
Abstract
Regenerative therapy using stem cells is a promising approach for the treatment of stroke. Recently, we reported that CD31−/CD146− side population (SP) cells from porcine dental pulp exhibit highly vasculogenic potential in hindlimb ischemia. In this study, we investigated the influence of CD31−/CD146− SP cells after transient middle cerebral artery occlusion (TMCAO). Adult male Sprague-Dawley rats were subjected to 2 h of TMCAO. Twenty-four hours after TMCAO, CD31−/CD146− SP cells were transplanted into the brain. Motor function and infarct volume were evaluated. Neurogenesis and vasculogenesis were determined with immunochemical markers, and the levels of neurotrophic factors were assayed with real-time reverse transcription–polymerase chain reaction. In the cell transplantation group, the number of doublecortin-positive cells increased twofold, and the number of NeuN-positive cells increased eightfold, as compared with the control phosphate-buffered saline group. The vascular endothelial growth factor level in the ischemic brain with transplanted cells was 28 times higher than that in the normal brain. In conclusion, CD31−/CD146− SP cells promoted migration and differentiation of the endogenous neuronal progenitor cells and induced vasculogenesis, and ameliorated ischemic brain injury after TMCAO.
Introduction
Materials and Methods
Isolation of CD31−/CD146− SP cells
CD31−/CD146− SP and CD31+/CD146− SP cells were isolated from porcine tooth germ, as described previously. 5 CD31−/CD146− SP cells were cultured in endothelial basal medium-2 (EBM-2, single quots cc–4176) with 10 ng/mL insulin-like growth factor 1 (IGF1), 10 ng/mL epidermal growth factor (EGF), and 10% fetal bovine serum (FBS). CD31+/CD146− SP cells were cultured in EBM-2 with 10 ng/mL bFGF, 10 ng/mL vascular endothelial growth factor (VEGF), 138 nM hydrocortisone, 0.09 mg/mL heparin, 50 μg/mL ascorbic acid, and 10% FBS. They were routinely subcultured up to 70% confluence under identical conditions.
Cerebral ischemia model
All animal experiments were approved by the Institutional Animal Care and Use Committee (National Center for the Geriatrics and Gerontology). Adult male Sprague-Dawley rats (Japan SLC, Inc.) weighing 300–400 g were used. Animals were initially anesthetized with 5% isoflurane (Abbott Laboratories) and maintained under anesthesia with 1.5% isoflurane in a mixture of 70% N2O and 30% O2. Rectal temperature was maintained at 37°C±0.5°C on a heating pad. Focal cerebral ischemia was induced by TMCAO with 2 h. 6 A 4-0 monofilament nylon suture (Shirakawa) with the tip rounded by flame heating and silicone (KE-200; Shin-Etsu Chemical) was advanced from the external carotid artery into the internal carotid artery until it blocked the origin of the MCA. Two hours after occlusion, reperfusion was performed by withdrawal of the suture. The regional cerebral blood flow of the MCA territory was measured using a laser-Doppler flowmeter (Omega FLO-N1; Omega Wave, Inc.) after occlusion. The response was considered positive and included only if the reduction in regional cerebral blood flow was >70%.
Transplantation
Twenty-four hours after TMCAO (day 0), the rats were again anesthetized with sodium pentobarbital (Schering-Plough) (0.25 mL/kg, intraperitoneally) and maintained under anesthesia with 1.5% isoflurane in a mixture of 70% N2O and 30% O2. Animals were randomly divided into three groups: (I) CD31−/CD146− SP cell transplantation group (n=24, day 3 sacrificed=6, day 9 sacrificed=7, day 21 sacrificed=11), (II) unfractionated pulp cell transplantation group (n=4, used for motor function), and (III) vehicle alone (phosphate-buffered saline [PBS]) group (n=20, day 3 sacrificed=6, day 9 sacrificed=5, day 21 sacrificed=9). The infarction site was targeted for transplantation at the striatum of the following coordinates: 1.0 mm rostral to the bregma, 6.0 mm lateral to the midline, 5.0 mm ventral to the dura (Fig. 1A, B). Subsequently, 1×106 CD31−/CD146− SP cells or unfractionated pulp cells at the fifth to seventh passage after labeling with 1,1-dioctadecyl-3,3,3,3 tetramethylindocarbocyanine perchlorate (DiI; Sigma), and removing all added factors into each medium were diluted with 2 μL of PBS, and were transplanted by Hamilton microsyringe (Hamilton). The control group consisted of an equal volume of PBS injected into the same site.

Immunohistochemistry
At day 3 or 21 after injection, the rat was transcardially perfused with 4% paraformaldehyde solution (Nakarai Tesque). The brain was removed and postfixed in paraformaldehyde. The following day, it was immersed in 30% sucrose solution. Twelve-micrometer-thick coronal sections were cut on a cryostat. For immunohistochemistry, the sections were preincubated in blocking solution (PBS containing 5% normal serum of the species in which the secondary antibody was raised) for 2 h at room temperature, and incubated with primary antibodies diluted for 1 h at room temperature. The primary antibodies were as follows: neuronal progenitor cells (NPC) marker, rabbit anti-doublecortin (1:50; Abcam, Inc.); neuron marker, rabbit anti-neurofilament H (1:200; Chemicon) and mouse anti-NeuN (anti-neuronal nuclei, 1:500; Chemicon); endothelial cell marker, mouse anti-RECA1 (rat endothelial cell antigen; Monosan); apoptosis marker, rabbit anti-cleaved caspase-3 (1:50; Cell Signaling Technology, Inc.); and VEGF marker, rabbit anti-VEGF (VEGF [P-20]: sc-1836; Santa Cruz Biotech). After washing, sections were incubated for 1 h at room temperature with secondary antibodies (on day 21, for neurofilament H/doublecortin, Donkey anti-rabbit IgG FITC [1:400; Jackson ImmunoResearch]; for NeuN/RECA1, Goat anti-mouse IgG FITC [1:200; MP Biomedicals]; and for VEGF, rabbit anti-goat IgG-HRP [1:400; Invitrogen Corporation]. On day 3, for cleaved caspase-3, goat anti-rabbit IgG-HRP [1:400; Invitrogen] and for VEGF, rabbit anti-goat IgG-HRP [1:400; Invitrogen]). The sections with HRP-conjugated secondary antibodies were incubated in anti-fluorescein-HRP (1:400; TSA™ Fluorescence Systems; PerkinElmer) for 7 min at room temperature. Adjacent sections were used as negative controls. In the control sections, all procedures were processed in the same manner except that the primary antibodies were omitted. To identify migration of NPC from SVZ, we observed the cryosections on days 9 and 21 with anti-doublecortin on fluorescence microscope (BZ-9000; Keyence) and BZ-HIC (Keyence).
Statistical analyses of the density of cells
The density of NPCs, neurons, endothelial cells, and apoptotic cells in the peri-infarct area (Fig. 1C) and the contralateral region in the CD31−/CD146− SP cell transplantation group and PBS groups were determined. In all groups (PBS group, CD31−/CD146− SP cell transplantation group, and contralateral group, n=3), each five sections at every 120-μm were stained with doublecortin, NeuN, RECA1, and cleaved caspase-3. The microscopic images were scanned and five typical frames (0.49 mm2) were measured for each section. Thus, 75 frames on an average were determined per group. The positively stained area relative to total area (7.41 mm2) was statistically analyzed using a Dynamic cell count, BZ-HIC (Keyence).
Real-time reverse transcription–polymerase chain reaction
Total RNA on cryosamples was extracted using Trizol (Invitrogen) from the area of the DiI-positive cells observed in the section. First-strand cDNA syntheses were performed from total RNA by reverse transcription with ReverTra Ace-α (Toyobo). Real-time reverse transcription–polymerase chain reaction (RT-PCR) amplifications were performed at 95°C for 10 s, at 62°C for 15 s, and at 72°C for 8 s using the porcine-specific primers VEGF. The specificity of the primers to porcine was confirmed by no amplification of the first-strand cDNA from rats with normal brains. The RT-PCR products were subcloned into a pGEM-T Easy vector (Promega) and confirmed by DNA sequencing based on published cDNA sequences. Gene expression of the transplanted cells in the infarct area was compared with that in the porcine normal brain tissue and that in transplanted cells in the normal brain after normalizing with β-actin.
In situ hybridization
Neurotrophic factors expressed in CD31−/CD146− SP cells were examined with in situ hybridization in cryosections on day 21. Porcine cDNA of VEGF (224 bp), glial cell line-derived neurotrophic factor (GDNF; 144 bp), brain-derived neurotrophic factor (BDNF;180 bp), and nerve growth factor (NGF; 210 bp) were linearized with NcoI, SpeI, NcoI, and SpeI, respectively, for anti-sense probes, and linearized with SpeI, NcoI, SpeI, and NcoI, respectively, for sense probes. The VEGF probe was constructed from plasmids after subcloning the PCR products using the same primers designed for real-time RT-PCR. The GDNF, BDNF, and NGF probes were also constructed in same way as the VEGF probe. Since a published porcine GDNF sequence was not available, human primers for GDNF (forward 5′-TATGGGATGTCGTGGCTGT-3′, reverse 5′-TCCACACCTTTTAGCGGAAT-3′) were used for cDNA subcloning of porcine GDNF (630 bp). The design of the oligonucleotide primers (Table 1) was based on both published porcine cDNA sequences and the newly cloned cDNA sequence of the porcine GDNF. The four probes were labeled with DIG (Invitrogen) and the DIG signals were detected with TSA system FITC-conjugated tyramide (Invitrogen).
BDNF, brain-derived neurotrophic factor; GDNF, glial cell line-derived neurotrophic factor; NGF, nerve growth factor; VEGF, vascular endothelial growth factor.
Migration, proliferation, and anti-apoptotic assays
At 50% confluence, the culture medium was switched to serum-free EBM-2. The conditioned medium (CM) from CD31−/CD146− SP cells, CD31+/CD146− SP cells, and unfractionated pulp cells were collected after 48 h.
For migration assay, modified Boyden chamber assays were performed with polyethylene terephthalate membrane (BD Bioscience) in a 24-well plate (BD Bioscience). SHSY5Y cells (Sanyo Chemical Industries, Ltd.) (1×105 cells/well) were seeded on the insert polyethylene terephthalate membrane, and 500 μL of DMEM-F12 (Sigma) with 20% of the three CMs was, respectively, poured into the tissue culture 24-well plate. SHSY5Y cells were derived from a neural crest tumor of early childhood, predominantly composed of undifferentiated neuroblast-like cells. 7 After 24 h, the SHSY5Y cells passing through the membrane were counted after detaching them with 0.05% trypsin–0.02% EDTA.
For cell proliferation assay, SHSY5Y cells (1×103/96-well plate) were cultured in DMEM-F12 containing 10% FBS for 24 h, and subsequently in serum-free DMEM-F12 containing 0.2% bovine serum albumin for further 24 h. Then, the medium was changed into each DMEM-F12 containing 0.02% FBS with 20% of three CMs. Ten micrometers of Tetra-color one (Seikagaku Kogyo, Co.) was added to the 96 well-plate, and cell numbers were measured by spectrophotometry at 450 nm at 2, 12, 24, 36, and 48 h of culture.
For the anti-apoptotic assay, SHSY5Y cells were cultured in DMEM-F12 in a 35-mm dish for 2 days and then incubated with 300 nM staurosporine 8 (Sigma) in DMEM-F12 with 20% of the three CMs. After 24 h, SHSY5Y were harvested, and the cell suspensions were treated with Annexin V-FITC (Roche Diagnostics) and PI for 15 min, and analyzed by flow cytometry JSAN.
BDNF (Peprotech), GDNF (Peprotech), VEGF-A (Peprotech), or NGF (Peprotech) at 50 ng/mL was used as a control for the three assays.
Evaluation of motor disability
Rats were blindly examined on days 0, 2, 6, and 9 with a standardized motor disability scale by slight modifications. 9 Rats were scored 1 point for each of the following parameters: flexion of the forelimb contralateral to the stroke when instantly hung by the tail, extension of the contralateral hind limb when pulled from the table, and rotation to the paretic side against resistance. In addition, 1 point was scored for circling motion to the paretic side when trying to walk, 1 point was scored for failure to walk out of a circle of 50 cm in diameter within 10 s, 2 points were scored for failure to leave the circle within 20 s, and 3 points were scored for inability to exit the circle within 60 s. In addition, 1 point each was scored for inability of the rat to extend the paretic forepaw when pushed against the table from above, laterally, and sideways. The motor disability scale was performed 3 times per animal time-point.
Assessment of infarct volume
The cryosections obtained from samples on days 3 and 21 were stained with hematoxylin and eosin. 10 ImageJ (National Institutes of Health) was used to determine each infarct area in 9 coronal sections in 12-μm thickness at 0.84-mm intervals. All of the infarction area was covered by these nine coronal sections. Regional infarct volumes were calculated by summing the infarct areas and multiplying these areas by the distance between sections (0.84 mm), followed by remediation for brain edema. 11
Statistical analyses
Data are reported as means±SD. p-Values were calculated using the unpaired Student's t-test.
Results
Pulp stem cell outcome
DiI-labeled transplanted CD31−/CD146− SP cells were characterized by round-to-oval nuclei with minimal variable cytoplasm. The transplanted cells survived and migrated from the original injection site to peri-infarct area in the cortex and striatum (Fig. 1D).
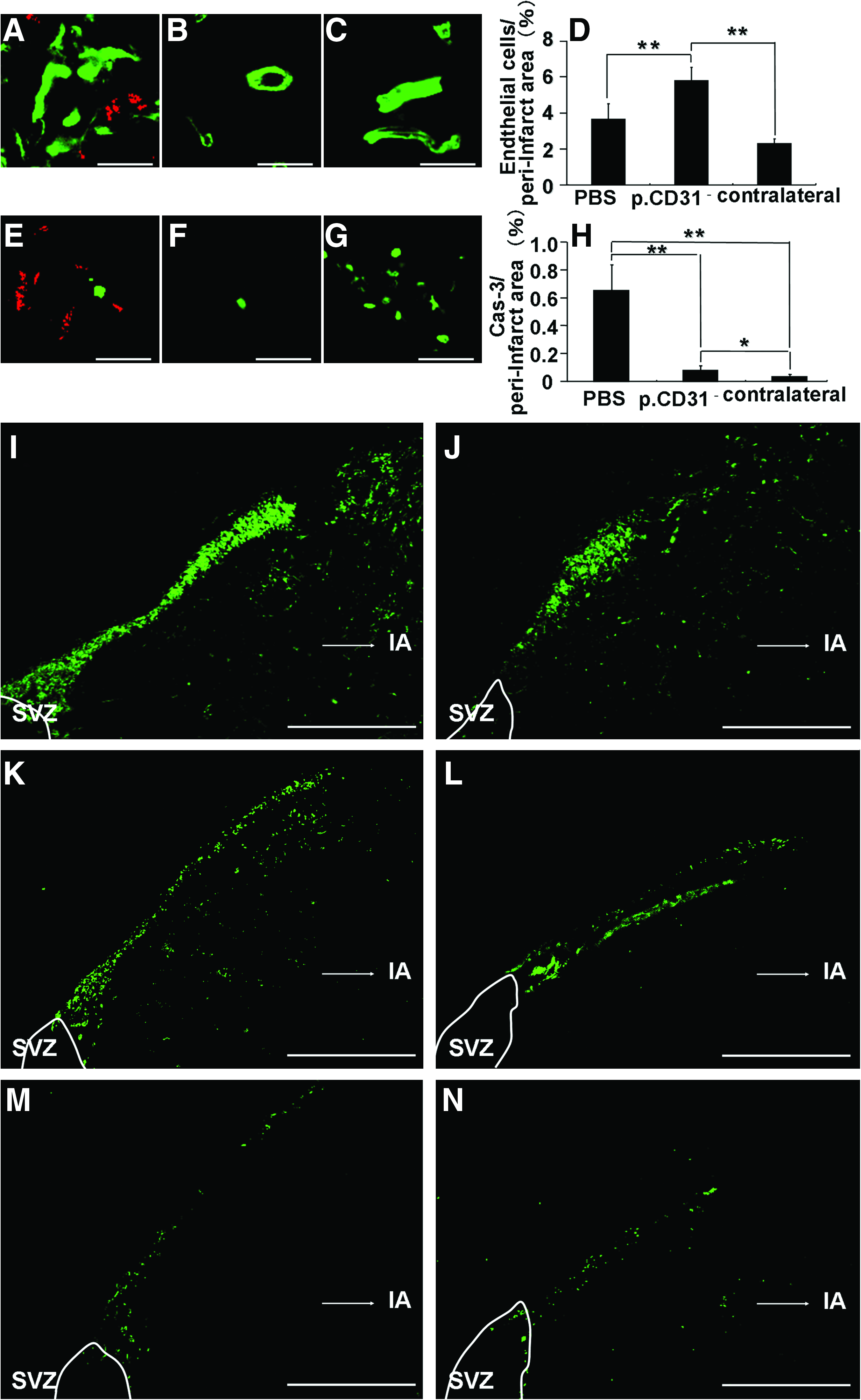
Transplanted cells localized in proximity of doublecortin (Fig. 2A) and neurofilament (Fig. 2D) or NeuN-positive cells (Fig. 2G) on day 21. Few doublecortin cells were observed in the contralateral side (Fig. 2B). There was a twofold increase in doublecortin-positive cells (Fig. 2J) and an eightfold increase in NeuN-positive cells (Fig. 2K) on day 21 in the CD31−/CD146− SP cell transplantation group compared with that in the PBS group. No evidence of differentiation of CD31−/CD146− SP cells into neurons or endothelial cells was detected. The migration of NPCs with doublecortin from SVZ to the peri-infarct area was observed on days 9 and 21. The migration on day 9 was more prominent (Fig. 3I, K, M). These results suggest that the transplanted cells support the migration and differentiation of the NPCs. The number of RECA1-positive cells on day 21 was increased in the CD31−/CD146− SP cell transplantation group compared with that in the PBS group (Fig. 3D), indicating that the transplanted cells also promote angiogenesis after ischemia. In the CD31−/CD146− SP cell transplantation group (Fig. 3H), there was a decrease in cleaved caspase-3-positive cells, suggesting that the transplanted cells have an anti-apoptotic function.
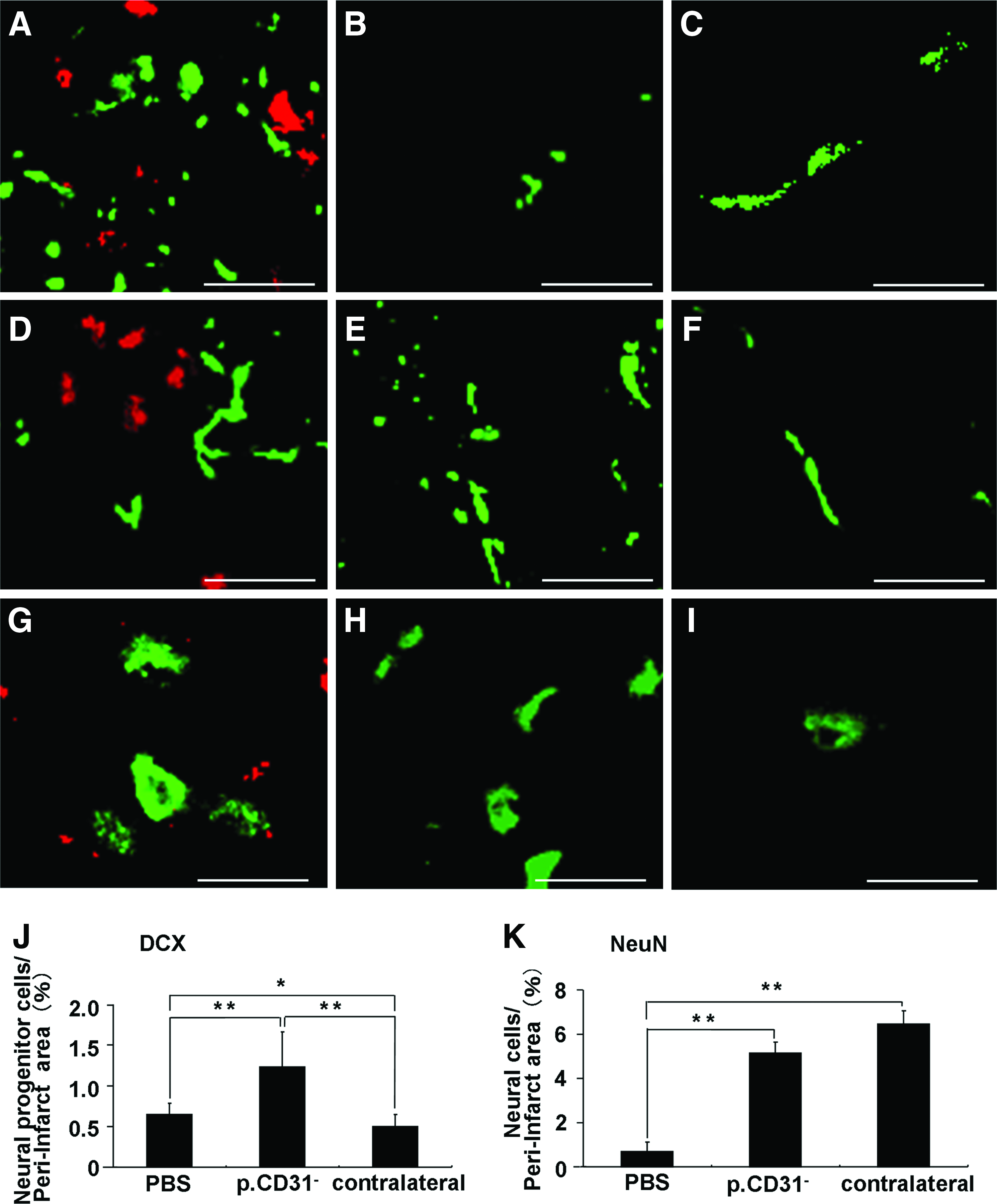
Doublecortin-positive cells
RECA1-positive cells on day 21
Expression of neurotrophic factors
The expression of several neurotrophic factors VEGF, GDNF, NGF, and BDNF was detected with in situ hybridization in the DiI-labeled CD31−/CD146− SP cells in the peri-infarct area on day 21 (Fig. 4A–D). Real-time RT-PCR analysis demonstrated that expression of VEGF mRNA by the transplanted CD31−/CD146− SP cells in the ischemic region on day 21 was 1,000 times and 28 times higher than that of normal porcine brain and that of the transplanted CD31−/CD146− SP cells into normal rat striatum, respectively (Fig. 4E, F). Immunohistochemistry of VEGF showed that the VEGF protein was highly expressed in the DiI-labeled CD31−/CD146− SP cells in the peri-infarct area on day 3 (Fig. 4G) compared with that on day 21 (Fig. 4H).

Analysis of expression of VEGF
Migration, proliferation, and anti-apoptotic assays
CM of CD31−/CD146− SP cells showed higher migratory effect on SHSY5Y cells than VEGF, NGF, and BDNF, and was similar to GDNF (Fig. 5A). Its proliferation effect was higher than VEGF and NGF, and similar to BDNF and GDNF (Fig. 5B). Its anti-apoptotic activity was higher than BDNF, GDNF, and VEGF (Fig. 5C).
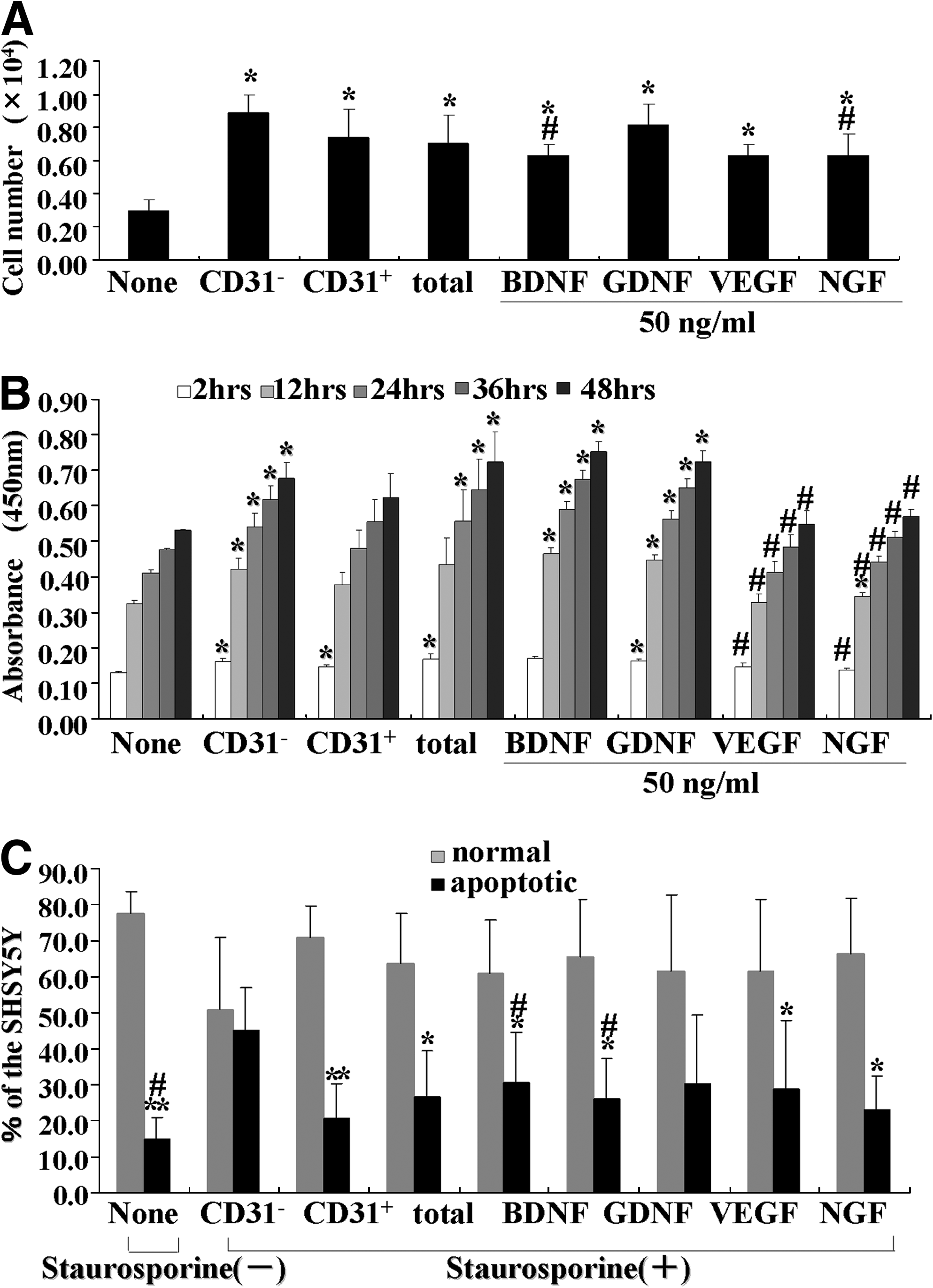
The migration
Evaluation of motor function
All groups (CD31−/CD146− SP cells, unfractionated pulp cells, and PBS) displayed high score for motor function at the early stage (day 0, scores are 8.08±0.79; 8.25±0.96; 8.42±0.79, and day 2, 5.08±0.90; 6.25±1.26; 7.67±0.78, respectively). Progressive improvement in motor disability in the CD31−/CD146− SP cell transplantation group after day 2 became significant on day 6 compared with PBS control group (2.67±1.23; 6.83±0.72), and more significant on day 9 compared with the unfractionated pulp cells and the PBS control group (1.33±0.78; 2.8±0.96; 6.50±0.67) (Fig. 6A). Persistent improvement in CD31−/CD146− SP cells transplantation group was noted on day 14 (1.00±0.71) and 21 (0.80±0.45), whereas persistent impairment of motor disability (score above 4) was observed in the PBS group on day 14 (5.60±0.55) and 21 (5.40±0.59) (Fig. 6A). Further, the video image demonstrated significant recovery in motor function of the CD31−/CD146− SP cell transplantation group compared with the unfractionated pulp cells and PBS control groups on day 6 (Supplementary Videos S1–S3; Supplementary Data are available online at www.liebertonline.com/tea).

Motor disability test by injection of the CD31−/CD146− SP cells, the unfractionated pulp cells and the PBS on days 0, 2, 6, and 9
Reduction of infarct volume
There was a significant decrease in the infarct volume on days 3 and 21 in the CD31−/CD146− SP cell transplantation group (day 3, 95.2±2.5 mm3, n=3: day 21, 84.7±6.5 mm3, n=4) compared to PBS group (day 3, 109.7±4.1 mm3, n=3: day 21, 123.9±7.4 mm3, n=4). The difference of infarct volume between the CD31−/CD146− SP cell transplantation group and the PBS group increased over time (reduced by 13.3% on day 3 and reduced by 32.9% on day 21) (Fig. 6D). These results suggest that transplanted CD31−/CD146− SP cells promoted the regeneration.
Discussion
In the current study, we demonstrated that transplanted CD31−/CD146− SP cells migrated to the peri-infarct area. In addition, these cells released neurotrophic factors, and promoted migration and differentiation of the endogenous NPCs in SVZ. They also induced vasculogenesis in the peri-infarct area. These results indicate that CD31−/CD146− SP cells ameliorated the ischemic tissue injury and accelerated the functional recovery after TMCAO. We have hypothesized that three mechanisms may contribute to the actions of VEGF. First, VEGF produced by transplanted cells may promote neurogenesis. NPCs in SVZ are known to migrate to the peri-infarct area and differentiate into neurons. 1 In this study, VEGF induced a chemotactic response in SHSY5Y cells. The transplanted CD31−/CD146− SP cells migrated to the peri-infarct area and expressed VEGF. These results suggest that VEGF released by CD31−/CD146− SP cells in the peri-infarct may promote migration of the endogenous NPCs in SVZ. Second, VEGF produced by transplanted cells may promote vasculogenesis. VEGF binds to its receptors on locally present vascular endothelial cells and directly initiates the angiogenic response. 12 In this study, the number of RECA1-positive endothelial cells significantly increased in the cell transplantation group. Third, VEGF may provide a neuroprotective effect. The neuroprotective effects of VEGF in experimental cerebral ischemia have been reported. 13 In cell the transplantation group, the number of cleaved caspase-3-immunopositive cells in the peri-infarct area was decreased compared with that in the PBS group, thus demonstrating the anti-apoptotic effects of VEGF on SHSY5Y cells. These results suggest that VEGF produced by CD31−/CD146− SP cells may inhibit apoptosis of neurons. Thus, VEGF demonstrates pleiotropic effects on neurogenesis, vasculogenesis, and neuroprotection.
As VEGF is a potent vascular permeability factor, it may accelerate brain edema after stroke. Administration of VEGF in early ischemia (1 h after ischemia) leads to significant increase in blood–brain barrier leakage as well as enlarged ischemic areas. 14 However, VEGF administration at 24 h after TMCAO reduces infarct size, improves neurologic recovery, enhances neurogenesis in the SVZ and angiogenesis in the ischemic border zone. 14 In this study, CD31−/CD146− SP cells were transplanted 24 h after TMCAO and we monitored the reduction of infarct size and improvement of motor disability. The time of administration of cells is critical. Thus, if CD31−/CD146− SP cells were transplanted during an optimal window of time, they exhibit beneficial effects without the deleterious effects of edema.
In addition, CD31−/CD146− SP cells expressed other neurotrophic factors such as GDNF, 15 NGF, 16 and BDNF 16 in the peri-infarct area. These neurotrophic factors had migratory, proliferative, and/or anti-apoptotic effects on SHSY5Y cells in vitro and may also contribute to the recovery from ischemic brain injury.
Finally, we explored the plausible underlying mechanisms of how injection of CD31−/CD146− SP cells into the brains of immunocompetent rats staved off graft rejection. Blood–brain barrier is known to play a critical role in maintaining the immune-privileged status of the central nervous system. 17 It is well known that mesenchymal stem cells from bone marrow are not rejected by hosts and immunosuppression is not required in rodents. 1 Dental pulp stem cells have many similarities to mesenchymal stem cells; transplanted CD31−/CD146− SP cells possess immunosuppressive properties. 18
Conclusion
In summary, the transplantation of porcine CD31−/CD146− SP cells promotes neurogenesis and vasculogenesis in an induced peri-infarct area, and enhances recovery after TMCAO in rats. Further research is needed to understand the underlying mechanisms. For potential clinical application and translational studies, the safety of CD31−/CD146− SP cells must be assessed, including tumor formation. In conclusion, regeneration therapy using CD31−/CD146− SP cells is a potential candidate in the treatment of stroke.
Footnotes
Acknowledgments
The authors thank Drs. Masataka Ito, Kayo Adachi, and Kiyomi Imabayasghi for their assistance. This work was supported by funds from Collaborative Development of Innovative Seeds, Potentiality verification stage from Japan Science and Technology Agency, a Grant-in-Aid for Scientific Research from the Ministry of Education, Science, Sports and Culture, Japan, No. 19659499 (M.N.), No. 20390504 (M.N.), and No. 18592173 (H.H.), and the Research Grant for Longevity Sciences (19C-2, 21A-7) from the Ministry of Health, Labour, and Welfare (M.N.).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.