
Abstract
Currently, there is no effective treatment available to patients with irreversible loss of functional salivary acini caused by Sjogren's syndrome or after radiotherapy for head and neck cancer. A tissue-engineered artificial salivary gland would help these patients. The graft cells for this device must establish tight junctions in addition to being of fluid-secretory nature. This study analyzed a graft source from human salivary glands (huSG) cultured on Matrigel. Cells were obtained from parotid and submandibular glands, expanded in vitro, and then plated on either Matrigel-coated (2 mg/mL) or uncoated culture dish. Immunohistochemistry, transmission electron microscopy, quantitative real-time–polymerase chain reaction, Western blot, and transepithelial electrical resistance were employed. On Matrigel, huSG cells adopted an acinar phenotype by forming three-dimensional acinar-like units (within 24 h of plating) as well as a monolayer of cells. On uncoated surfaces (plastic), huSG cells only formed monolayers of ductal cells. Both types of culture conditions allowed huSG cells to express tight junction proteins (claudin-1, -2, -3, -4; occludin; JAM-A; and ZO-1) and adequate transepithelial electrical resistance. Importantly, 99% of huSG cells on Matrigel expressed α-amylase and the water channel protein Aquaporin-5, as compared to <5% of huSG cells on plastic. Transmission electron microscopy confirmed an acinar phenotype with many secretory granules. Matrigel increased the secretion of α-amylase two to five folds into the media, downregulated certain salivary genes, and regulated the translation of acinar proteins. This three-dimensional in vitro serum-free cell culture method allows the organization and differentiation of huSG cells into salivary cells with an acinar phenotype.
Introduction
In an effort to treat these patients, we have been working to develop an artificial salivary gland device.7–9 Recently, we developed a method to obtain primary salivary epithelial cells capable of forming a polarized monolayer.10,11 These cells, termed human salivary gland (huSG) cells, expressed tight junctions (TJs) and showed reasonable transepithelial electrical resistance (TER). However, huSG were mainly ductal cells and did not express the main water channel protein aquaporin-5 (AQP-5); thus, they were incapable of unidirectional fluid secretion. A major hurdle in the development of this artificial salivary gland device has been finding a suitable fluid-secretory graft cell. 12 Also, to better understand the functional capabilities of salivary acini, we need an in vitro model to expand and study acinar secretory cells.
Cells cultured on Matrigel allow the in vitro modeling of cell behavior, including differentiation, apoptosis, pathophysiology, and cancer invasion. 13 Interestingly, Matrigel has been used to stimulate and maintain differentiation of several cell types,14–19 including primary salivary cells. 20 Recently, Szlavik et al. (2008) cultured primary human salivary acinar cells that expressed acinar as well as TJ markers on Matrigel. Their culture model required the use of animal serum, which is known to promote fibroblast overgrowth in epithelial cultures. 20 The risk of infection increases when animal-derived components are used in culture. In this study we aimed to modify our previous huSG culture method to obtain differentiated acinar cells using serum-free Hepato-STIM medium and Matrigel. We analyzed the primary huSG from both parotid and submandibular salivary glands.
Materials and Methods
Source of huSG tissue
Biopsies of human submandibular (n = 9) and parotid (n = 3) salivary glands were obtained from the Department of Pathology, University of Virginia, through the Cooperative Human Tissue Network and the Department of Pathology, Royal Victoria Hospital, McGill University. For tissue use, we received approval from the Institutional Review Boards at McGill University. The glands were from eight men and four women who had received no prior radiotherapy/chemotherapy. These glands were excised as part of the resection of head and neck cancers. All glandular tissues were judged as histologically normal by a surgical pathologist.
Tissue culture
Tissue biopsies (0.5–1.5 g) of huSG were prepared as previously described. 10 In brief, they were mechanically minced with a pair of curved scissors, dissociated enzymatically, and the cell pellet filtered through a 70 μm strainer. For all experiments described in this article, huSG cells from passage 2 and 3 were employed at a density of 5 × 104 cell/cm2. Cells were seeded on (1) 24-mm Transwell-Clear filters for TER measurements, (2) 8-well slide chambers for confocal microscopy and apoptosis analyses, (3) 96-well, 12-well, or 6-well dish for thiazolyl blue tetrazolium bromide (MTT) assay, transmission electron microscopy (TEM), Western blot, and quantitative real-time–polymerase chain reaction (RT-PCR) analyses, respectively. Cells were cultured on either noncoated or Matrigel-coated (19.6 mg/mL; BD Biosciences) surfaces. Matrigel was thawed on ice and diluted in Dulbecco's modified Eagle's medium (1:6, final concentration = 2 mg/mL). The plates/slide chambers/polyester filters were coated with a thin layer of this diluted Matrigel and incubated at 37°C for 30–60 min before cell seeding. For example, one well in an eight-well slide chamber of 0.8 cm2 surface area received 30 μL of this diluted Matrigel mix. The Matrigel acted as a rigid thin layer that allowed cells to grow on it. MDCK-II cells obtained from BD Biosciences Clontech were maintained in Dulbecco's modified Eagle's medium/F-12 supplemented with 10% fetal bovine serum.
Measurements of TER
huSG cells were seeded on 24-mm Transwell-Clear polyester filters (Corning Life Sciences). TER was measured at day 3, 5, and 7 in six separate determinations, using a Millicell ERS epithelial volt-ohmmeter (Millipore). MDCK-II cells were used as a positive control. TER readings from filters without cells (blank or Matrigel alone) were subtracted from readings obtained from filters seeded with cells.
Evaluation of apoptosis
An ApopTag® peroxidase in situ apoptosis detection kit (Chemicon International) was used. Apoptotic reactions were performed in eight-well slide chambers (five slides per culture type) at day 7 and scored at 400 × magnification. In addition, the media from these eight-well slide chambers were collected and cytospin was used to collect any floating/detached cells, which were then included in the apoptosis analysis. The slides were assessed by two observers from 10 randomly chosen fields per slide.
MTT cell proliferation assay
MTT cell assay (Invitrogen) was performed to evaluate cell viability and proliferation of huSG cells grown on plastic and on Matrigel at days 0, 1, 3, 5, 7, and 9.
Immunofluorescence imaging
The following primary antibodies were used to characterize huSG grown on plastic and on Matrigel: rabbit anti-claudin-1 (CLDN-1) (Cat. 51-9000), CLDN-2 (Cat. 51-6100), CLDN-3 (Cat. 34-1700), occludin (Cat. 42-2400), JAM-A (Cat. 36-1700), mouse anti-ZO-1 (Cat. 33-9100), and CLDN-4 (Cat. 32-9400) from Zymed; Mouse anti-cytokeratin (panel, Cat. 18-0132), vimentin (Cat. 18-0052), and α-smooth muscle actin (Cat. 080106) from Invitrogen; Goat anti-AQP-5 (Cat. SC-28628) and mouse anti-mucin-1 (Cat. SC-25274) from Santa Cruz; rabbit anti-α-amylase (anti-AMY) (Cat. A8273) from Sigma Aldrich; and goat anti-Musashi-1 (Cat. AF2628) and mouse multipotent mesenchymal/stromal cell marker antibody panel kit (Cat. SC017) from R&D Systems. The panel contains a group of antibodies for the positive identification of human mesenchymal/stromal cells; anti-Stro-1, anti-CD90, anti-CD106, anti-CD105, anti-CD146, anti-CD166, anti-CD44, plus leukocytes markers; anti-CD19 and anti-CD45. The antibodies used are reactive against the respective proteins from human and other species. In addition, we used nonimmune goat (R&D Systems, Cat. AB-108-C), rabbit, and mouse (Zymed Labs; Cat. 08-6599 and 08-6199) isotype control antibodies. Cells were incubated with a primary antibody (diluted 1:100) overnight at 4°C. Afterward, cells were incubated with either donkey anti-mouse/rabbit/goat-fluorescein isothiocyanate-conjugated (FITC) or -Rhodamine Red-X-conjugated (RRX; Jackson ImmunoResearch) secondary antibodies for 1 h at room temperature (diluted 1:100) in dark. Finally, 4′,6-diamidino-2-phenylindole, dihydrochloride (Invitrogen) was added for 3–5 min. Fluorescence images were taken using a Leica DM6000 fluorescent microscope equipped with Volocity software (PerkinElmer). Images shown are representative of at least five separate experiments with multiple images taken per slide.
Western blot analysis
Conditioned media from wells were collected. Then, huSG cells (including detached/floating cells) grown on plastic or on Matrigel (at 1,3,5, and 7 days) were lysed in 200 μL/well of cold RIPA buffer. Lysates were collected and centrifuged at 10,000 g for 20 min to pellet the cell debris and keep the supernatants for further analysis. Protein samples of 60 μg each (measured by the Thermo Scientific Pierce BCA Protein Assay Kit) from cell lysate supernatants and conditioned media were immune-blotted overnight with rabbit anti-AMY antibody (1:2000 dilution; Sigma Aldrich, Cat. A8273) at 4°C. We also used a nonimmune rabbit isotype control antibody (Zymed Labs, Cat. 08-6599). Protein bands were visualized on X-ray film. All experiments were performed in duplicate and repeated at least three separate times.
Transmission electron microscopy
huSG cells cultured on uncoated and Matrigel-coated 12-well plates were collected (including floating/detached cells) and fixed (at days 1, 2, 3, 5, and 7) in 2% formaldehyde and 2% glutaraldehyde in 0.1 M cacodylate buffer (pH 7.4) for 2 h, rinsed in 0.1 M cacodylate buffer, and postfixed in 1% osmium tetroxide for 1 h. After rinsing in cacodylate buffer, samples were dehydrated through ethanol series and infiltrated and embedded in Epon812 (Electron Microscopy Sciences). Thin sections (70 nm) were cut with a diamond knife and mounted onto copper grids. Grids were examined at various magnifications using a 100-CXII TEM (JEOL) at an accelerating voltage of 120 kV.
Quantitative RT-PCR analysis
At days 3, 5, 7, and 9, total RNA of huSG cells grown on plastic and on Matrigel were isolated using the RNeasy micro kit (Qiagen). The concentration of RNA was determined using Qubit (Molecular Probes). Total RNA (2 μg per sample) was reverse transcribed using the high-capacity cDNA Archive Kit (Applied Biosystems) in 20 μL volume. For PCR amplification, 5% of the cDNA was used with RT-PCR primers and 6-carboxy-fluorescein-labeled minor groove binder probes. The probes and primers for CLDN-1 (Assay ID: Hs00154575_m1), cytokeratin-18 (CK-18; Assay ID: Hs01941416_g1), epidermal growth factor (EGF; Assay ID: Hs00610104_m1), kallikrein-1 (KLK-1; Assay ID: Hs00158490_m1), AMY (Assay ID: Hs00420710_g1), AQP-5 (Assay ID: Hs00387048_m1), and glyceraldehyde-3-phosphate dehydrogenase (GAPDH Assay ID: Hs99999905_m1, used as an endogenous reference) were selected from the Applied Biosystems. PCRs (20 μL) were performed in duplicates using TaqMan Universal Master Mix (Applied Biosystems) on a Prism Sequence Detection System 7500 (Applied Biosystems) with the default settings (50°C for 2 min, 95°C for 10 min, and 40 cycles [95°C for 15 s, 60°C for 1 min]). Every culture experiment was repeated at least five times. Gene expression levels were calculated by normalizing the target RNA value to the value of GAPDH in the same sample. Results are expressed as fold-changes in gene expression relative to control huSG sample cultured on plastic at day 9. This sample was chosen as our standard control as it was the last day of all experiments and had the lowest level of expression for most genes tested. Therefore, comparing all results to day 9 would show higher levels of gene expression and would be easier for interpretation.
Statistical analysis
Data are presented as means ± standard deviation of results from three or more separate experiments. The data were analyzed by one-way analysis of variance where p-value <0.05 represents significant differences between groups at specified times. Results obtained from immunofluorescence technique are based on the examination of five slides per culture type. At least 800 cells were examined per culture type and the expression level of each marker/protein is indicated as a percentage.
Results
Morphological changes
The majority of huSG cells (90%–95%) grown on plastic were of a striated ductal phenotype, an observation similar to what we reported in a previous study. 10 In the current study, huSG cells formed epithelial-like isolated clusters on plastic (Fig. 1A). On Matrigel-coated surfaces (2 mg/mL), huSG cells in proximity to each other were able to self-assemble into three-dimensional (3D) acinar-like units, (15% of cells were observed with this 3D structure within 24 h of seeding; Fig. 1B). Cells located further from each other started to divide and formed a cell monolayer. In some areas of the culture dish, the 3D structures formed on top of the continuous huSG monolayer, without interfering with each others. Initially, these 3D units consisted of 6–8 cells and then increased in size/number (10–12 cells) at day 3. At day 5, some 3D units started to detach/separate from the bottom of the culture dish. These 3D units were still vital based on ultrastructure analysis by TEM. At day 7, these detached 3D units started to disunite into single cells, which were apoptotic based on TEM and apoptosis analyses. At day 9, 30%–50% of 3D units were seen floating in the media. All cells (dead and alive) were included for the Western blot of cell lysates. However, for the analysis of proteins secreted in the culture media (such as amylase) by Western blots, all cells were excluded.
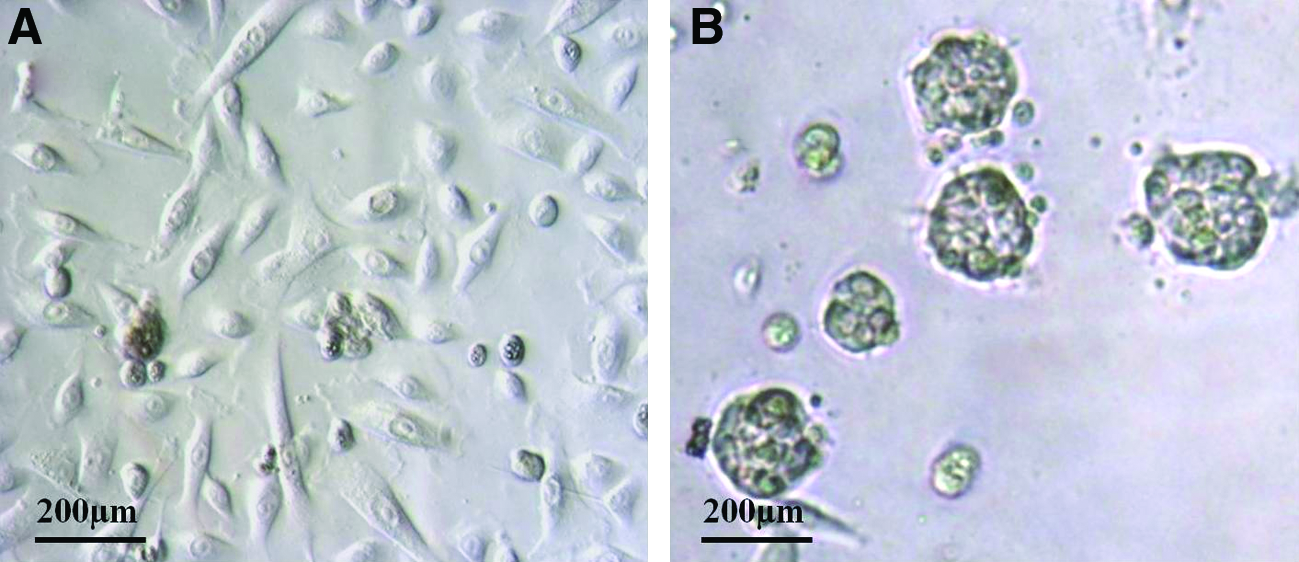
The huSG samples used in this study were judged free of any pathological changes (both by the surgical- and histopathology reports). All patients had their glands removed within a safety margin from the primary tumor, and none of these patients had a salivary gland tumor. Therefore, we considered huSG cells used here as normal. huSG's cell growth was much slower than that of the cell line HSG, obtained from a salivary gland tumor. We were able to culture huSG cells until passage 10 without observing any apparent cell behavior changes. No differences in cell culture results were seen between the gland types and patients' gender. Patients' age ranged between 20 and 73 years. We observed that 3D acinar units from old patients (60–70 years) appeared less organized relative to those from younger patients (20–52 years). In addition, younger patients' cells reached confluence faster than those from older patients.
Measurements of epithelial barrier (TJs) formation
TER levels of huSG cells were compared to those of MDCK-II (positive control) cells at days 3, 5, and 7 from six separate determinations. TER levels of huSG cells both on plastic (381–415 ′Ω cm2) and on Matrigel (407–415 ′Ω cm2) were significantly different from those of MDCK-II (470–601 ′Ω cm2; p < 0.01). This was expected because kidney cells have tighter junctions than those found in salivary glands. At day 3 only, TER levels of huSG cultured on Matrigel were significantly higher than those of huSG cultured on plastic (p < 0.01; Fig. 2). Both huSG culture types (i.e., with or without Matrigel) exhibited moderate TER levels, and thus could equally form a functional epithelial barrier.

TER measurements at days 3, 5, and 7. Both huSG culture types (on plastic and on Matrigel) exhibited moderate TER levels, and thus could equally form a functional epithelial barrier. TER levels of huSG cells both on plastic and on Matrigel were significantly different from those of MDCK-II at days 3, 5, and 7 (*p < 0.01). Only at day 3 the TER levels of huSG cultured on Matrigel were statistically significant from those of huSG cultured on plastic (*p < 0.01). TER, transepithelial electrical resistance; SD, standard deviation.
Apoptosis versus proliferation
The apoptotic activity of huSG cells increased significantly (evaluated at day 7, p < 0.01) when grown on Matrigel (26.6%) than on plastic (10.5%) (Fig. 3). Apoptotic cells were mainly seen at the center of intact 3D units where a lumen is being formed, as well as from floating dead cells collected by cytospin. MTT cell assay revealed a significant (p < 0.05) decrease in the viability and proliferation activity of huSG cells on Matrigel (at day 7 and 9), when compared to cells grown on plastic (Fig. 4).
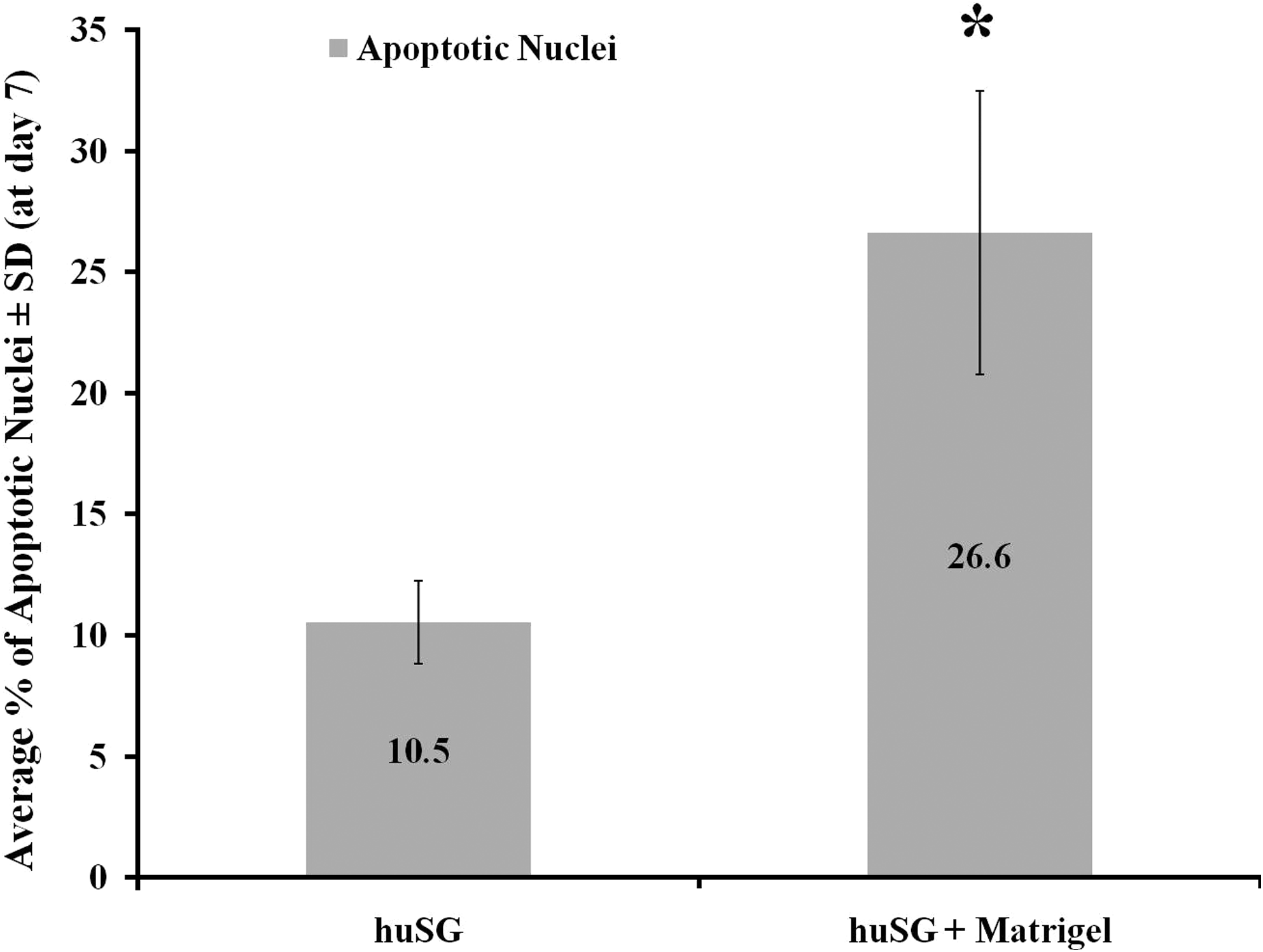
Apoptotic activity was evaluated at day 7 for huSG cultured on plastic and on Matrigel (using ApopTag kit). Apoptotic activity increased significantly (*p < 0.01) on Matrigel (26.6%) as compared to huSG on plastic (10.5%). Results are based on the examination of five slides per culture type. At least 800 cells were examined per slide.

Proliferation and viability activities of huSG cells cultured on plastic and on Matrigel were evaluated using MTT assay (from day 0 to 9). Proliferation and viability activities decreased significantly (*p < 0.05) on Matrigel at days 7 and 9 as compared to huSG on plastic. Results are reported as means of cell numbers from parallel cultures.
Characterization of huSG cells with immunofluorescence staining
Both huSG cultures expressed all TJ proteins tested: CLDN -1, -2, -3, and -4; occludin; JAM-A; and ZO-1 in comparable percentage (Table. 1A; Fig. 5). On Matrigel, huSG cells expressed the acinar-specific proteins AMY (99%), AQP-5 (99%), and mucin-1 (50%). Adhesion-related proteins such as CD44 (99%) and CD166 (99%) (Fig. 6), cytokeratins (99%), and vimentin (99%) were also expressed (Table 1B). However, huSGs were negative for α-smooth muscle actin, Musashi-1, CD105, CD106, CD90, CD146, Stro-1, CD45, and CD19 (Table 1B, C). On plastic, huSG cells were negative for AQP-5 and mucin-1 (Table 1B). This result indicated that Matrigel provided the appropriate cues for a differentiation into an acinar phenotype.

Immunofluorescence micrographs (Volocity software) of huSG cultured on Matrigel for 5 days express all TJ proteins tested: Label
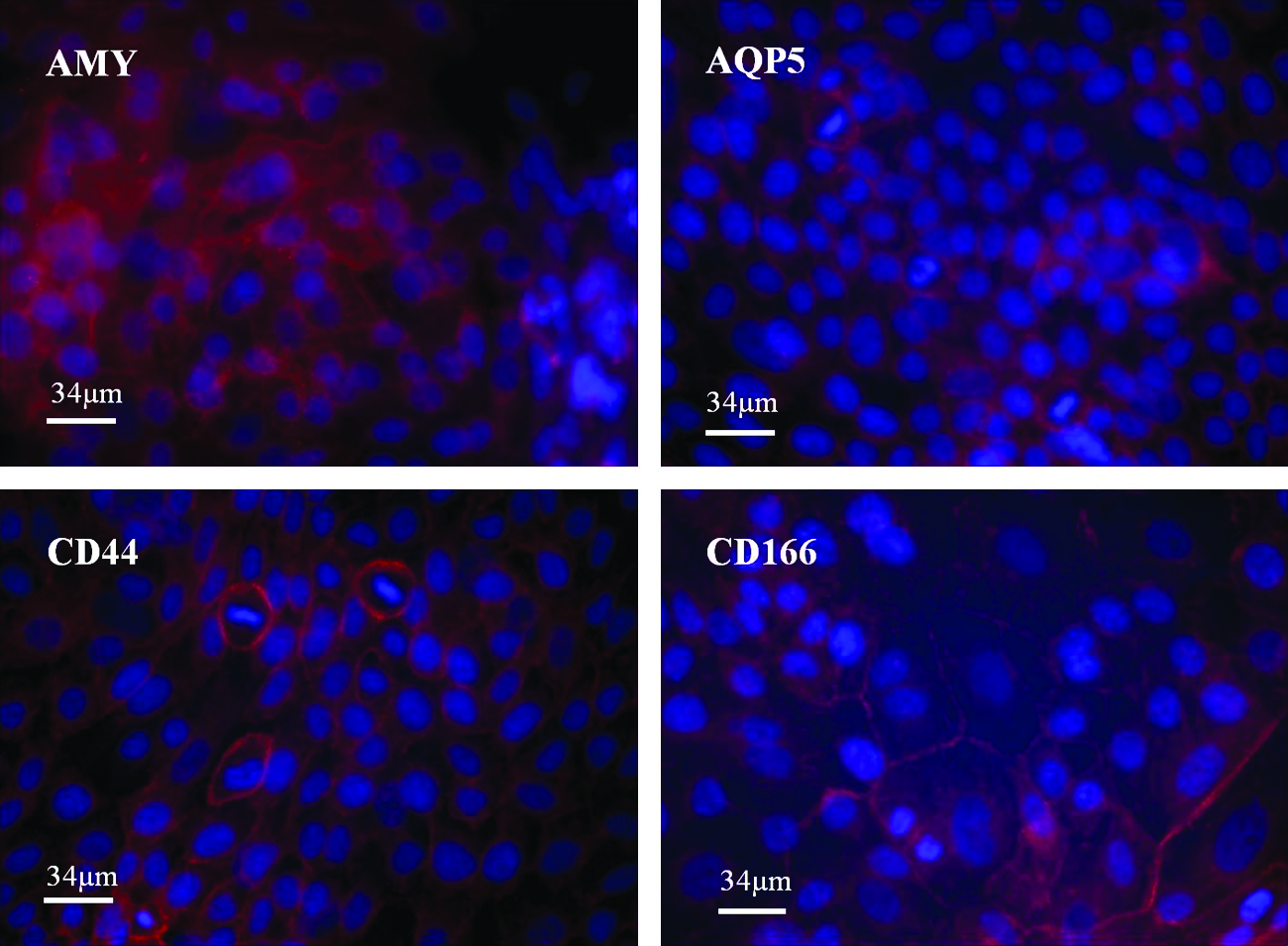
Immunofluorescence micrographs of huSG cultured on Matrigel for 5 days. Expression of acinar secretory proteins AMY, water channel AQP-5, and adhesion-related proteins CD44 and CD166 (shown in red). Nuclei are stained with 4′,6-diamidino-2-phenylindole, dihydrochloride (shown in blue). Scale bar = 34 μm, × 400. AMY, α-amylase; AQP-5, aquaporin-5. Color images available online at www.liebertonline.com/tea
Results reported upon counting at least 800 cells per culture type. The frequency of each marker/protein is indicated as a percentage. (A) Tight junction proteins: claudin-1, -2, -3, and -4; occludin; JAM-A; and ZO-1. (B) Cytokeratins panel (epithelial), vimentin (mesenchymal, salivary ductal, immature acinar, and their progenitors), α-SMA (myoepithelial), α-amylase (serous acinar), AQP-5 (acinar), and mucin-1 (mucous acinar). (C) Stem cell markers; Musashi-1 (neural), Stro-1, CD44, CD166, CD105, CD106, CD90, CD146 (mesenchymal), CD45, and CD19 (leukocyte).
huSG, human salivary glands; α-SMA; α-smooth muscle actin; AQP-5, aquaporin; Msi-1, Musashi-1.
Western blot analysis
AMY synthesis and secretion were evaluated at days 1, 3, 5 and 7. Both huSG cells cultured with or without Matrigel could synthesize and secrete AMY into the culture media. Results from the cell lysates were similar to those from the conditioned media. The steady-state content of AMY inside the cells and the content of AMY in the media as it accumulates over time, together, reflect the balance between the rate of AMY synthesis and the rate of degradation. Matrigel increased the synthesis and secretion of AMY protein from two- to five-folds, when compared to huSG cultured on plastic (Fig. 7).

Western blot analysis of AMY (60 kDa) from conditioned media of huSG cells (at days 1, 3, 5, and 7). When cultured on Matrigel, huSG cells secreted two to five times more AMY protein into the media than when cultured on plastic.
Transmission electron microscopy
Both huSG culture types exhibited TJ structures at the apicolateral cell membranes, confirming that huSG cells were polarized. However, no basal lamina was observed. Ultrastructural analysis confirmed that huSG cells cultured on Matrigel were comparable to normal human acinar cells. Their apical cytoplasm was packed with electron-dense secretory granules (Fig. 8A), resembling those of normal human salivary secretory granules. 21 Only huSG cells cultured on Matrigel showed features of active protein synthesis with many Golgi saccules (Fig. 8B) and rough endoplasmic reticulum. The huSG cultured on plastic retained their striated duct cell phenotype. 10 Their cytoplasm contains multiple large mitochondria, tonofilaments, and small isolated islands of glycogen. No Golgi apparatus, rough endoplasmic reticulum, secretory granules, or microvilli were observed (Fig. 8C).
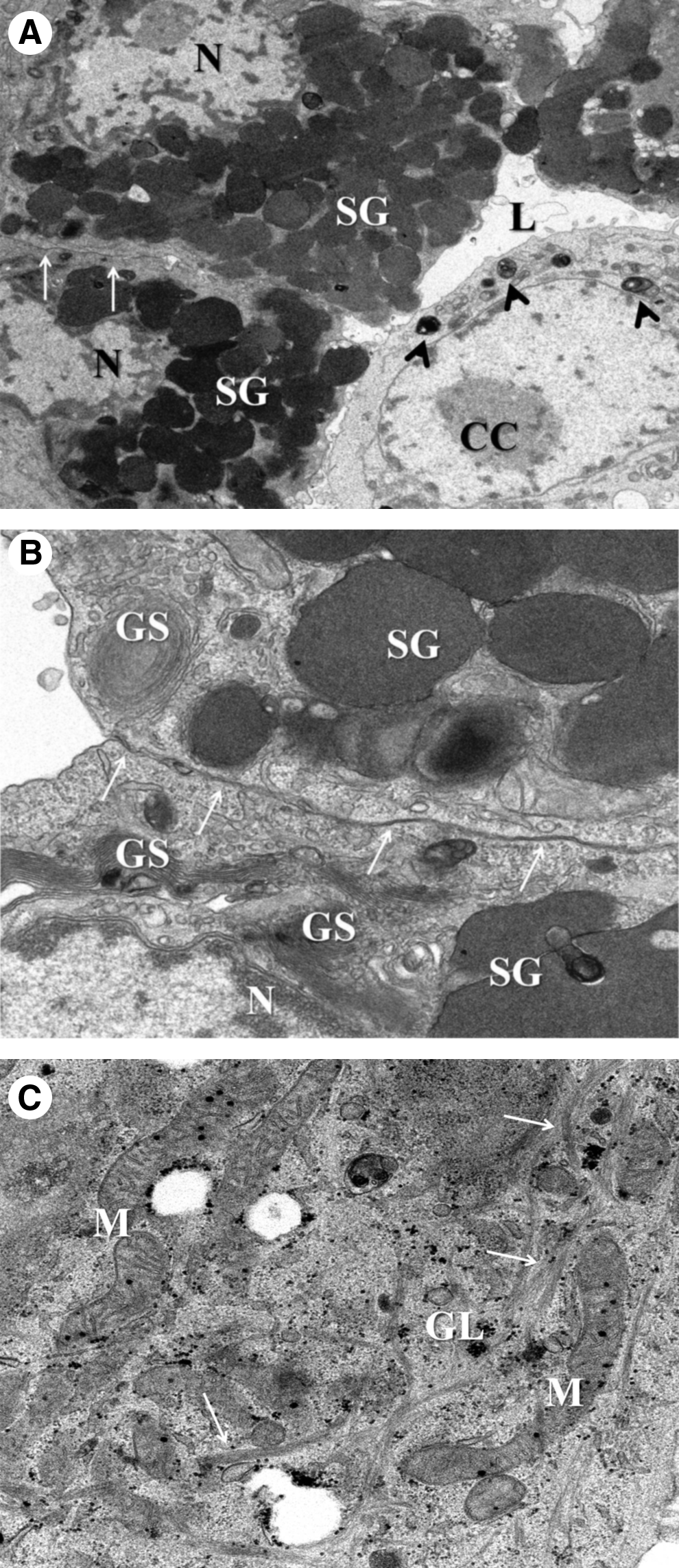
Transmission electron microscope micrograph of huSG cells.
At day 3 on Matrigel, 60% of huSG cells (from both the monolayer and 3D units) had secretory granules. At days 5 and 7, this proportion increased to 90%. The huSG 3D units were well organized around central lumens and cells in proximity to the lumen possessed more secretory granules than those at the periphery of the 3D units (Fig. 8A). This suggests that the huSG cells adopted an acinar phenotype. Cells in the middle of the 3D unit showed signs of apoptosis, including detachment of chromatin from the nuclear envelope and chromatin condensation at one side of the nucleus along with degradation of cytoplasmic organelles enclosed into multiple lysosomes (Fig. 8A).
Quantitative RT-PCR analysis
Gene expression levels were measured in five patients. Results are reported as fold-changes in gene expression relative to control sample (huSG cultured on plastic for 9 days) of at least five separate experiments. On Matrigel, the expression of TJ protein CLDN-1, acinar-specific markers (AMY and AQP-5), and ductal-specific genes (CK-18, EGF, and KLK-1) were decreased. This downregulation was statistically significant (p < 0.05) for the CLDN-1, AMY, AQP-5, CK-18, and KLK-1 genes. Figure 9 shows quantitative RT-PCR results at day 3. Similar trends were observed at days 3, 5, and 7. These data suggest that the protein expression pattern associated with huSG cultured on Matrigel was achieved through the regulation of translation, rather than the transcription of these relevant genes. Gene expression levels were different among patients and this might be related to the patients' age.

Gene expression levels (at day 3) of huSG cultured on Matrigel and on plastic for CLDN-1, AMY, AQP-5, CK-18, EGF, and KLK-1 genes. At least 5 separate experiments were performed per gene. Results are reported as means (fold change) of parallel cultures (*p < 0.05). The protein expression pattern associated with huSG cultured on Matrigel was achieved through the regulation of translation rather than the transcription of these relevant genes. CK-18, cytokeratin-18; EGF, epidermal growth factor; KLK-1, kallikrein-1.
Discussion
This study describes a convenient and reproducible method (on Matrigel) to grow and maintain polarized acinar units from primary human salivary cells, as a 3D in vitro model to study the regeneration of salivary glands. We report the ability of huSG cells cultured on Matrigel to differentiate into 3D polarized acinar units. These cells expressed TJs proteins (such as CLDN-1, -2, -3, and -4; occludin; JAM-A; and ZO-1), acinar proteins (AQP-5, AMY, and mucin-1), and adhesion-related proteins (CD44 and CD166). Moreover, the secretion of AMY was increased two- to five-folds. TEM confirmed the formation of differentiated acinar polarized cells with numerous secretory granules. Quantitative RT-PCR revealed downregulation of several salivary genes along with regulation of the translation of their corresponding proteins (confirmed by immunofluorescence analysis).
We began to culture huSG cells for the purpose of seeding a tissue-engineered artificial salivary device. Accordingly, it was critical for us to establish that huSG cells could form TJs and were appropriately polarized. Other investigators have reported successful cultures of human salivary epithelial cells.22–25 However, none of them used functional assays of polarization (TER) to examine their cells. Epithelial cells must be polarized to secrete fluid unidirectionally. In general, matrix proteins of the basement membrane guide epithelial cells to differentiate. 26 Formation of TJs is critical to establish cell polarity and unidirectional secretion. In this study, Matrigel was a substitute for the components of the basement membrane. However, TEM examination did not show a basal lamina. TEM confirmed the establishment of TJ structures in huSG cells (in both monolayers and 3D acinar units) and thus cell polarization. Schechter et al. (2002) purified rabbit lacrimal acinar cells and demonstrated histotypic polarity in Matrigel rafts maintained for 28 days. 27 TJs act as selective barriers for the passage of fluid, salts, and proteins through the paracellular milieu. Moreover, TJs are involved in the regulation of proliferation, 28 differentiation, 29 and cell signaling. 30 Selvam et al. (2007) described the polarity and the ability of rabbit lacrimal acinar cells to generate short circuit current characteristic of chloride secretion when cultured on microporous polyester membranes. They localized Na+-K+-ATPase and demonstrated the direction of chloride secretion to be apical. 31 In previous studies,10,11 our group has evaluated the presence of Na+-K+-ATPase in human and monkey salivary cells when cultured on plastic. Experiments done by Schechter et al. (2002) (using Matrigel) 27 and by Selvam et al. (2007) (on polyester membrane) 31 would be useful to determine the direction of key ions secretion in huSG cells on Matrigel. McDonald et al. (2009) demonstrated the influence of certain secretagogues (histamine and serotonin) on the vectorial secretion of b-hexosaminidase in lacrimal acinar cells from rabbits. 32 The ability to maintain the polarized distribution of TJs, Na+-K+-ATPase, various ion co-transporters, antiporters, and channels are essential for fluid secretion. Further experiments are needed to study such proteins and their functionality in 3D salivary acinar units grown on Matrigel in the presence of certain salivary secretagogues.
We found that huSG cells proliferate faster on plastic than on Matrigel, as recorded by the MTT assay (Fig. 4). The proliferation and viability of huSG cells on Matrigel were reduced significantly (at day 7 and 9, p < 0.05), whereas apoptosis increased significantly (at day 7, p < 0.01) when compared to huSG cells grown on plastic. The decrease in the proliferation rate of huSG cells grown on Matrigel was consistent with findings of the HSG cell line33–35 and of primary huSG. 20
We investigated the phenotypic (by immunofluorescence staining) and functional characteristics (by Western blot) of huSG cells cultured on Matrigel for salivary-specific markers (e.g., amylase). Immunofluorescence staining resulted in 99% amylase-positive cells on Matrigel but with only a two- to five-folds increase in secreted amylase from Western blot results. The huSG cells induced into an acinar phenotype by Matrigel might be still immature acini (as 99% of them were still positive for vimentin 36 ) and had delayed or retained amylase secretion into the media. Although the 3D acinar units formed lumens (on Matrigel) and secreted a higher concentration of AMY into the media, we do not know whether AMY secretion occurred through these lumens (although we assumed this was likely as the secretory granules were located at the apical end of the cell based on TEM examination). Additional studies are needed to determine the extent to which stimulation with sympathomimetic agent, such as isoproterenol, would affect the rate and the direction (apical or basolateral) of amylase secretion from huSG cells.
The level of downregulation in gene expression (quantitative RT-PCR results for AMY, AQP-5, CLDN-1, KLK-1, EGF, and CK-18) on Matrigel varied between cultures derived from different patients. Similar findings were reported by Szlavik et al. (2008). 20 However, TJ proteins immunoreactivity was evident with comparable percentages in both culture types. The overall data presented here showed that Matrigel tended to direct the cells toward an acinar phenotype. However, long-term (>9 days) experiments would be useful to determine the effect of other variables such as the patient's age and their associated diseases on the capacity of the salivary cells to regenerate.
Although useful as an in vitro 3D, our model cannot be used for clinical application, as Matrigel is derived from a mouse sarcoma. Looking for a substitute for Matrigel, we are currently testing other gel formulations amenable for clinical testing. However, the ability to grow and maintain mixed cell culture containing all huSG components in 3D acinar units will have important implications in the study of regenerating salivary glands or as graft cells in a tissue-engineered artificial salivary gland device (in an animal model). In addition, for further in vitro studies of huSG physiology, it is important to simulate the normal gland in vivo by culturing mixed cells. Therefore, our 3D culture model provides a unique approach for studying the expression of specific salivary proteins in vitro. Innovative cell culture methods will be important to generate sufficient amounts of cells with the capacity to differentiate and replace damaged salivary tissues.
Footnotes
Acknowledgments
The authors are grateful to Dr. Christopher Moskaluk for the histopathological analysis of salivary glands and Cooperative Human Tissue Network for providing the salivary tissue samples. We are most appreciative of the technical help provided by Mrs. Jeannie Mui and Facility of Electron Microscopy Research (FEMR) at McGill University for TEM samples preparation and Mr. David Hori, the pathologist assistant at Royal Victoria Hospital. The authors would like to thank the following funding agencies: Natural Sciences and Engineering Research Council (NSERC) of Canada, Le Reseau de Recherche en Sante Buccodentaire et Osseuse (RSBO), and the Egyptian Ministry of Higher Education and Scientific Research.
Disclosure Statement
No competing financial interests exist.