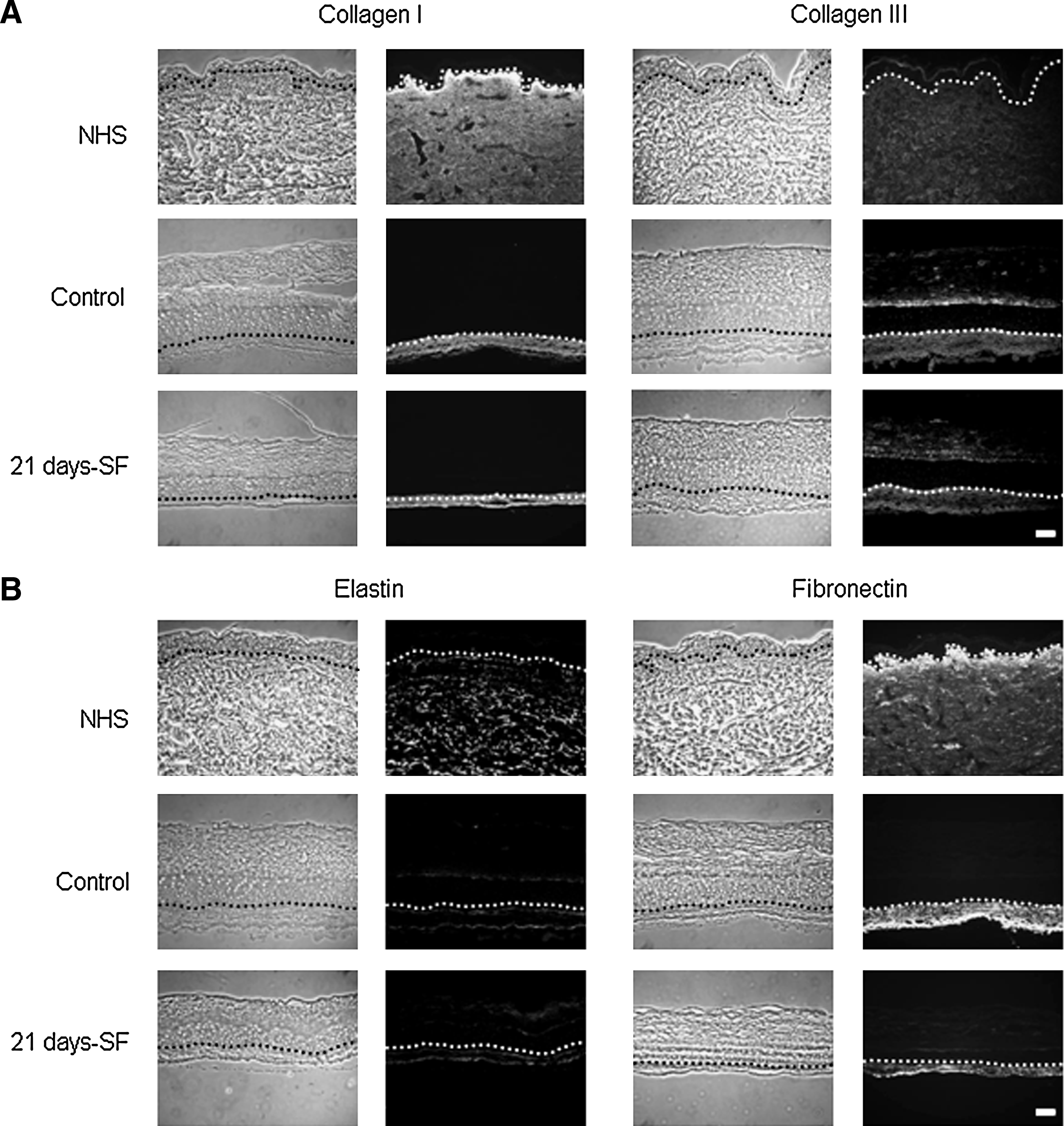
Immunolabeling of extracellular matrix:
ATR-FTIR spectroscopy
Figure 9 showed the temperature-dependent frequencies of the symmetric CH2 mode of vibration associated to the lipids of the stratum corneum and measured between the temperature ranges of 28°C–80°C by an ATR-FTIR instrument. In skin substitutes made with cells from a 26-year-old woman (cell population: ♀26), it appears that the lowest frequencies observed were associated with the stratum corneum of skin substitutes cultured without serum over the complete period of culture at the air–liquid interface (21 days-SF condition). The 14 and 7 days-SF conditions both showed similar frequency-variation patterns without any possibility to identify significant differences. The variations observed between 28°C and 80°C were as follows: 2850.27–2852.84 cm−1; 2850.64–2853.42 cm−1; and 2850.87–2853.42 cm−1 for the 21, 14, and 7 days-SF conditions, respectively, whereas the controls lead to the highest frequencies observed (2850.75–2853.77 cm−1).
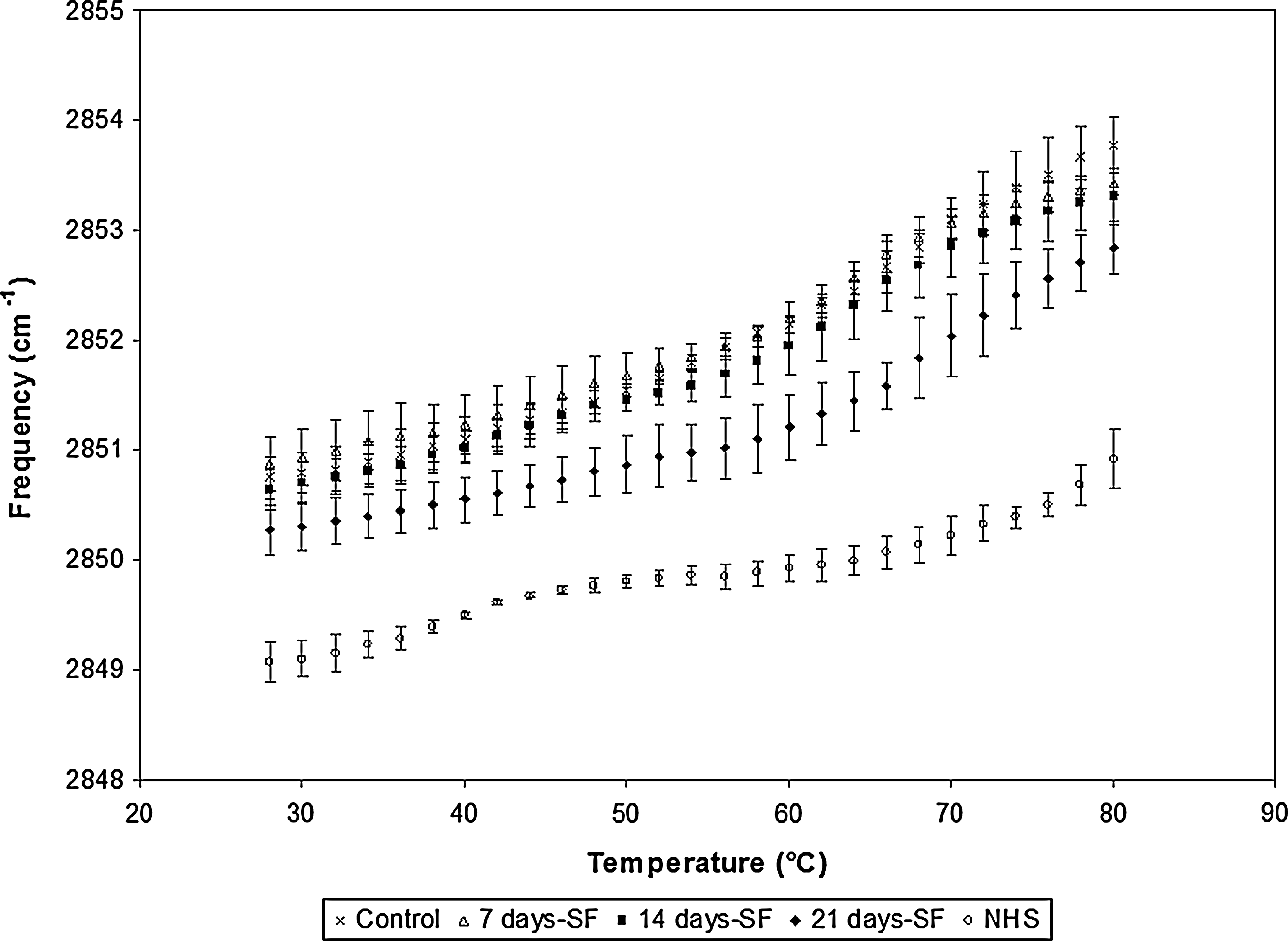
ATR-FTIR spectroscopy results obtained for skin substitutes produced with cells obtained from a 26-year-old woman. Substitutes cultured in 5% serum over the complete culture period (control), serum-free medium for 7 days (7 days-SF), serum-free medium for 14 days (14 days-SF), and serum-free medium for 21 days (21 days-SF). The experiments were performed with the five cell populations presented. ATR-FTIR, Attenuated Total Reflectance-Fourier Transform Infared.
ATR-FTIR results suggested a more disorganized lipidic system for the stratum corneum obtained with the 7 and 14 days-SF conditions because of the presence of higher frequencies associated to gauche conformers. The same observation had to be pointed out for conditions where serum is present for the complete air–liquid interface culture period (controls). Our study suggested that skin substitutes cultured in the serum-free medium over 21 days showed a significant better lipid organization of the stratum corneum for three (♀18a, ♀18b, and ♀26; Fig. 8) out of five cell populations tested compared to the 7 days-SF, 14 days-SF conditions, and even controls, whereas no significant differences for the two other cell populations tested (♀29 and ♀57) were observed (data not shown).
Percutaneous absorption
Figure 10 showed that after 8 h of hydrocortisone application, the absorption profile through the samples reached 2 ± 0%, 2 ± 1%, and 3 ± 1% for the 7, 14, and 21 days-SF conditions, respectively, compared with 2 ± 0% for controls and 1 ± 0% for NHS. It appears that there were no significant difference between skin substitutes cultured with or without serum but, most of the time, controls displayed a profile more similar to NHS than the other conditions tested. Similar results were obtained with the three other cell populations (♀18a, ♀29, and ♀57) used. It was with cells obtained from a 18-year-old woman (♀18b) that a slight difference was observed between the substitutes produced with serum over the complete experiment (controls) and those produced without serum during the culture at the air–liquid interface (21 days-SF). At time points of 8 and 24 h, the 21 days-SF condition allowed the production of skin substitutes more permeable than controls (data not shown). However, it could be argued that after 8 h, the pH of the skin surface or the hydratation of stratum corneum could affect the skin barrier integrity.
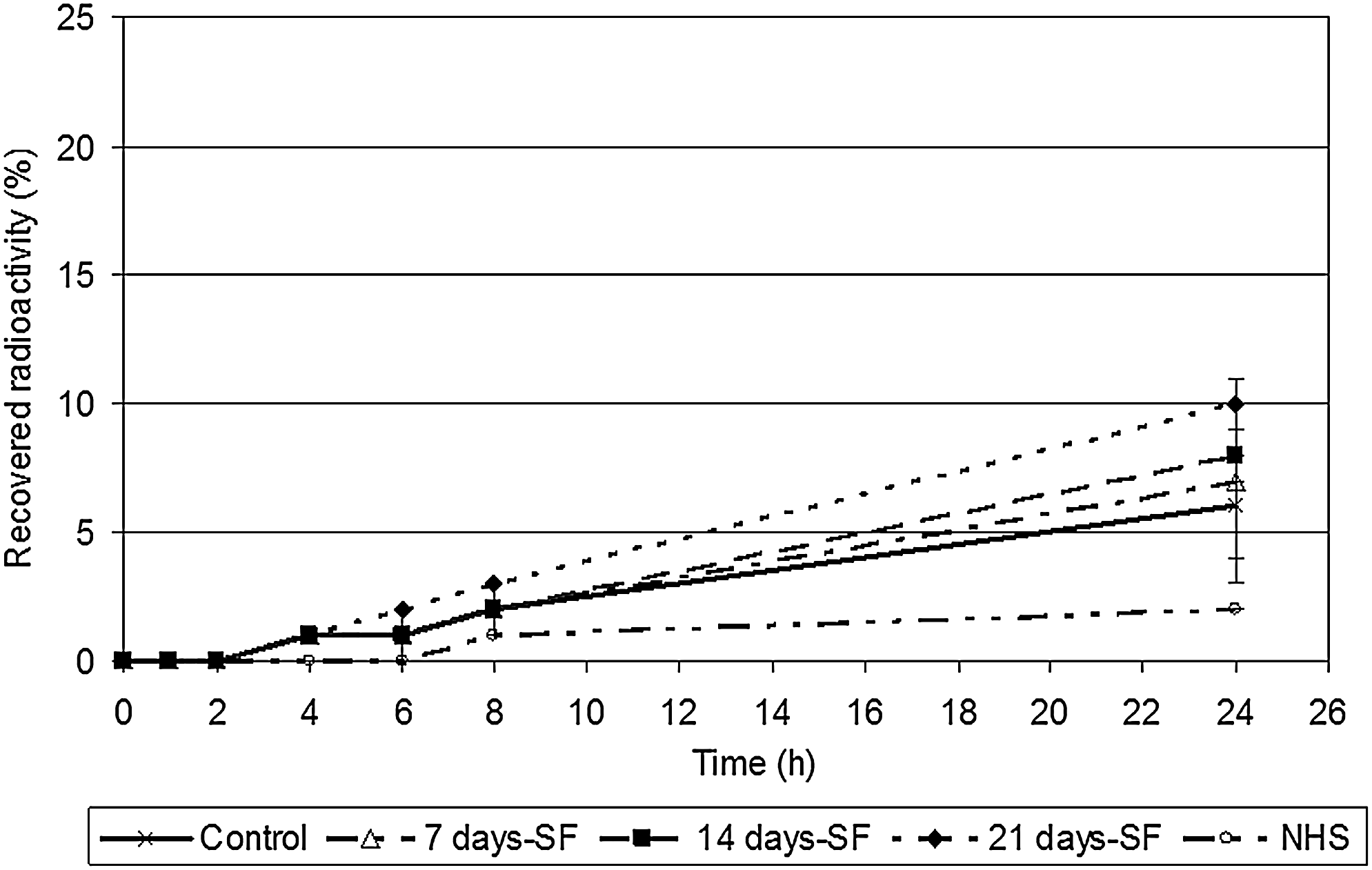
Hydrocortisone penetration profiles through skin substitutes produced with the cells obtained from a 26-year-old woman. The samples were taken after 0, 1, 2, 4, 6, 8, and 24 h. Normal human skin, substitutes cultured in 5% serum over the complete culture period (control), serum-free medium for 7 days (7 days-SF), serum-free medium for 14 days (14 days-SF), and serum-free medium for 21 days (21 days-SF). The results were confirmed with cells from three other patients (♀18a, ♀29, and ♀57; data not shown) (n = 4 substitutes for each condition).
Discussion
Serum is necessary for growth and cell proliferation. However, production is costly and composition is variable.14,19 The use of well-defined culture conditions or serum-free media was reported to improve keratinocytes terminal differentiation and the reproducibility of skin substitutes, which can be influenced by growth factors and inflammation mediators contained in calf serum.8,9,13 According to Black et al., serum-free media culture should improve reproducibility and avoid interference from ill-defined supplements. 13 The aim of the present study was to compare skin substitutes cultured in a complete medium with skin substitutes cultured in a serum-free medium over different periods of time at the air–liquid interface to optimize the self-assembly method.
In this study, we show that the use of a three-dimensional skin model, with the presence of fibroblasts and keratinocytes interactions, can produce a well-differentiated epidermis independently of the presence or absence of serum. In fact, substitutes cultured without serum show a well-developed epidermis with no difference in the epidermal thickness, suggesting that the absence of serum does not lead to either hypo- or hyperproliferation in the substitutes. Moreover, expression of differentiation markers observed in substitutes cultured in the serum-free medium is similar to what is observed in controls and in human skin.29–31 It is interesting to note the presence of hyperproliferative keratins 6 and 16 in the substitutes, but not in the NHS. These results concord with previous studies demonstrated that in vitro skin substitutes are hyperproliferative.32,33 It was demonstrated by our group that this hyperproliferation was even more important for psoriatic substitutes. 6 Further, no difference in the expression of keratins 6 or 16 was reported between control and serum-free substitutes. A large number of growth factors and cytokines have been reported to play a role in keratinocyte proliferation and differentiation.34,35 Green et al. have suggested that keratinocytes are dependant on fibroblasts or fibroblast products to grow and function properly.36,37 Effectively, even if Rosdy et al. showed that epidermal cells are able to produce hemidesmosomes and to secrete the major components of the dermo-epidermal junction in the absence of serum and dermal factors, their results suggest that the presence of dermal factors are probably necessary for the production of all the components of the lamina densa and the formation of an entire and intact basement membrane. 38 Further, Baenziger et al. have showed that human skin fibroblasts produce prostaglandin E2, which stimulates keratinocyte proliferation.39,40 Such as demonstrated by Higham et al. in a coculture model free of serum, fibroblasts provide all the necessary growth factors and extracellular matrix proteins sufficient to support keratinocyte differentiation. 41
In NHS, both keratinocytes and fibroblasts contribute to the production of proteins of the basement membrane zone. 34 In the present study, substitutes cultured in serum-free conditions display expression of laminin and collagen VII, two proteins of the basement membrane that stabilize the dermo-epidermal junction, 42 similar to that observed in human skin and controls. The starvation of serum at the air–liquid interface does not affect the synthesis of laminin and collagen VII, suggesting that if serum is removed during that step, the stability of the basement membrane is not affected and expression of basement membrane proteins is maintained. These proteins play a crucial role in the generation of a well-structured epidermis and in the regulation of normalized epithelial phenotype in a three-dimensional human skin model. 43
Collagen I, collagen III, elastin and fibronectin, four constituents of the extracellular matrix, are expressed in the dermis of the substitutes.42,44 Using serum-free conditions, expression of collagen I, collagen III, elastin and the localization of fibronectin are similar in our substitutes to what is observed in controls. A diminution of fibronectin expression, caused by the absence of exogenous fibronectin provided by the serum, is observed in serum-free medium substitutes. However, the production of fibronectin by basal epidermal cells 45 and fibroblasts 41 seems sufficient to avoid negative impact on the epidermal differentiation (previously demonstrated in Figs. 2–6). In this study, high-glucose Dulbecco's medium was used to culture the substitutes. High-glucose media are suitable for the culture of bilayer substitutes since glucose seems an important factor of the collagen synthesis. Previous publications had reported that glucose deprivation results in stimulation of protein degradation. 46 In fibroblast culture, the effects of glucose deprivation, such as a slight decrease of protein content and a marked decrease of collagen and hydroxyproline, can be observed. 47 Removal of the serum at an advanced stage of culture, the presence of high-glucose medium, and the interaction between fibroblasts and keratinocytes allow the secretion and the accumulation of growth factors necessary to maintain the quality of the extracellular matrix even after 21 days of culture at the air–liquid interface. Our immunolabeling results are in accordance with previous works of Black et al., which demonstrate that starvation of serum at an advanced state of culture does not bring any negative change of the morphological characteristics of the serum-free substitutes. 13
Stratum corneum is the product of the keratinocyte differentiation process. 48 To be efficient, lipids of the stratum corneum must be well organized. 1 Higher frequencies in CH2 bands are generally characteristic of high content of gauche conformers, whereas bands at lower frequencies are associated to the presence of trans conformers. The study of the variation of frequencies in function of temperature allows the observation of a phase transition from an ordered system to a disordered conformation. Our ATR-FTIR results do not show any significant differences in the CH2 stretching mode, meaning that lipidic system of the stratum corneum of the substitutes cultured in a serum-free medium are not more disorganized. Contrariwise, the ATR-FTIR spectroscopy results indicate that a serum-free medium helps the substitutes create a better lipid organization in three out of five cell populations, suggesting that serum deprivation could be a good choice for a better control. In fact, these results are in accordance with previous studies. Effectively, it is known that vitamin A, a component of serum, has an inhibitory effect on the differentiation of keratinocytes.49,50 Recently, Breiden and colleagues demonstrated that vitamin A precursor reduced the ceramide levels, the major component of the stratum corneum. 50
Percutaneous absorption results show no significant difference in the penetration profile of hydrocortisone after serum deprivation. In fact, percentages of recovered radioactivity are similar between skin substitutes cultured with or without serum. This suggests that the permeability profile of skin substitutes cultured in a serum-free medium is not adversely affected relative to controls. These results are in accordance with those obtained with ATR-FTIR analysis. Here, the better lipid organization observed with the ATR-FTIR results did not lead to an increased impermeability of the skin substitute. An explanation to this is related to the fact that percutaneous absorption is a technique that allows to observe the permeability in a global way, whereas the ATR-FTIR technique gives a more focused scrutiny of the permeability process. 1 More specifically, on one hand, the percutaneous absorption studies allow to observe the penetration of a drug through all skin components with a preference for cells or lipids depending on the physicochemical properties of the drug used. In this study, we tested hydrocortisone, which is well known for its lipophilic character. In brief, in the stratum corneum, hydrocortisone can pass through the intercellular lipids and through the lipids of the cornified envelope. On the other hand, the ATR-FTIR spectroscopy allows to specifically observe the organization of targeted components such as the intercellular lipids, thus providing results from a different angle. In fact, the two techniques are complementary.
To summarize, there is no obvious difference observed in the results of the three serum-free medium conditions (serum-free medium over 7, 14, or 21 days). According to our results, it could be possible to use any of those serum-free conditions. However, the 21 days-SF condition shows even lower frequencies of the CH2 stretching mode in three out of five cell populations compared with the three other culture conditions. Consequently, we would recommend this condition in future studies if controlled parameters are required: (1) to avoid any unforeseen interactions, (2) to reduce variations that could be brought by the presence of serum over a period of time, (3) to have a better control, and (4) to reduce production costs.
Taken together, the results presented here prove that it is possible to obtain an efficient keratinocyte differentiation when serum starvation begins at an advanced stage of culture. The presence of fibroblasts supports and provides a feeder layer that prolongs life and growth of keratinocytes13,51,52 thus counteracting the lack of factors present in serum. Such as discussed by Higham et al., in complete medium, proteins will be provided by the serum, but in serum-free condition the cells themselves may secrete the proteins. 41 Our observations agree with results obtained in a serum-free coculture model produce by Higham et al., who demonstrated that when irradiated fibroblasts and keratinocytes were cocultured with serum, the contribution of the fibroblasts to keratinocyte proliferation was marginal compared with their contribution under serum-free conditions. 41
This research concludes that the absence of serum in the medium at the air–liquid interface does not compromise cell proliferation and differentiation of the epidermis, expression of dermo-epidermal and dermal components, physico-chemical characteristics, and permeability profile to hydrocortisone in the skin substitutes. Finally, substitutes produced in controlled conditions would make it easier to meet the strict regulatory compliance imposed for clinical uses (burn patients and diabetic ulcers) and provide even more controlled production conditions for fundamental researches (percutaneous absorption, metabolic and toxicologic studies).1,4–6
Footnotes
Acknowledgments
This study was supported by the Canadian Institutes of Health Research (MOP-86498; MOP-90283). R.P. was recipient of a research fellowship from the “Fonds de la Recherche en Santé du Québec,” Québec, QC, Canada. J.J. held a scholarship from “Fonds d'enseignement et de recherche” of the Faculté de Pharmacie, Université Laval, Québec, QC, Canada. Special thanks to Dr. Dan Lacroix for the revision of the article.
Disclosure Statement
No completing financial interests exist.